

Summary
수정되지 hyperphosphorylated 타우 단백질 과인산 화 - 의존성 응집 빠른 반응 속도를 나타 내기 위해 두 응집 시험 관내 어 세이에 사용 하였다. 상기 분석 방법은 알츠하이머 질환의 진행의 기초를 형성하는 피 브릴 hyperphosphorylated 타우 성향을 변조 할 화합물에 대한 미래의 화면에 대해 방법을 포장.
Introduction
(AD) 알츠하이머 병은 퇴행성 신경 알려진 신경 퇴행성 장애의 큰 컬렉션 중 하나이다. 전형적인 병리 기본 타우 병증은 신경 세포, 성상 세포와 미세 아교 세포 1-4의 신경 섬유 엉킴, NFTS입니다. NFT 밀도는인지 장애 3,5 및 신경 세포의 손실 6와 상관 관계. NFT는 직선 또는 쌍 나선형 필라멘트 (PHF) 7,8를 형성 (이하, "P-타우"이라한다) 주로 hyperphosphorylated 타우 단백질이 포함되어 있습니다. 타우 신경 신호 및 인신 매매 9,10 필수적이다 축삭 수송을 용이하게 생각 미세 소관 관련 단백질이다. 각 타우 분자는 2 ~ 3 정상적인 뇌의 인산염,하지만 타우 병증 환자 (11)의 여러 주름으로 포스 콘텐츠 증가가 포함되어 있습니다. 다중 키나아제는 GSK3β (글리코겐 신타 제 키나제 3β) 및 CDK5 (사이클린 드를 포함 타우 과인산 화에 기여할 가능성이있다늘어진 키나제 5) 12, 13,하지만 병적 인 인산화에 대한 직접 트리거가 어려운 14 남아있다. 또는 미세 소관 결합 모티프 근처 이상 인산화는 미세 소관 (15)에서 타우 해리 및 P 타우 결국 NFT 흠으로 중합 할 수 직선 또는 쌍 나선형 필라멘트에 oligomerizes somatodendritic 구획에 타우 잘못 지역화됩니다. 타우 과인산, NFT 형성 및 신경 퇴행 긴밀한 넥타이 P-타우 엉킴은 세포 사멸 및 기타 세포 독성 반응을 유도하고, 따라서 타우 병증의 신경 퇴행 16, 17에 대한 근본 원인임을 널리 가설되었다. 이 전제 약물 화면과 초기 임상 시험은 (18)을 출시했다. 그러나,이 가설은 (19, 20) 도전에 직면 해있다. 예를 들어,기도 Santacruz 등.이 트랜스 제닉 마우스의인지 기능이 돌연변이의 발현을 억제함으로써 개선 될 수 있음을 보여 주었다NFTS 기존 타우 분자 (21)로부터 계속하여 형성하더라도 인간 타우. 초파리 모델에서 근본적인 NFT는 신경 세포를 보호하기 위해 22, 23, 독성 세포질 타우를 격리시키는 것으로 나타났다. 분명히, NFT의 병인 역할은, 어떤 경우, 크게 타우 병증 치료제 개발의 방향에 영향을 미칠 것이다.
몇몇 β 시트 바람직한 형광 염료, 전자 현미경 및 광 산란 분광법 24-27의 결합으로 나타낸 바와 같이 높은 농도, 재조합 또는 정상 뇌 타우 단백질 자발적하지만 서서히 체외 PHF 형 구조로 중합한다. 추가 헤파린 또는 아라키돈 산, 인간 뇌에서의 풍부한 지방산 대폭 타우 isoform- 및 유도제 농도 의존적으로 28-32 PHF의 형성을 가속화. 흥미롭게도, hyperphosphorylated 타우는 AD 뇌에서 정제 또는 체외 인산화 반응에 철저한으로 제조빠른 ggregates보다 효율적으로 26,33-35. 이러한 결과는 P 타우 병리학 적 역할과 매우 잘 일치하고 있습니다. P 타우의 집계에 따라 체외 시스템은 따라서 AD 약물 검사를위한 강력한 도구로 역할을 할 수있다.
타우 집계 및 AD의 진보적 인 신경 퇴행 간의 밀접한 관계뿐만 아니라 아밀로이드 베타 플라크를 대상으로 신약 개발의 최근 실패, AD 36-38의 또 다른 핵심 조직 학적 마커, 타우 집계가 상승 제어 약물 발견에 대한 관심을 감안할 때. 사실, 여러 그룹은 이미 기본 분석 등 체외 타우 집계 반응에 사용하여, 다른 처리량 약물 화면을 시작했다. 화학 물질의 수는 39-42 관내에서 타우 응집에 억제 또는 역전 활동을 나타내는 것으로 밝혀졌다. 그러나, 현재의 모든 타우 집계 레귤레이터 화면 형광체의 주요 병리 마크를 벗어났습니다 수정되지 않은 타우를 사용ylation, AD의 치료에 이러한 화합물을 사용하는 특이성 및 효능에 대한 우려를 제기.
생화학 적 특성화 및 AD 약물 스크리닝 응집 분석법을 개발하는 주요 장애물 중 하나는 중요한 pathophysiologically hyperphosphorylated 타우 단백질의 충분한 양의 제조이다. 타우 1N4R 이성체 및 GSK-3β 키나아제 E.에서 공동 발현되는 지퍼 원조 된 촉매 시스템을 사용하여 대장균은 류신 지퍼 융합 단백질로, 우리는이 도전 극복 (. 수이 등, 제출, 타우와 P 타우의 최종 제품에 대한 그림 1 참조; 또한 P 타우 예비 질량 분석 특성화를 위해 43 참조). 다른 타우 인산화 부위에 대한 특정 항체의 아홉 패널에서 양성 신호가 여덟 위치에서 관찰되었다 (데이타 미기재). 다음, 우리는 통합 운동 D를 차별화 할 수 프로토콜 및 측정 장치를 설명수정되지 않은 타우와 P-타우 종 사이의 ifferences. 이러한 분석은 아밀로이드 (타우 집계)에 따라 thioflavin T (THT) 또는 thioflavin S (THS)의 형광의 증가 (26) 결합을 측정 출판 프로토콜에서 수정되었습니다. 제 "터미널"에서, 무 염색 방법은, 응집 반응은 조립 및 아밀로이드 염료의 부재 하에서 배양 하였다. 상이한 시점에서, 각 반응의 분취 액을 제거하고, 타우 응집체를 바인딩 THT를 응집을 중지하도록 THT - 함유 버퍼의 동량 혼합. 형광은 IAP FluoroMax-2 형광 분석기에 의해 측정된다. "와 염색"두 번째에서 지속적인 모니터링 분석은, THT 또는 THS는 응집 반응에 포함되어 있습니다. 수동 형광 실험 전체에 걸쳐 연속적으로 측정 또는 다중 플레이트 판독기를 사용하여 할 수있다. 또한, 우리는 지속적으로 측정 미주리 집계 타우 및 P 타우 거의 생리적 농도를 사용하는 분석을 설명드. 인산화 효과는 용이하게 검출 가능한 남아있다. 다음, 우리는 단계별 작업 절차를 설명하고 이러한 분석의 대표적인 결과를 보여줍니다. 각 방법의 장단점 일부의 토론뿐만 아니라 잠재적 인 약물 검사 응용 프로그램을 따를 것이다.
고농도, 타우 자발적 아밀로이드 - 유사 구조로 집계. 그러나, 실험실에서, 타우 fibrillization 유도제는 일반적으로 헤파린 (평균 분자량 6,000g / mol) 및 아라키돈 산으로 가속된다. 여기에 기술 된 예는 30 μm의 헤파린을 포함한다. 타우 아밀로이드 응집체의 형성은 thioflavin T (THT) 또는 S (THS) thioflavin 결합 아밀로이드에 의한 형광 모니터링합니다. (:; : 485 nm의 피크 방출 450 nm의 여기) 타우 집계에 결합하면, THT는 형광의 적색 이동을 나타낸다. THS, 다른 한편으로는, 아밀로이드 결합 전의 파장 510 ㎚ (450 nm 파장에서)에서 약한 방출을 가지고 있지만,이 fluorescenc이러한 응집 된 타우 (44)들을 주요 아밀로이드 단백질의 존재하에 예를 커진다. 두 염료는 타우와 P 타우 집계를 감지에서 잘 작동합니다. THT 때문에 (도 2 참조)의 비교적 강한 넓은 발광 피크, 510 nm에서 형광 단위의 30 % 감소가있다. 어느 염료를 사용하는 경우의 편의를 위해, 우리는 타우 응집을 모니터링 여기 / 방출 파장 (즉, 450 nm의 / 510 ㎚)의 동일한 조합을 사용한다.
타우 응집 분석의 목적 및 타우 단백질의 가용성에 따라, 염료의 존재 또는 부재 하에서 수행 될 수있다. 반응의 두 모드는 다음과 같습니다. 단일 샘플 형광 (ISA-SPEX FluoroMax-2) 및 다중 - 플레이트 판독기 (를 SpectraMax M2) - 또한, 우리는 두 개의 다른 기기의 동작을 보여준다. 독자의 특정 요구 및 장비 가용성에 맞게이 프로토콜을 적용 할 수 있어야한다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
시약 1. 준비
- 응집 버퍼 준비 (20 mM 트리스, pH가 7.4, 100 mM의 염화나트륨, 1 mM의 EDTA). 개월 동안 실온에서 안정을 저장합니다. 사용하기 전에 1 mM 디티 오 트레이 톨 (DTT)을 보충합니다.
주 : HEPES 계 완충액 (10 mM HEPES, pH 7.5, 0.1 mM의 EDTA, 5 mM의 DTT)도 타우 응집 유사한 결과를 생성한다. - T thioflavin 0.22 μm의 살균 필터 단위로 S 재고 (집계 완충 용액 3 mM의) 솔루션 및 필터 thioflavin 준비합니다. 달 동안, 알루미늄 호일 안정적인 덮여 튜브에서 -20 ° C에서 보관.
- (300 μM, 집계 완충 용액)을 헤파린 주식 솔루션을 준비합니다. -20 ° C에서 보관, 개월간 안정.
- (물에 용해 1 M) 디티 오 트레이 톨 (DTT) 주식을 준비합니다. 1.5 ml의 튜브로 나누어지는. -20 ° C에서 보관하십시오. 집계 분석하기 전에 실온에서 1 M 솔루션을 해동. 이 1,000 배 주식에서, 탈 이온수로 주식을 작업 100 mm의 나누어지는을 준비합니다. 얼음에 남겨주세요준비가 될 때까지.
- C 냉동고 -80 °에서 타우를 제거합니다. 얼음에 해동. 집계 버퍼와 소정의 농도에 타우를 조정합니다. 4 ° C에서 10 분 동안 20,800 XG에 마이크로 원심의 스핀 미리 형성된 큰 집계를 제거합니다. 이 사전 회전 단계는 단백질 준비의 각 배치의 후속 형광 측정의 일관성을 증가시킨다. 다른 튜브에 뜨는을 전송; 응집 반응을 조립 준비가 될 때까지 얼음에 둡니다.
2. 무 염료, 터미널 분석
주 :이 분석의 응집 반응은 형광 염료의 부재하에 수행된다. 모든 성분을 혼합 한 후, 반응물을 미리 정해진 시간 지점으로 진행시킨다. 분취 량은 집계 반응에서 꺼내 THT 또는 형광 읽기 전에 바인딩 아밀로이드에 대한 THS와 혼합된다. 응집 반응의 초기 부피는 필요한 시점의 수에 의존한다. 이 방법은 REQU 수 있습니다타우 단백질의 다량 반사성이지만 간단, 신속하고, 형광 또는 멀티 웰 플레이트 판독기 (설명 참조)에서 수행 될 수있다. 다음은 단계별 작업이며, 형광 정량 ISA SPEX FluoroMax-2 소형 분광을 사용.
- 표 1에서와 같이 1.5 ml의 에펜 도르프 튜브에 집약 혼합물을 설정. 각 열은 하나의 시간 지점 측정 충분하다 100 μL 반응에 필요한 성분을 나타낸다. 특정 실험에 필요한 시점에 기초하여 전체 집계 믹스 양을 조정한다. 오류를 피펫 팅을위한 공간을 제공하기 위해 각 구성 요소의 추가로 10 %를 추가합니다. 여기에 표시된 전형적인 반응 포함 헤파린은 아라키돈 산 또는 집계 버퍼에 의해 대체 될 수있다. 반응 혼합물에 1 mm로 DTT를 추가합니다. 전체 반응은 하루 이상 지속되는 경우, 환원 환경을 위해 매일 신선한 DTT (1mm)으로 보충.
- 혼합 튜브를 몇 번 반전. 괞 찮아37 ° C 배양기 또는 물 목욕 각각의 반응을 CE. 교반은 타우 집계 필요하지 않습니다.
- 형광을 측정하기 전에 (컴퓨터 다음, 첫 번째 램프) 분광 켭니다.
참고 : 바로 사용할 수있는 크세논 아크 램프. 그러나 최상의 결과를 기계가 형광을 읽기 전에 약 10 분 동안 워밍업 할 수 있습니다. - 컴퓨터의 소프트웨어를 시작합니다.
- 510 nm의 (5 nm의 슬릿)에 인스트루먼트 컨트롤 센터에서 실시간 디스플레이 모드, (2 nm의 슬릿) 450 nm의 설정 여기 파장과 방출 파장을 선택합니다. 닫기 실시간 디스플레이 모드 창은 인스트루먼트 컨트롤 센터로 돌아갑니다.
- 파장 세트를 추가 할 수있는 상부 프레임 정수 파장 분석을 눌러 추가 >> 키를 선택합니다. 1 표준 오차를 설정 인수 매개 변수와 3에 최대 시련, 다음 추가를 클릭합니다. Go!를 클릭하여 데이터 표시를 엽니 다창.
- 데이터 표시 창에서 새 샘플 대화 상자를 열려면 시작 ACQ을 클릭합니다. 샘플 유형에 대해 "알 수없는"를 선택합니다.
- 매 100 μL 집계 혼합물에 98 μL 집계 버퍼와 T. 피펫 여러 번 섞어 3 mM의 thioflavin 2 μl를 추가합니다.
- 큐벳 (FCA3, 외부 차원, wxlxh = 12.5 mm X 12.5 mm X 45mm)에 전체 혼합물을 전송합니다. 샘플 실에서 샘플 홀더에 큐벳을 넣고 뚜껑을 닫습니다. 형광 데이터를 수집하려면 실행을 클릭합니다. 데이터를 기록한다.
- 큐벳을 제거하고 솔루션을 가만히 따르다. 증류수 3 배 큐벳을 씻어. 와 큐벳 외부 공기를 불어 건조.
와 염색 3.를 SpectraMax M2 플레이트 리더에 지속적인 모드 분석
주 :이 분석은 형광 염료 또는 THS THT는 aggrega에 포함된다는 점에서 이전의 것과 다른기 반응. 이 반응의 동일한 세트의 연속 측정을 허용한다. (를 SpectraMax M2의 동작 이하 도시 된 바와 같이) 반응으로 인해 반복 사용으로,이 방법은 더 나은 자동 멀티 웰 플레이트 판독기로 이루어진다. 일반 형광 또한 빠른 작동하지만 응집 반응의 측정 주파수 인해 작업의 설명서 특성상 다소 제한적이다.
- (96 웰 흑색 고체 플레이트, 잘 부피 360 μL, 평바닥) 표 2와 같이 96 웰 플레이트에 응집 믹스를 설정한다. 각 칼럼은 하나로 충분 200 μL 반응에 필요한 성분을 나타낸다 시간 지점 측정. 수회 피펫 팅하여 잘 혼합한다. 실험의 과정을 통해 매일 신선한를 1mM DTT를 보충.
- 37 ° C에서 96 웰 플레이트를 품어.
- 형광 측정 전에 각 시점에서, 멀티 모드 리더와 마이크로 컴퓨터를 켜. 엄마를위한 충분한 시간을 허용차임에는, 약 10 분간 안정.
- 컴퓨터의 소프트웨어를 시작합니다. 37 ° C로 온도를 설정하고 형광 강도 (FI-최고 읽기) 모드, 510 nm에서 450 nm의 방출 파장에서 설정 여기 파장을 선택합니다.
- 서랍에 96 웰 플레이트를 삽입하고 측정을 시작하기 위해 READ 키를 누릅니다.
- 읽은 후, 플레이트를 제거하고 37 ° C 배양기로 다시 돌아갑니다. 데이터 분석 및 플로팅을위한 Excel 스프레드 시트에 데이터를 복사하여 붙여 넣습니다.
4.와 염색, 지속적인 모드 분석 컴팩트 분광에
- 표 3에서 1.5 에펜 도르프 튜브에 집계 혼합물을 설정합니다. 각 열은 한 번에 포인트 측정을 위해 충분하다 200 μL 반응에 필요한 재료를 나타냅니다.
- 혼합 튜브를 몇 번 반전.
- 분광 켜고 단계 2.3 및 2.4에서와 소프트웨어를 설정합니다.
- 전체 혼합 t 이동OA 큐벳. 샘플 실에서 샘플 홀더에 큐벳을 넣고 뚜껑을 닫습니다. 형광 데이터를 수집하려면 실행을 클릭합니다. 데이터를 기록한다.
- 데이터를 실행을 클릭하고 기록하여 적절한 간격으로 계속 읽기. 집계는 고주파수 (예를 들면, 매 30 분 또는 60 초)에서 감시하는 경우, 하나의 측정이 완료 될 때까지 기계 큐벳과의 반응을 그대로 또는 반응 또는 큐벳을 교환하기에 충분한 시간이있을 때.
- 큐벳을 제거하고 솔루션을 가만히 따르다. 증류수 3 배 큐벳을 씻어. 와 큐벳 외부 공기를 불어 건조.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
재조합 타우와 P 타우 (그림 1)을 사용하여, 우리는 단백질 집계를 amyloidal 바인딩에 THT와 THS의 강한 형광 방출의 포함 타우를 활용, 타우 및 P 타우 집계의 반응 속도를 비교하는 두 개의 서로 다른 프로토콜을 설립 및 P 타우 (그림 2). 또는 응집 반응에서 형광 염료없이 우린 과인산 (도 3-5)로 타우 응집 일관된 향상을 관찰 하였다. 이 자극은 헤파린 무관 (데이타 미기재). (도 3 및 5), P-타우 실험 과정을 통해 더 높은 형광을 나타내는 단위로 크게 느려 전에 제 30 분이내 빠른 속도에서 전형적인 반응, 타우 및 p- 타우으로 올리고머에서. 응집 반응에 THT를 포함하는 것은 통합의 속도 (그림 4)에서 상당한 지연이 발생합니다. 두 이소 접근 작은지면반응 후 오 160 시간이 시작했다. THS, 다른 한편으로는, 응집 (도 5)의 상당한 저하가 발생하지 않는다.

샘플은 10 % SDS-PAGE 겔에 의해 해결하고, 쿠마시 블루 (Coomassie blue) R250 (왼쪽)으로 염색 또는 (오른쪽 반 타우 모노클로 날 항체로 프로빙 하였다. 본 연구에서 사용 된도 1 타우 hyperphosphorylated 타우 (p 타우)로 정제하여 패널). 레인 M : 분자량 마커; 레인 1, 3, 인산화 된 타우; lane2 4, hyperphosphorylated 타우.

또는 타우 집계를 결합하지 않고 THT (30 μM) 그림 2. 방출 스펙트럼. 배출 획득했다의 450 nm에서 여기, 600 nm의 (0.1 초 통합, 5 nm의 슬릿 폭 1 nm의 증분)에 대한 460 nm 내지 통조림. 타우 집계는 50 μM 타우 집계가 37 ° CO / N (자세한 내용은 프로토콜 2 참조)에서 진행 할 수 있도록하여 얻었다.
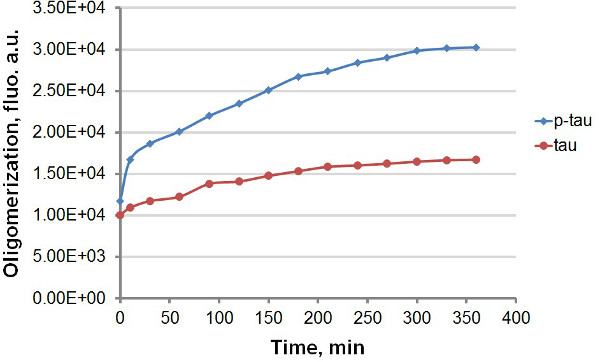
단말기 분석에서 타우와 P 타우 그림 3. 집계 곡선. 집계 50 μM의 타우와 P 타우는 유도 30 μm의 헤파린 완료되었습니다. 반응 개시 후 상이한 시간에, 반응 100 ㎕를 제거하고 형광 측정 전에 60 μM THT의 동량 혼합. 형광을 450 nm의 여기, 발광 파장 510 nm에서 측정 하였다. "AU", 임의의 단위. 시간 척도 분에 있습니다.
"그림 4"SRC = "/ 파일 / ftp_upload / 51537 / 51537fig4highres.jpg"/>
단독 THT 그림 4. 집계 곡선, THT의 존재 하에서 연속 측정 모드 타우 및 p- 타우. 각 반응은 0 또는 50 μM 타우 또는 p- 타우, 30 μM 헤파린 이루어져, 응집 완충액 30 μM THT . 반응은 96 웰 플레이트에서 37 ℃에서 배양 하였다. 상이한 시점에서, 판 배양기로부터 제거하고, 형광 판독 (여기 450 nm에서 510 ㎚)에 대한 플레이트 판독기에 로딩. 판독 사이에, 플레이트 덮개 아래에 동요하지 않고 인큐베이터에 보관되었다. THT의 존재는 크게 수정되지 않은 대응을보다 중요한 것은, hyperphosphorylated 타우는 아직 집계의 빠른 속도를 전시, 집계를 느리게,하지만. 시간의 규모가 시간에 있습니다.
PLOAD / 51537 / 51537fig5highres.jpg "/>
thioflavin가 지시 염료로서 S를도 THS의 존재 하에서 5 소규모 타우 응집 분석법. 6 μM 타우 및 p- 타우 체외 헤파린 - 유도 된 응집 연속 측정 모드에서 평가 하였다. 단백질에 추가하여, 각각의 반응물을 30 μM 헤파린 및 응집 HEPES 완충액 (10 mM의 HEPES pH가 7.5, 5 mM의 DTT, 0.1 mM의 EDTA) 20 μM THS를 함유 하였다. 헤파린을 제외한 모든 성분을 혼합하고, RT에서 평형화 하였다. 헤파린을 첨가 한 후, 반응물을 큐벳에 옮기고 시료 홀더에 배치. 형광 T 0으로 즉시 기록, 약 2 시간 동안 또는 형광 증가가 근처 제로로 둔화 될 때까지 계속되었다. 때문에 비교적 짧은 반응 시간의 전체 반응은 동일한 큐벳에서 실온에서 수행 하였다.
| 타우 | P 타우 | |
| 60-100 μM 타우 | 50 μL | 0 μL |
| 60-100 μM 피 타우 | 0 μL | 50 μL |
| 300 μM 헤파린 | 10 μL | 10 μL |
| 집계 버퍼 | 39 μL | 39 μL |
| 100 mM의 DTT | 1 μL | 1 μL |
표없이 염색 1. 집계 혼합물의 구성 요소, 터미널 분석.
| 타우 | P 타우 | 혼자 염료 | |
| 60-100 μM 타우 | 50 μL | 0 μL | 0 μL |
| 60-100 μM 피 타우 | 0 μL | 50 μL | |
| 300 μM 헤파린 | 20 μL | 20 μL | 20 μL |
| T thioflavin 3 mM의 | 2 μL | 2 μL | 2 μL |
| 집계 버퍼 | 126 μL | 126 μL | 176 μL |
| 100 mM의 DTT | 2 μL | 2 μL | 2 μL |
플레이트 리더에와 염색, 지속적인 분석 표 2. 집계 혼합물의 구성 요소.
| 타우 | P 타우 | 혼자 염료 | |
| 60-100 μM 타우 | 5 μL | 0 μL | 0 μL |
| 60-100 μM 피 타우 | 0 μL | 5 μL | 01, L |
| 300 μM 헤파린 | 20 μL | 20 μL | 20 μL |
| S thioflavin 3 mM의 | 1.5 μL | 1.5 μL | 1.5 μL |
| 집계 버퍼 | 171.5 μL | 171.5 μL | 176.5 μL |
| 100 mM의 DTT | 2 μL | 2 μL | 2 μL |
소형 분광에와 염색, 지속적인 분석 표 3. 집계 혼합물의 구성 요소.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
이 프로토콜은 상이한 분석 조건으로 의존 인산화 타우 응집 빠른 반응 속도를 감지기구를 보여준다. 단말기 분석에서, 형광 염료 THT는 각 시점에서의 마스터 믹스로부터 제거 반응 부에 첨가된다. 아밀로이드 후 26 형광 측정을 유도 결합된다. 두번째 방법으로 염색 모드 타우 응집은 타우 응집체의 성장을 실시간 자동 평가에 적합한 유형의 반응을 렌더링 또는 THT THS의 존재하에 수행된다. 각 방법마다 장단점이 있습니다.
터미널 모드 반응은 타우 집계에 필요한 만 재료를 수행한다. 희석 thioflavin T 반응물을 혼합하여 본질적으로 대폭 형광 정량 반응을 정지 형광 증가 속도를 느리게. 따라서이 방법은 또한 수동 조작과 호환된다. 그러나 reactio 때문에n이 실질적 THT 첨가 종료시, 타우 다량 응집 곡선을 플롯 요구 될 수있다. 이 방법의 또 다른 잠겨진 반응 혼합물에 빈번한 액세스가 미생물 오염이나 단백질 분해 또는 단백질 산화를 도입 할 수 있다는 것이다. 대조적으로 염색 모드 THT 또는 THS의 존재 아밀로이드의 생성을 허용한다. 집계의 발전 적 반응을 방해하지 않고 지속적으로 모니터링 할 수 있습니다. 자동화 된 분석 플랫폼을 구축 할 때이 기능은 특히 매력적이다. 그러나, 다른 염료는 특정 반응을 유도 할 수 있습니다. 실제로, THT 크게 타우 및 P 타우 집계를 지연시킨다, 그러나 THS 거의 효과를 (그림 3과 5를 비교)가 있습니다. PHF 형성을 조직 학적 및 세포 생물학 연구에서 사용되어왔다 thiazines 콩고 레드와 같이 여러 다른 형광 염료가있다. 적어도 하나의 보고서는 이러한 염료 중 일부는 조직의 타우 응집을 유도 할 수 있음을 언급배양 세포 (45). 아밀로이드 역학 연구를위한 화합물을 선택할 때 따라서주의는 연습을해야하고 다른 염료를 비교해야 할 수 있습니다.
악기의 선택과 관련하여, 첫 번째 방법에서 사용되는 단일 샘플 형광 신뢰성이 높은, 그러나보다 약간 반응이 비교 될 때 작업이 번잡 할 수있다. 이 깨지기 쉬운 석영 큐벳의 비용이 일부 금지 수 있지만 여러 큐벳는 반응 사이의 교차 오염을 방지 할 수 있습니다 사용. 반대로, 멀티 웰 마이크로 플레이트 리더는 동시에 여러 반응을 조사 할 수있다. 일회용 96- 웰 플레이트의 사용은 또한 유리하다. 발열체, 마이크로 플레이트 리더는 장시간 동안 37 ° C에서 다중 반응을 모니터링하기위한 전용 장치 일 수있다. 그러나 증발이 문제가 될 수 있습니다. DiNitto는 외. 방지 미네랄 오일과 유사한 반응을 겹쳐증발 (46).
일부주의 사항은 일관되고 정량적 결과를 확인하기 위해 위의 프로토콜을 채취한다. 우선, 타우 및 p- 타우 자발적 특히 고농도로, 시간이 지남에 따라 아밀로이드 응집체를 형성한다. 이는 단백질의 상태로 준비 모든 분취 량을 동결하고, 실험 전에 오직 필요한 양을 해동하는 것이 필수적이다. 그럼에도 불구하고, THT (47)에 의해 검출 세분화 된 중간체 등 일부 집계, 재조합 단백질의 제조 동안 형성되었을 수 있습니다. 전형적인 응집 반응의 실질적인 초기 형광 판독 따라서 일반적이다. 그럼에도 불구하고, 프리 - 방사 공정을 추가하고, 심지어 보이는 단백질 펠렛 않고, 별도의 튜브에 상청을 전송, 감소, 타우 및 p- 타우 준비의 동일한 배치의 일관성, 초기 형광을 유지할 수있다. 둘째, THT의 작업 원액 (즉, 60 μM)는 일하기 전에보다 일주일 동안 실온에서 안정전자 형광 감소. 따라서 그것은 60 μM THT마다 몇 일 다시 확인하는 것이 좋습니다.
P-타우 집계의 연구의 기초가 하나 눈에 띄는 이유는 새로운 AD 진단 및 치료제의 개발이다. 재조합 타우 응집을 억제하거나 되돌릴 화합물은 높은 처리량 화면과 대상 시험 18,40,41,48에서 확인되었다. P 타우 집계 이러한 화합물의 효능이 밝혀지지 남아있다. 이러한 화면은 다른 화합물과의 개별 웰에 다판 염료없이 공통 집합을 혼합하여 분배 단말 모드에서 수행 하였다. 일반적으로 O / N 배양 한 후, THT 또는 THS 많은 화합물의 억제 전력을 공개, 형광 측정에 추가됩니다. 와 염색 방법은 위에서 순위 등 알 언급. 49이 아직 높은 처리량 화면으로 통합 될 수 있습니다. 지금 쌍의 운동 및 제약 연구에 사용할 수 hyperphosphorylated 타우와나선형 필라멘트 형성은 치매 약물 발견 멀리 진행하기 쉽다.
마지막으로, P-타우 응집 연구뿐만 아니라 퇴행성 신경에 매우 중요 할뿐만 아니라 넓은 모집단에도 영향을 미칠 수 있음을 주목할 필요가있다. 예를 들어, 신경 섬유 엉킴은 프로 미식 축구 선수와 권투 선수 50-52 만성 외상성 뇌병증, 일부 환자에서 검출 있다는보고가있다. 비슷한 상관 관계는 군인 (53)를 포함하는 단일 또는 반복적 인 외상성 뇌 손상 환자에 대한보고되었다. 이 작품에 설명 된 프로토콜은 따라서 신경 세포에서 P-타우 집계을 대상으로 새로운 치료제의 발견과 개발을하는 데 도움이 될 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q.
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S.
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M.
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B.
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
