

Summary
菲PJ-34在肿瘤细胞进行有丝分裂的细胞毒活性,实时记录现场共聚焦成像。 PJ-34根除人类乳腺癌MDA-MB-231细胞有丝分裂窝藏额外中心体。额外中心体不像正常的双向焦点有丝分裂,不聚集在两个纺锤体的两极存在的PJ-34。
Abstract
菲衍生物作为有力PARP1抑制剂阻止编外中心体多中心体的人类癌症细胞有丝分裂的双焦聚类。菲啶PJ-34是最有力的分子。额外中心体的分簇多中心体细胞有丝分裂失败的原因和细胞死亡。最坚实的人力癌症有额外中心体的发生率较高。 PJ-34的活性共聚焦成像活的人乳腺癌MDA-MB-231细胞转染载体编码荧光γ-微管蛋白,这是非常丰富的中心体和荧光蛋白H2B目前在实时记录染色体。转染与PJ-34处理后的MDA-MB-231细胞中检测到异常的染色体安排和γ-微管蛋白聚集疫源代表declustered中心体。非集群额外两个纺锤体两极的中心体细胞死亡之前。这些结果与FOr的第一次检测独家额外中心体的聚类在有丝分裂,有丝分裂失败导致细胞死亡在人类癌细胞中的PJ-34的细胞毒活性。根据以前的研究结果固定细胞共聚焦成像,PJ-34独家消灭癌细胞的多中心体,而不损害正常细胞发生有丝分裂,两个中心和双焦主轴的观察。这PJ-34的细胞毒活性不共享等烈性PARP1抑制剂,观察,表明其独立性PARP1抑制PARP1缺失的MEF窝藏extracentrosomes。现场共聚焦成像提供了一个有用的工具,用于识别新的分子,消除细胞在有丝分裂过程中。
Introduction
菲衍生PARP1抑制剂,包括PJ-34,旨在保护耗能PARP1介导的DNA修复应力条件下(中风或心肌梗死)1静止期细胞诱导细胞凋亡。然而,最近我们发现,PJ-34,比诱导PARP1抑制浓度高出一倍,完全可以导致细胞死亡在人类癌细胞中2,3。更快速的细胞增殖,细胞更有效地消灭了。 PJ-34的细胞毒活性归因于额外中心体聚集在有丝分裂2。许多人类癌细胞海港multicentrosomes 4,5。孵化人类乳腺癌细胞MDA-MB-231,怀有编外中心体,为20μmPJ-34有效地根除这些细胞在72-96小时内不损害静止细胞或一些良性增生的细胞,窝藏两个中心在有丝分裂
双极双极4,5在有丝分裂纺锤体的形成是至关重要的中心体组装。因此,有两个以上的中心体的细胞已经开发了一种几乎不理解的分子机制,多余的中心体,在两极4-9聚类。双极其中心体组装的失败可能会导致多极化扭曲纱锭和异常染色体分离逮捕细胞周期G2 / M期阻滞,并导致细胞死亡归因于有丝分裂失败4,5。额外中心体聚类的分子机制的深入研究<SUP> 10。了解这个死亡机制将启用独占消灭癌细胞的同时保留健康组织5,10。
因此,激活有丝分裂灾难细胞死亡的化合物有选择性的癌症治疗提供了一种新的模式,这可能是有效率的在广泛的人类固体cancers.Our结果的建议聚焦成像,可用于识别分子影响额外中心体聚类2,3有丝分裂,使这些化合物癌症靶向药物候选人。
我们已经证明通过扫描固定的和活的人体癌症细胞(额外中心体在有丝分裂的发生率较高),相对于正常细胞的细胞毒活性对菲啶PJ 34。一步一步描述用于识别PJ-34在人类癌症细胞的细胞毒活性的成像程序包括下面。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1。细胞培养制备
MDA-MB-231细胞购自美国典型培养物保藏中心 (ATCC),并保存于液氮中。
- 种子10 6个MDA-MB-231细胞在92毫米直径的陪替氏培养皿在10毫升完全培养基中Dulbeco改进的Eagle培养基(DMEM),10%马血清,1%L-谷氨酰胺和1%两性霉素B。Penstrep允许细胞的激增至约80-100%汇合。
- 从盘取出培养基并丢弃。
- 简单细胞层洗净,用0.25%的胰蛋白酶-EDTA溶液(W / V)的血清清除所有痕迹。
- 添加2.0毫升胰蛋白酶-EDTA溶液菜倒置显微镜观察细胞,直到细胞层分散(通常在5至15分钟内)。
- 加入18毫升完全培养基,用吸管轻轻吸出细胞。转移到管。
- 在1,200 rpm离心细胞悬浮液。
- 重悬细胞沉淀在24毫升的铜介质lture。
- 加入2 ml的细胞悬液,以35毫米的玻璃底菜(约25%汇合),在培养箱(5%CO 2,37℃)。
- 解决方案:
- 的完整的细胞的增殖培养基:DMEM培养基含10%FBS,1%抗生素(100单位/毫升青霉素G,100微克/毫升链霉素,笔链球菌AMPHO的溶液)和2mM L-谷氨酰胺。
- 胰蛋白酶-EDTA溶液中,含有0.25%的胰蛋白酶-EDTA。
- 菜:
92毫米直径的培养皿。
直径35毫米,聚-D-赖氨酸镀膜玻璃底培养皿。
2。现场共聚焦成像单元的制备
- 种子2×10 5 MDA-MB-231细胞的玻璃底培养皿中第1节中提到的2毫升完全培养基。当细胞培养达到60-70%汇合(约3-4×10 5细胞每盘),继续转。
- 与ENCOD两个质粒转染的细胞ing的融合蛋白,γ-微管蛋白(GFP荧光检测中心体)和组蛋白红色(H2B红色,荧光检测染色体)采用脂质体转染试剂喷射PI,制造协议。简言之,混合2微克从每个质粒在管用100微升氯化钠(150毫米)。用100μl150mM的氯化钠()的第二管中的转染试剂(100微升)混合,并在室温(RT)下孵育5分钟。然后将二者结合起来的解决方案,混合(使用温和涡)和自旋向下。在RT下孵育30分钟。
- 在温育的转染混合物,洗涤细胞一次,用PBS和替换的细胞培养基中,用2毫升温暖的DMEM培养液(37℃)没有补充。
- 轻轻加在DMEM培养液中的细胞的转染混合物,然后返回的细胞培养箱中(37℃,5%CO 2)8小时。
- 8小时的潜伏期后,更换2毫升完全培养基DMEM细胞在孵化器和孵化24小时。
- 转染后24小时,更换的培养基中的细胞,用2毫升完全培养基中含有20μMPJ-34。
- 孵育细胞18小时(37°C,5%CO 2)。
- 主题的细胞生活共焦成像的细胞在5%CO 2和37℃下保持至少16小时,在成像室
- 同时,检查转染效率36小时转染后用荧光显微镜如下:
- 种子2×10 5的MDA-MB-231细胞在6孔板含有1盖玻片,在2毫升完全培养基中每孔的。
- 转染的细胞中提到的第2.2-2.5。
- 转染后36小时固定安装在盖玻片上培养转染的细胞在冷甲醇:丙酮(1:1)溶液,7分钟,-20℃下
- 吸出固定的解决方案,让安装的细胞,干化学罩盖玻片。
- 申请延长Gold抗淬灭试剂用DAPI,让日É盖玻片干燥6小时在黑暗中。
- 在荧光显微镜下检查的幻灯片,并计算转染的细胞(红色和绿色信号)从总人口的细胞(DNA用DAPI染色)的百分比。 100-200细胞计数时所需的转染百分率大约是20%至40%。
3。现场共聚焦成像扫描仪设置的技术参数
- ScanMode XYZT;针孔通风] 1.00;变焦3.5;分辨率8位,561纳米激光DPSS氩气,可见激光488纳米激光他/ NE可见的633纳米目的HCX PL APO CS 63X 1.40油抗紫外线;数值孔径1.4;扫描速度700赫兹;折射率1.52。
- 3-D图像演示制备Imaris里成像软件7.0。
4。固定细胞有丝分裂的共聚焦成像
- 种子为2×10 5 MDA-MB-231乳腺癌细胞(ATCC),正常小鼠胚胎成纤维细胞(MEF),或PARP1缺陷MEF(PARP - / - ,制备议员弗朗索瓦丝Dantzer)上在2毫升完全培养基的6孔板中的盖玻片。盖玻片,用96%乙醇洗涤后,用无菌DD水洗涤,干燥2小时,并放置在6孔盘的每个孔中。
- PJ-34(10-30微米)添加到培养基中,室温孵育规定时期内(通常可达96小时)。
- 一旦清洗盖玻片,用PBS(磷酸盐缓冲液),并固定细胞,用孵化,在冰冷的甲醇:丙酮(1:1)溶液,-20℃下,7分钟
- 吸固定溶液,并让盖玻片化学罩中干燥(在此阶段,盖玻片可在-20℃保存数周)。
- 用PBST(PBS含0.1%Tween-20的补充),一次清洗盖玻片通透细胞膜,并阻止细胞在RT下的1小时,用10%在PBST NDS(正常驴血清)(封闭溶液)。
- 孵育通透固定细胞原发性antibodiES的RT 2小时(主轴和中心体染色)。如下:抗α微管蛋白(1:250稀释)和抗γ-微管蛋白(1:200稀释)在封闭液稀释抗体。一抗的应用方式如下:适用于将100μl(在下拉式菜单中)为每个盖玻片的6孔板盖(盖是上下颠倒)的封闭溶液中的抗体的混合物。轻轻把盖玻片上的抗体下降,面临着下降的种子细胞。孵育面临的抗体在室温下搅拌2小时的盖玻片。
- 将盖玻片孔中,并用PBST洗涤细胞3次。然后用4.6所述的相同步骤,用荧光二级抗体标记细胞在盖玻片上。二次抗体1小时,RT孵育的细胞在盖玻片上,在黑暗中。该抗体稀释如下:封闭液的Alexa Fluor 488(1:1,000稀释,绿色)的Alexa Fluor 568(1:1,000稀释;红色)。
- 安装盖玻片使用延长黄金的抗淬灭试剂用DAPI(染色体染色),在室温孵育过夜,在黑暗干燥。
- 通过共聚焦显微镜检查盖玻片。
5。 ATP的测量细胞活力
ATP产量测定发光ATP检测试剂盒。
- 种子细胞的96孔培养板中,在各孔中的800微升培养基中大约20,000细胞。应该用于测定介质的背景发光的三个空白孔。
- 准备约10〜100μm的ATP标准品稀释系列,并保持在冰上。
- 向每孔中加入50μl的洗涤剂和为5分钟,每分钟700转的轨道摇床中摇板。
- 复溶试剂盒中的底物缓冲液,用5ml的的冻干基板的每个管形瓶中。
- 加入50微升底物溶液的重组到各孔中,并摇动板轨道摇床,700转5分钟。
- 保持板在黑暗的10分钟。
- 测量每孔luminiscence用ELISA酶标仪。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34是一种稳定的水溶性菲啶1( 图1)。我们先前的研究发现细胞死亡和集群额外中心体在几种类型的固定的多中心体肿瘤细胞,分别用PJ-34。相比之下,正常增殖细胞没有受损2,3。针对centrine1在固 定的额外中心体细胞2和γ-微管蛋白的抗体双标记进行鉴定中心体。
在这里,记录在这些现场额外中心体细胞的杀伤活性PJ-34实时通过现场共聚焦显微镜。现场人乳腺癌MDA-MB-231细胞,其中有一个发生率较高(> 50%)的额外中心体4,5,专注于与γ-微管蛋白GFP转染细胞共聚焦成像扫描至少16小时(荧光标记的γ-微管蛋白灶2)和组蛋白H2B红(荧光拉贝染色体玲)。六至十个活转染的细胞平行扫描,在每个实验中。 γ-微管蛋白在转染细胞centrin1灶双免疫标记在技术上是不可能的。
灶分散的γ-微管蛋白和异常染色体的安排,很少发现在随机选取的未经处理的MDA-MB-231细胞的有丝分裂。 γ-微管蛋白灶,相当于额外中心体双焦聚类,聚类双光记录在大多数现场未经处理的MDA-MB-231细胞( 图2),相比之下,非集群的中心体和染色体异常的安排现场检测转染的MDA-MB-231细胞培养与PJ-34(20μM),在这些细胞中有丝分裂的细胞死亡( 图3)结束。在这些细胞中加入PJ-34,18 -扫 描前24小时和16小时,在扫描过程中( 图3)。实时的文档的单元格的运动员在有丝分裂过程中强烈支持先前定义的人恶性细胞的数目与多极的主轴,在有丝分裂和PJ-34(20μM)2细胞中的细胞死亡的百分比的正相关关系。
PJ-34作为一个强有力的PARP1抑制剂1。因此,我们研究PARP1抑制导致细胞死亡与有丝分裂失败的可能性。实时成像不同,固定的MDA-MB-231细胞的成像启动,检查一个人口众多的细胞培养物中的细胞,从而使PARP1抑制剂在多种人类肿瘤细胞株的影响的统计分析。 PJ-34的活性进行比较等烈性,非菲PARP1抑制剂在正常或PARP1缺失的细胞( 即正常和PARP1( - / - )小鼠胚胎成纤维细胞(MEF))( 图4)的活性。 PARP1缺失MEF海港多中心体在有丝分裂,但他们没有吨肿瘤细胞11。这些细胞制备的弗朗索瓦丝Dantzer博士,法国斯特拉斯堡。
固定正常和PARP1( - / - )MEF免疫标记的α-和γ-微管蛋白的标记,其心轴和中心体,分别为2之前,如报告。某些检查的细胞培养用PJ-34或其它有效的非菲PARP1抑制剂,包括ABT-888和AG01469,抑制PARP1的酶活性,BSI-201的化合物,显然衰减PARP1结合12-14缺口的DNA。没有被测PARP1抑制剂浓度抑制PARP1活性( 图4)正常的MEF受损。与此相反,PJ-34剂量依赖性地引起未聚类的γ-微管蛋白灶,主轴和细胞死亡的失真PARP1( - / - ):MEF( 图4A和B)。这是不遵守正常MEF治疗与PJ-34(FIGURë键4B)或在PARP1( - / - )MEF用PARP1抑制剂非phenenthrene的ABT-888或AG014699( 图4C)。应该指出的是PJ-34的浓度超过20μM并妨碍正常的MEF,虽然正常的MEF PJ-34的活性比PARP1更耐( - / - )MEF。
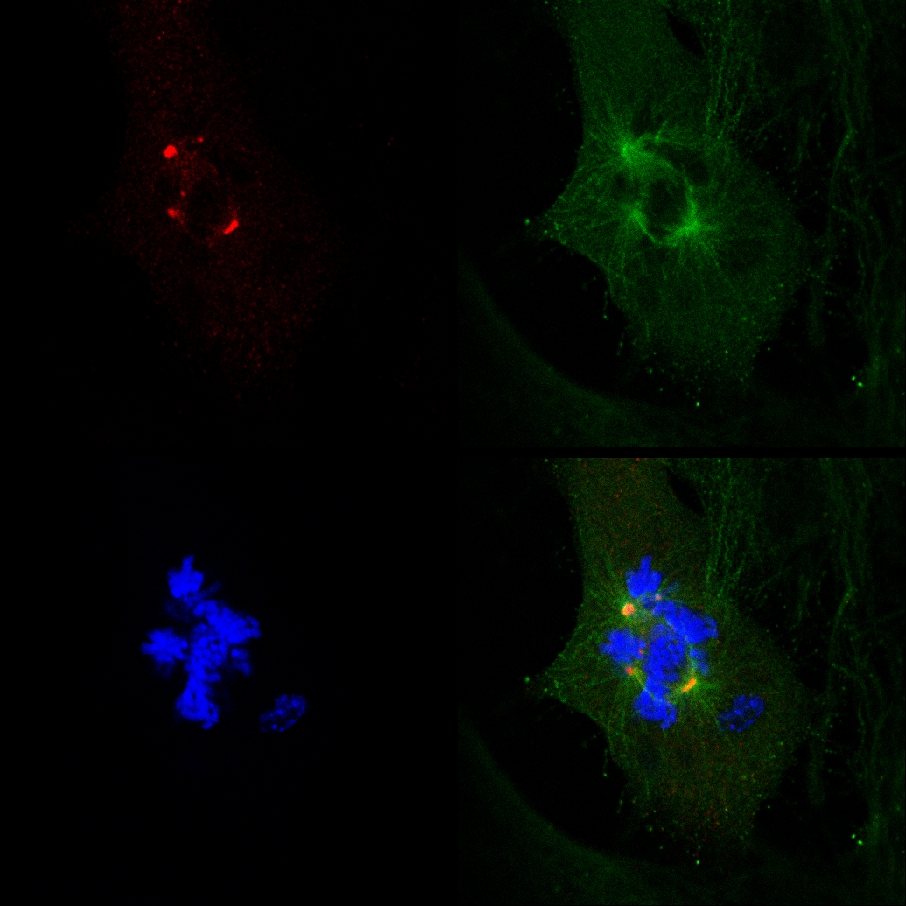
的事实,PJ-34根除PARP1( - / - )MEF尽管PARP1不足的,多焦点的主轴的形成和消除细胞PARP1( - / - )的 MEF培养PJ-34的浓度之间的相关性高于PARP1抑制需要,不符合额外中心体聚类PARP1( - / - )MEF和PARP1抑制( 图4A)之间的因果联系。 PARP1 PJ-34的细胞毒性活性( - / - )MEF可以更好地解释其活性作为一个额外的中心体聚类剂在多中心体的细胞2( 补充资料 “/> 图3。 在随机选择的MDA-MB-231细胞的有丝分裂与非集群γ-微管蛋白标记的额外中心体细胞死亡的现场共聚焦成像文档。在扫描之前和过程中16小时的活的共焦成像,MDA-MB-231细胞与PJ-34 24小时。 PJ-34采用γ-微管蛋白GFP(绿色,γ-微管蛋白灶和中心体)和H2B-RED(红色,染色体)的表达载体转染后24小时。 
图1。菲啶PJ-34:N-(6 -氧代-5,6 -二氢-phenanthridin -2 -基)-N,N-二甲基乙酰胺。 
图2。 A.双焦聚类额外中心体在现场随机选择的MDA-MB-231细胞的有丝分裂。上面板 :在一个随机选择的活MDA-MB-231细胞转染γ-微管蛋白-GFP。 面板 :在一个随机选择的MDA-MB-231细胞转染的组蛋白H2B-RED的有丝分裂过程中染色体重新安排的标记的中心体。 B.双焦点有丝分裂集群额外中心体确定在一个随机选择的邪教主组分的MDA-MB-231细胞。细胞转染γ-微管蛋白(GFP标签γ-微管蛋白灶;绿色)和组蛋白H2B RED(标记染色体,红色)。转染后48小时,细胞暴露于16小时现场共焦成像。在每个实验中,6个细胞,并行扫描。四个不同的实验进行。另见补充资料。 点击这里查看大图 。 
图3。额外中心体聚类细胞死亡之前,现场MDA-MB-231细胞的治疗与PJ-34。随机选取现场MDA-MB-231细胞的有丝分裂有散在中心体( 第一帧左 )截至细胞死亡( 第二和第三帧 )。此CELL随机选取24小时培养的细胞培养与PJ-34(20微米)申请24小时后,转染载体表达γ-微管蛋白标签灶,包括中心体γ-微管蛋白GFP(绿色)和组蛋白H2B红色(标记染色体;红色)。细胞16小时现场共聚焦成像扫描。在每个实验中,6个细胞,并行扫描。三种不同的实验进行。另见补充资料。 点击这里查看大图 。 
图4。 PARP1 PJ-34的细胞毒活性( - / - )小鼠胚胎成纤维细胞。 A.(左 )在n多焦点主轴计算的百分比ormal(黑线)和PARP1 - / - (灰色线)MEF PJ-34在指定的浓度,培养48小时。多极化纱锭的百分比计算出20总纱锭在3个不同的实验检测。 ( 右 )降低细胞的存活检测到的温育72小时,在细胞培养用PJ-34(20μM)相控制的未经处理的细胞的生存(正常(黑线)和PARP1 - / - (灰线),MEF)。细胞存活率测定细胞的ATP生产(协议5)。 4个测量值的平均值,每个细胞系中的3个不同的实验。B.主轴在随机选择的固定的正常PARP1 - / -的 MEF的有丝分裂,未经处理(对照组)或PJ-34温育48小时,在所指示的的浓度。 PJ-34造成多极化纱锭。细胞固定,透化和免疫标记的α-和γ-微管蛋白(绿色标记的主轴和红色标记的中心体,分别)。染色体用DAPI标记的试剂(蓝色)。代表3种不同的实验结果。C.强效非菲PARP1抑制剂并不会影响PARP1的聚类中心体( - / - )MEF。心轴的随机选择的正常和PARP1 - / - MEF,未经处理的MEF(对照组)或MEF非菲PARP抑制剂,AG01469(20μM)或ABT888(20μM)的48小时处理。染色体用DAPI标记的试剂(蓝色)。得到类似的结果,在3个不同的实验。 点击这里查看大图 。
点击此处查看大图 。 
点击此处查看大图 。
图2B。随机选择的MDA-MB-231细胞,后期与双焦集群γ-微管蛋白标记的扫描额外中心体γ-微管蛋白-GFP的表达载体转染后,由现场共焦成像,48小时和16小时的补充图。 H2B-RED(标签γ-微管蛋白发灶和在固定的细胞的中心体(绿色)和组蛋白H2B标记染色体(红色),)。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
实时共焦成像提供了一种实时PJ-34的细胞毒作用在活的多中心体的细胞在有丝分裂过程中( 图3和补充信息)的文档。这是第一次现场文档PJ-34在人类癌细胞中的细胞毒性归因额外中心体聚类和细胞死亡,表明诱导细胞死亡的有丝分裂灾难PJ-34 5-9。与此相反,双焦聚类数目的超中心体中观察到活的未处理的MDA-MB-231细胞进行正常的有丝分裂与双向focali的群集的多余的中心体( 图2,补充资料)。
根据这些结果,活转染细胞共聚焦成像牵连额外中心体聚类在转染细胞的蛋白质在有丝分裂过程中5,9,15,16检测易位可能是有用的。 PJ-34的影响在多centros的蛋白质的鉴定理解的死亡机制,激活额外中心体聚类,OMAL细胞可能会提供一些线索。
在活细胞的有丝分裂过程中提供实时信息,现场共聚焦成像的优势也势必一些限制。扫描每个实验的细胞的数量是有限的。因此,检测细胞在有丝分裂中的机会是低,并且所需要的一个实时文件扫描细胞的有丝分裂中的一些重复实验。此外,成功是高度依赖于高效率的被标记的蛋白质的表达载体转染的细胞。因此,尽管可靠,现场共聚焦成像是费时,需要经验丰富的工人。
在每个实验中,可靠的统计分析,这是需要大量的细胞,固定细胞的比较,免疫细胞化学和激光共聚焦成像启用检查。我们用这种方法来比较PJ-34影响正常的良性细胞的有丝分裂,其影响的肿瘤细胞进行有丝分裂编外中心体2,3。同样,此方法用于比较PJ-34非菲PARP1 PARP抑制剂的影响( - / - )MEF窝藏额外的中心体(细胞的发生率较高的多中心体11)( 图4)。
总之,我们的结果表明相结合的最有价值的实时信息,在有丝分裂中的活细胞( 图2和图3)与固定的细胞的细胞化学和激光共聚焦分析提供的共焦成像的优点。这些方法的组合可能是有用的用于识别的细胞毒活性的小分子,像PJ-34,针对特定的机制为细胞的生存至关重要。额外中心体的两极聚类许多人类癌症细胞的增殖和生存的唯一依赖人呈现PJ-34用于癌症治疗的一个可能的候选人。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者宣称,他们有没有竞争经济利益。
Acknowledgments
这项研究的资金来源:特拉维夫大学的技术转移公司,拉莫特和示巴医学中心(M. CA和SI),ICRF - 以色列癌症研究基金会(M. CA)和以色列科学基金会的联合基金( SI)。
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}