

Summary
חלבוני טאו ללא שינוי וhyperphosphorylated שמשו בשני מבחני צבירה במבחנה כדי לחשוף את קינטיקה הצבירה מהירה hyperphosphorylation תלוי. מבחני אלה יסללו את הדרך למסכים בעתיד לתרכובות שיכולות להשפיע על הנטייה של טאו hyperphosphorylated כדי ליצור סיבים העומדים בבסיס ההתקדמות של מחלת אלצהיימר.
Introduction
מחלת אלצהיימר (AD) היא אחד מאוסף גדול של הפרעות ניווניות הידועות כtauopathies. Tauopathy פתולוגיה שבבסיס המובהק הוא סבכי neurofibrillary, NFTS, בתאי עצב, האסטרוציטים ומיקרוגליה 1-4. צפיפות NFT האזור בקורלציה עם ליקוי קוגנטיבי 3.5 ונוירון הפסד 6. NFT האזור מכיל חלבון בעיקר hyperphosphorylated טאו (המכונה "p-טאו" מעתה ואילך) שיוצר חוטים ישרים או לזווג סליל (PHF) 7,8. טאו הוא חלבון הקשורים microtubule חשב כדי להקל על תחבורת axonal שהיא חיוני לאיתות וסחר 9,10 עצביות. כל מולקולת טאו מכילה 2 עד 3 פוספטים במוח נורמלי, אבל עליות תוכן phosphoryl על ידי כמה קפלים בחולי tauopathy 11. קינאז מרובים עשוי לתרום לhyperphosphorylation טאו כולל GSK3β (3β kinase synthase גליקוגן) וCDK5 (cyclin-dekinase התליון 5) 12,13, אבל ההדק הישיר לזירחון פתולוגי נשאר 14 חמקמקים. זירחון החריג בתוך או ליד המוטיבים מחייב microtubule dissociates טאו מmicrotubule 15, וגורם לטאו אי-לוקליזציה לתא somatodendritic, שבו p-טאו oligomerizes לחוטי סליל ישרים או לזווג שיכול בסופו פלמר לתכלילים NFT אזור. הקשר ההדוק בין hyperphosphorylation טאו, היווצרות NFT אזור, וניוון של מערכת עצבים הוביל להשערה הרווחת כי סבכי p-טאו לעורר תגובה ציטוטוקסיות אפופטוטיים ואחרת, ובכך הוא הגורם הבסיסי לניוון של מערכת עצבי tauopathy 16,17. מסכי סמים ובדיקות קליניות מוקדמות המבוססים על הנחה זו כבר הושקו 18. עם זאת, השערה זו עומדת בפני אתגרים 19,20. לדוגמא, Santacruz et al. הראה כי תפקודים קוגניטיביים של עכברים הטרנסגניים ניתן לשפר על ידי דיכוי הביטוי של מוטציהטאו האנושי, למרות שNFTS המשיך ליצור ממולקולות טאו קיימות 21. במודל דרוזופילה, NFT אזור הוצגו לעקל טאו cytosolic הרעיל כדי להגן על תאי עצב שבבסיס 22,23. ברור, את התפקיד בפתוגנזה של NFT אזור, אם בכלל, ישפיע באופן משמעותי את הכיוון של פיתוח תרופות tauopathy.
בריכוזים גבוהים, חלבון tau המוח רקומביננטי או נורמלי באופן ספונטני אבל לאט לאט polymerizes למבנה כמו PHF-במבחנה, כפי שצוין על ידי הקשירה של כמה צבעי β גיליון העדיפו ניאון, במיקרוסקופ אלקטרונים, וספקטרוסקופיה פיזור אור 24-27. הפרין הוספה או חומצה הארכידונית, חומצת שומן בשפע במוח אנושי, באופן דרסטי מאיצה היווצרות PHF בisoform- טאו ונימוסי ריכוז תלוי inducer 28-32. מעניין לציין, שטאו hyperphosphorylated מטוהר ממוח לספירה או שהוכן על ידי ממצה בתגובות זירחון מבחנהggregates מהיר ויעיל יותר 26,33-35. תוצאות אלו הן בהסכם מצוין עם התפקידים פתולוגית של p-טאו. מערכת במבחנה המבוססת על הצבירה של p-טאו ניתן אפוא לשמש ככלי רב עוצמה להקרנת סמים לספירה.
לאור קשר ההדוק בין צבירת טאו והניוון של מערכת העצבים ההדרגתית של הספירה, כמו גם הכישלון האחרון בפיתוח תרופות מיקוד פלאק Aβ, עוד סמן היסטולוגית מפתח של הספירה 36-38, העניין בגילוי תרופות השולטות צבירת טאו עולה. ואכן, כמה קבוצות כבר החלו מסכי סמים בתפוקה שונה, תוך שימוש בתגובות צבירת טאו מבחנה, כמו assay העיקרי. מספר הכימיקלים נמצא להציג פעילויות מעכבות או היפוך על הצטברות טאו במבחנה 39-42. עם זאת, כל מסכים רגולטור צבירת טאו טאו הנוכחי להשתמש ללא שינוי שמחטיא את מטרת פתולוגי המפתח של זרחןylation, העלאת דאגה לייחודו ויעילותו של שימוש בתרכובות אלו בטיפול לספירה.
אחד המכשולים העיקריים של פיתוח מבחני קיבוץ לאפיון ביוכימי והקרנת הסמים AD הוא הייצור של כמויות מספיקות של חלבון טאו hyperphosphorylated הרלוונטי pathophysiologically. שימוש במערכת קטליזה-Assisted הרוכסנים שבי איזופורם 1N4R של טאו וkinase GSK-3β הם שיתוף לידי ביטוי בE. coli כחלבוני היתוך רוכסן לאוצין, יש לנו להתגבר על האתגר הזה (. סוי et al, הוגש; ראה איור 1 עבור המוצרים הסופיים של טאו וp-טאו; ראה גם 43 לאפיון ספקטרומטריית מסה ראשוני של p-טאו). מהרכב של תשעה נוגדנים ספציפיים לאתרי זירחון שונים של טאו, אותות חיוביים נראו בשמונה עמדות (מידע לא מוצג). בהמשך, אנו מתארים פרוטוקולים וinstrumentations שיכול להבדיל ד הקינטית הצבירהifferences בין טאו ללא שינוי ומיני p-טאו. מבחני אלה שונה מפרוטוקולים שפורסמו שנמדדו העלייה של הקרינה של thioflavin T (THT) או thioflavin S (THS) על עמילואיד (מצרפי טאו) מחייב 26. ב" המסוף "הראשון, גישה לא-צבע, תגובות צבירה הם התאספו וטופחו בהעדר צבע עמילואיד. בנקודות זמן שונות, aliquot של כל תגובה מוסרת ומעורבב עם נפח שווה של חיץ tht המכיל לעצור צבירה ולאפשר tht להיקשר אגרגטים טאו. הקרינה נמדדת על ידי fluorometer IAP FluoroMax-2. במקרה השני "עם צבעים" assay מעקב הרציף, THT או THS כלול בתגובות הצבירה. הקרינה ניתן למדוד באופן רציף לאורך כל הניסוי באופן ידני או באמצעות קורא רב-צלחת. בנוסף, אנו מתארים assay המשתמש בריכוז ליד-פיסיולוגי של טאו וp-טאו לקיבוץ במו המדידה המתמשךדה. ההשפעה של זירחון נשארה לזיהוי בקלות. להלן, נתאר נהלי פעולת צעד-אחר-צעד, ולהראות תוצאות נציג של מבחני אלה. דיון בכמה מהיתרונות והחסרונות של כל גישה, כמו גם יישומי הקרנת סמים פוטנציאליים יבוא בעקבותיו.
בריכוז גבוה, טאו אגרגטים למבנים דמויים-עמילואיד באופן ספונטני. עם זאת, במעבדה, fibrillization טאו מואץ בדרך כלל על ידי מעוררים כגון הפרין (משקל מולקולרי ממוצע, 6,000 g / mol) וחומצה הארכידונית. דוגמאות שמוצגים במסמך זה כוללים 30 מיקרומטר הפרין. ההיווצרות של אגרגטים עמילואיד טאו מנוטרת על ידי הקרינה הנובעת מעמילואיד הכריכה בthioflavin T (THT) או thioflavin S (THS). על כריכה למצרפי טאו, THT מציג משמרת אדומה בקרינה (עירור: 450 nm; פליטת שיא: 485 ננומטר). THS, לעומת זאת, יש פליטה חלשה ב 510 ננומטר (עירור ב 450 ננומטר) לפני עמילואיד מחייב, אבל fluorescenc זהעליות דואר באופן משמעותי בנוכחות של חלבון עמילואיד כגון טאו מצטבר 44. שני הצבעים לעבוד גם באיתור צבירת טאו וp-טאו. בגלל שיא הפליטה החזק ורחב יחסית של tht (ראה איור 2), יש הפחתת% רק 30 ביחידת הקרינה ב 510 ננומטר. לנוחיותכם, אנו משתמשים באותו שילוב של אורכי גל עירור / פליטה (כלומר, 450 nm / 510 ננומטר) כדי לפקח על צבירת טאו בעת שימוש או צבע.
צבירת טאו ניתן לעשות זאת בנוכחות או עדר של הצבע, בהתאם למטרה של assay ואת הזמינות של חלבון טאו. שני המצבים של תגובות מוצגים להלן. בנוסף, אנו מדגימים את פעולתם של שני מכשירים שונים - fluorometer יחיד מדגם (ISA-SPEX FluoroMax-2) וקורא רב-צלחת (SpectraMax M2). קוראים צריכים להיות מסוגלים להתאים את הפרוטוקולים הללו כדי להתאים לצרכימים הספציפיים שלהם וזמינות מכשיר.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. הכנת ריאגנטים
- הכן חיץ צבירה (20 מ"מ טריס, pH 7.4, 100 mM NaCl, 1 mM EDTA). אחסן בRT, יציב במשך חודשים. להשלים dithiothreitol 1 מ"מ (DTT) לפני השימוש.
הערה: חיץ מבוסס HEPES (10 מ"מ HEPES, pH 7.5, 0.1 מ"מ EDTA, 5 מ"מ DTT) גם מייצר תוצאות דומות בצבירת טאו. - הכן thioflavin T או thioflavin פתרון S המניה (3 מ"מ, מומס חיץ צבירה), ולסנן לפי 0.22 מיקרומטר יחידת סינון סטרילי. חנות ב -20 ° C בצינור מכוסה על ידי רדיד אלומיניום, יציב במשך חודשים.
- הכן פתרון מניות הפרין (300 מיקרומטר, מומס חיץ צבירה). חנות ב -20 ° C, יציבה במשך חודשים.
- הכן dithiothreitol מניות (DTT) (1 M, מומסים במים). Aliquot לתוך צינורות 1.5 מיליליטר. חנות ב -20 מעלות צלזיוס. לפני מבחני צבירה, להפשיר את הפתרון 1 M ב RT. מהמניות 1,000x זה, להכין aliquot של 100 מ"מ עובדים המניה עם מים ללא יונים. השאר על קרחעד מוכן.
- הסר טאו ממקפיא -80 מעלות צלזיוס. להפשיר על קרח. התאם טאו לריכוז שנקבע מראש עם חיץ הצבירה. ספין בmicrocentrifuge ב20,800 XG במשך 10 דקות ב 4 ° C כדי להסיר אגרגטים גדולים מראש יצרו. צעד מראש ספינינג זה מגדיל עקביות במדידת הקרינה שלאחר מכן של כל מנה של הכנת החלבון. מעביר את supernatant לצינור אחר; להשאיר על קרח עד מוכן להרכיב את תגובת הצבירה.
2. אין-צבע, Assay Terminal
הערה: תגובת הצבירה של assay זה נעשה בהעדר הצבע פלואורסצנטי. לאחר ערבוב כל הרכיבים, התגובה מותר להמשיך לנקודות זמן שנקבע מראש. אז Aliquots לקוחים מתוך תגובת הצבירה ומעורבבת עם tht או THS לעמילואיד מחייב לפני קריאת הקרינה. הנפח הראשוני של תגובת הצבירה תלויה במספר נקודות זמן הדרוש. גישה זו עשויה requזעם כמות גדולה של חלבון טאו, אבל הוא מהיר, פשוט, וניתן לעשות זאת בfluorometer או קורא צלחת רב היטב (ראה דיון). להלן פעולת צעד-אחר-צעד, באמצעות spectrofluorometer הקומפקטי ISA SPEX FluoroMax-2 לכימות הקרינה.
- הגדר את תערובת הקיבוץ בצינורות 1.5 מיליליטר Eppendorf כמו בטבלה 1. כל עמודה מייצגת את המרכיבים דרושים לתגובת 100 μl, וזה מספיק למדידת נקודת זמן אחת. להתאים את הכמות לתערובת הצבירה כולו מבוססת על נקודות הזמן דרושות לניסוי המסוים. להוסיף 10% נוספים מכל רכיב לתת מקום לpipetting שגיאה. ניתן להחליף הפרין התגובה מכילה הטיפוסי מוצג כאן על ידי חומצה הארכידונית או חיץ צבירה. להוסיף DTT עד 1 מ"מ לתערובת התגובה. אם כל התגובה נמשכת יותר מיום אחד, להשלים יומיומי DTT טרי (1 מ"מ) כדי להבטיח סביבת הפחתה.
- להפוך את הצינור כמה פעמים כדי לערבב. Plaהספירה כל תגובה באמבט חממה או מים 37 מעלות צלזיוס. תסיסה אינה נחוצה לצבירת טאו.
- לפני מדידת הקרינה, להדליק את spectrofluorometer (המנורה ראשונה, ולאחר מכן מחשב).
הערה: מנורת קשת קסנון, שניתן להשתמש מייד. עם זאת, התוצאות הטובות ביותר עבור לאפשר להתקן להתחמם במשך כ -10 דקות לפני קריאת הקרינה. - הפעל את התוכנה במחשב.
- בחר במצב בזמן אמת תצוגה במכשיר Control Center, גל עירור מוגדר 450 ננומטר (חריץ 2 ננומטר) ואורך גל פליטה 510 ננומטר (חריץ 5 ננומטר). מצב חלון תצוגה לסגור בזמן אמת כדי לחזור למרכז בקרת Instrument.
- בחר מפתח אורך גל ניתוח, הוספת עיתונות >> קבוע במסגרת העליונה להוסיף סטי אורכי גל. פרמטרים שנקבעו רכישת שגיאת תקן 1 וניסויים מרביים עד 3, ולאחר מכן לחץ על הוסף. לחץ על Go! כדי לפתוח את תצוגת נתוניםחלון.
- בחלון תצוגת נתונים, לחץ על התחל ACQ כדי לפתוח את תיבת הדו-שיח מדגם החדשה. בחר "לא ידוע" לסוג מדגם.
- לכל תערובת צבירה 100 μl, להוסיף 98 μl חיץ צבירה ו -2 μl 3 מ"מ thioflavin מספר פעמים ט Pipet לערבב.
- העבר את התערובת כולה לקובט (FCA3, ממד חיצוני, wxlxh = 12.5 מ"מ x 12.5 מ"מ x 45 מ"מ). מניחים את קובט בבעל המדגם במדגם-התא ולסגור את המכסה. לחץ על הפעל כדי לאסוף את נתוני הקרינה. רשום את הנתונים.
- הסר את קובט ולמזוג את הפתרון. יש לשטוף את קובט על ידי מים מזוקקים 3 פעמים. יבש על ידי נשיפת אוויר ובמחוץ לקובט.
3. עם צבעים, מתמשך המצב Assay על קורא צלחת SpectraMax M2
הערה: assay זו שונה מהקודמת שבצבע הניאון tht או THS כלול בaggregaתגובת tion. זה מאפשר מדידה רציפה של אותה הקבוצה של תגובות. בשל השימוש החוזר ונשנה של התגובה, שיטה זו טובה יותר לעשות עם קורא צלחת רב גם אוטומטי (כפי שמוצגת להלן הניתוח של SpectraMax M2). Fluorometer רגיל גם עובד, אבל תדירות מדידה של תגובות צבירה מהירות היא מוגבלת למדי, בשל האופי הידני של הפעולה.
- להגדיר את תמהיל הצבירה בצלחת 96-היטב (96-גם צלחת מוצקה שחורה, גם בנפח 360 μl, תחתית שטוחה) כמו בטבלה 2. כל עמודה מייצגת את המרכיבים דרושים לתגובת 200 μl, וזה מספיק לאחד מדידת נקודת הזמן. מערבבים היטב על ידי pipetting מספר פעמים. להשלים טרי 1 המ"מ DTT בכל יום במהלך כל ניסויים.
- דגירה את צלחת 96-גם על 37 מעלות צלזיוס.
- בכל נקודת זמן לפני מדידת הקרינה, להפעיל את קורא microplate מצב מרובה ומחשב. לאפשר מספיק זמן לmachine לייצב, כ -10 דקות.
- הפעל את התוכנה במחשב. להגדיר טמפרטורה ל 37 מעלות צלזיוס ולבחור עוצמת הקרינה מצב (FI-Top קרא), גל עירור להגדיר באורך גל ופליטה 450 ב 510 ננומטר.
- הכנס את צלחת 96-היטב למגירה ולחץ על מקש READ להתחיל מדידה.
- לאחר הקריאה, להסיר את הצלחת ולהחזיר אותו בחזרה לחממה 37 ° C. להעתיק את הנתונים ולהדביק לתוך גיליון אלקטרוני Excel לניתוח נתונים ומזימות.
4. עם צבעים, המתמשך המצב Assay על Spectrofluorometer קומפקטי
- הגדר את תערובת הקיבוץ בצינורות 1.5 מיליליטר Eppendorf כבטבלה 3. כל עמודה מייצגת את המרכיבים דרושים לתגובת 200 μl, וזה מספיק למדידת נקודת זמן אחת.
- להפוך את הצינור כמה פעמים כדי לערבב.
- הפעל את spectrofluorometer ולהגדיר את התוכנה כבצעדים 2.3 ו -2.4.
- להעביר את כל התערובת tקובט OA. מניחים את קובט בבעל המדגם במדגם-התא ולסגור את המכסה. לחץ על הפעל כדי לאסוף את נתוני הקרינה. רשום את הנתונים.
- המשך קריאה במרווחי זמן המתאים על-ידי לחיצה על הפעלה והקלטת נתונים. אם הצבירה היא להיות במעקב בתדירות גבוהה (לדוגמא, בכל שנייה 30 או 60), השאר את התגובה בקובט ובמכונה עד או המדידה סיימה, או כאשר יש מספיק זמן כדי להחליף תגובות או cuvettes.
- הסר את קובט ולמזוג את הפתרון. יש לשטוף את קובט על ידי מים מזוקקים 3 פעמים. יבש על ידי נשיפת אוויר ובמחוץ לקובט.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
באמצעות טאו רקומביננטי וp-טאו (איור 1), שהקמנו שני פרוטוקולים שונים כדי להשוות קינטיקה של צבירה של טאו וp-טאו, מנצל את פליטת הקרינה החזקה של tht וTHS על מחייב לamyloidal אגרגטים חלבון, טאו כולל וp-טאו (איור 2). עם או בלי צבע הניאון בתגובת הצבירה, אנו נצפו שיפור של צבירת טאו עקבי על ידי hyperphosphorylation (איורים 3-5). גירוי זה אינו תלוי בהפרין (מידע לא מוצג). בoligomerize טיפוסי תגובה, טאו וp-טאו בקצב מהיר בתוך 30 דקות הראשונות לפני ההאטה משמעותית (איורים 3 ו -5), עם p-טאו מציג יחידות הקרינה גבוהות יותר בכל מהלך הניסוי. כולל tht בתגובות הצבירה גורם פיגור משמעותי בשיעור של צבירה (איור 4). isoforms שני plat התקרבמי טואלט 160 שעות לאחר התגובות שנכתבו על. THS, לעומת זאת, אינו גורם להאטה ניכרת של צבירה (איור 5).

איור 1. מטוהרי טאו טאו וhyperphosphorylated (p-טאו) שימש במחקר זה. דוגמאות נפתרו על ידי 10% ג'ל SDS-PAGE, ומוכתמות על ידי Coomassie הכחול R250 (משמאל) או נחקרו על ידי נוגדנים חד שבטיים נגד טאו (מימין פנל). ליין M, סמן משקל מולקולרי; נתיבי 1 ו -3, טאו unphosphorylated; lane2 ו -4, טאו hyperphosphorylated.

איור 2. ספקטרום פליטה לtht (30 מיקרומטר) עם או בלי מחייב אגרגטים טאו. היה של רכישת הפליטה שימורים מננומטר 460 עד 600 ננומטר (ננומטר תוספת 1; אינטגרציה 0.1 שניות, 5 רוחב חריץ ננומטר), עירור ב 450 ננומטר. אגרגטים טאו התקבלו על ידי המאפשר צבירת טאו 50 מיקרומטר כדי להמשיך על 37 מעלות CO / N (ראה פרוטוקול 2 לפרטים נוספים).
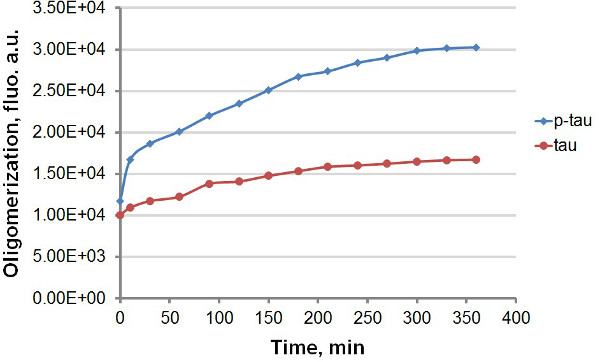
איור 3. עקומות צבירה לטאו וp-טאו בassay המסוף. צבירה של 50 מיקרומטר טאו וp-טאו הושלמו עם 30 מיקרומטר הפרין כinducer. בזמנים שונים לאחר תחילת התגובה, תגובת 100 μl הוסר ומעורבב עם נפח זהה של 60 מיקרומטר tht לפני מדידת הקרינה. הקרינה נמדדה ב 450 ננומטר עירור, 510 פליטת ננומטר. , יחידות שרירותיות "au". שים לב שציר הזמן הוא בדקות.
src = /> "/ קבצים / ftp_upload / 51,537 / 51537fig4highres.jpg" "איור 4"
איור 4. עקומות צבירה לtht לבד, טאו וp-טאו במצב המדידה המתמיד בנוכחות tht. כל תגובה היו מורכבים מטאו 0 או 50 מיקרומטר או p-טאו, 30 מיקרומטר הפרין, ו -30 מיקרומטר tht במאגר צבירה . תגובות הודגרו ב 37 מעלות צלזיוס בצלחת 96-היטב. בנקודות זמן שונות, הצלחת הוסרה מהחממה והעמיסה על צלחת הקורא לקרינת קריאה (ננומטר עירור 450, פליטה 510 ננומטר). בין קריאות, הצלחת נשמרה בחממה ללא תסיסה תחת כיסוי. הנוכחות של tht האטה באופן משמעותי את הצטברות, אבל חשוב מכך, טאו hyperphosphorylated עדיין הציג קצב מהיר יותר של צבירה מאשר מקבילו ללא שינוי. שים לב כי בקנה מידה של הזמן הוא בשעות.
טען / 51,537 / 51537fig5highres.jpg "/>
איור 5. מבחני צבירת טאו בקנה מידה קטנה בנוכחות THS. במבחנה צבירה מושרה הפרין של טאו 6 מיקרומטר וp-טאו הוערך במצב המדידה המתמשך עם thioflavin S כצבע המחוון. בנוסף לחלבון, כל תגובה הכילה 30 מיקרומטר הפרין ו -20 מיקרומטר THS במאגר צבירת HEPES (10 מ"מ HEPES pH 7.5, 5 מ"מ DTT, 0.1 mM EDTA). כל החומרים מלבד הפרין היו מעורבים וequilibrated ב RT. לאחר הוספת הפרין, התגובה הועברה לקובט והניחה בבעל המדגם. הקרינה נרשמה מייד כT 0, ונמשכה כשעה 2 או עד עליית הקרינה האטה כמעט לאפס. בגלל התקופה הקצרה יחסית התגובה, כל התגובה בוצעה ב RT באותו קובט.
| טאו | p-טאו | |
| טאו 100 מיקרומטר - 60 | 50 μl | 0 μl |
| p-טאו 100 מיקרומטר - 60 | 0 μl | 50 μl |
| הפרין 300 מיקרומטר | 10 μl | 10 μl |
| מאגר צבירה | 39 μl | 39 μl |
| 100 מ"מ DTT | 1 μl | 1 μl |
טבלה 1. מרכיבי תערובת הצבירה ללא-צבע, assay מסוף.
| טאו | p-טאו | לצבוע לבד | |
| טאו 100 מיקרומטר - 60 | 50 μl | 0 μl | 0 μl |
| p-טאו 100 מיקרומטר - 60 | 0 μl | 50 μl | |
| הפרין 300 מיקרומטר | 20 μl | 20 μl | 20 μl |
| 3 מ"מ thioflavin T | 2 μl | 2 μl | 2 μl |
| מאגר צבירה | 126 μl | 126 μl | 176 μl |
| 100 מ"מ DTT | 2 μl | 2 μl | 2 μl |
רכיבי 2. תערובת צבירת שולחן, assay המתמשך עם צבעים על קורא צלחת.
| טאו | p-טאו | לצבוע לבד | |
| טאו 100 מיקרומטר - 60 | 5 μl | 0 μl | 0 μl |
| p-טאו 100 מיקרומטר - 60 | 0 μl | 5 μl | 01; l |
| הפרין 300 מיקרומטר | 20 μl | 20 μl | 20 μl |
| 3 מ"מ thioflavin S | 1.5 μl | 1.5 μl | 1.5 μl |
| מאגר צבירה | 171.5 μl | 171.5 μl | 176.5 μl |
| 100 מ"מ DTT | 2 μl | 2 μl | 2 μl |
רכיבי 3. תערובת צבירת שולחן, assay המתמשך עם צבעים על spectrofluorometer קומפקטי.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
פרוטוקול זה מדגים תנאי assay שונים ומכשירים המזהים קינטיקה צבירת טאו המהיר זירחון תלוי. בassay המסוף, צבע הקרינה tht מתווסף לחלק מהתגובה הוסרה מתערובת האמן בכל נקודת זמן. עמילואיד-induced כריכת הקרינה נמדדה 26 אז. במקרה השני, עם צבעי מצב, צבירת טאו מתבצעת בנוכחות tht או THS, טיוח זה סוג של תגובה מתאימה להערכה אוטומטית בזמן אמת של הצמיחה של מצרפי טאו. לכל אחת מהשיטות הללו יתרונות והחסרונות שלה.
תגובת מסוף המצב מתנהלת רק עם אלה מרכיבים הדרושים לצבירת טאו. דילול וערבוב התגובה עם thioflavin T באופן דרסטי מאיטה את קצב גידול הקרינה, במהות עוצר את התגובה לכימות הקרינה. שיטה זו ובכך היא גם תואמת עם תפעול ידני. עם זאת, מכיוון שreaction הוא כמעט נפסק בנוסף tht, כמות גדולה של טאו עשויה להידרש לזוממת עקומת צבירה. עוד הערת אזהרה פוטנציאל לשיטה זו היא שגישה תכופה לתערובת התגובה עשויה להציג את החיידקים או זיהום מפרקי חלבונים, או חמצון חלבון. בניגוד לכך, המצב עם צבעים מאפשר את הדור של עמילואיד בנוכחות tht או THS. קידום הצבירה ניתן לנטר באופן רציף מבלי להפריע לתגובה. תכונה זו היא אטרקטיבית במיוחד בעת הקמת פלטפורמת assay אוטומטית. עם זאת, צבעים שונים עשויים לעורר תגובה מסוימת. ואכן, THT מעכב הצטברות טאו וp-טאו באופן משמעותי, אך יש THS השפעות קטנות (השווה איורים 3 ו -5). יש כמה צבעי הקרינה אחרים, כולל thiazines האדום וקונגו, אשר היו בשימוש במחקרים היסטולוגית וביולוגיה של תא ליצירת PHF. דו"ח אחד לפחות הצהיר כי חלק מהצבעים אלה עלול לגרום להצטברות טאו ברקמהתאי תרבות 45. לכן, בעת בחירת תרכובות אלה ללימודי קינטיקה amyloidogenesis, זהירות תהיה מתורגלת ושצבעים שונים עשויים להיות בהשוואה.
עם כל כבוד לבחירה של מכשיר, fluorometer היחיד המדגם המשמש בגישה הראשונה הוא אמין ביותר, אבל הפעולה עשויה להיות מייגע, כאשר יותר מכמה תגובות הן להיות בהשוואה. באמצעות cuvettes מרובה יכולה לעזור למנוע זיהום צולב בין תגובות, למרות שעלות cuvettes קוורץ השבירה אלה עשויה להיות מונעת מסוימים. לעומת זאת, קורא microplate רב גם יכול לבחון את התגובות מרובות באותו הזמן. השימוש ב96-גם צלחות חד פעמיות הוא יתרון גם כן. עם גוף חימום, קורא microplate יכול להיות מכשיר ייעודי לניטור תגובות מרובות על 37 מעלות צלזיוס במשך תקופה ארוכה של זמן. עם זאת, אידוי עשוי להיות מקור לדאגה. DiNitto et al. מעולף תגובה דומה עם שמן מינרלים כדי למנועאידוי 46.
אמצעי זהירות מסוימים שיש לנקוט לפרוטוקולים לעיל כדי להבטיח תוצאות עקביות וכמותיים. ראשית, טאו וp-טאו באופן ספונטני ליצור אגרגטים עמילואיד לאורך זמן, במיוחד כאשר בריכוז גבוה. זהו אפוא הכרחיים כדי להקפיא את כל aliquots של preps חלבון, ולהפשיר את הנפח הדרוש רק לפני הניסויים. אף על פי כן, כמה אגרגטים, כוללים ביניים פרטניים לזיהוי על ידי tht 47 אולי נוצרו במהלך תקופת ההכנה של חלבונים רקומביננטי. קריאת הקרינה ראשונית ניכרת של תגובת צבירה אופיינית היא כך נפוצה. עם זאת, הוסיף צעד מראש ספינינג והעברת supernatant לצינור נפרד, אפילו בלי גלולה חלבון גלויה, יכול להפחית, ולשמור על הקרינה עולה בקנה אחד, ראשונית מאותו הקבוצה של הכנת טאו וp-טאו. שנית, פתרון מניות העבודה של tht (כלומר, 60 מיקרומטר) הוא יציב בRT עבור פחות משבוע לפני ההקרינה דואר פוחתת. הוא יעץ לכך מחדש לעשות 60 מיקרומטר tht כל כמה ימים.
סיבה בולטת אחת שבסיס מחקרים של צבירת p-טאו היא הפיתוח של כלי אבחון לספירת רומן ותרופות. תרכובות המעכבות או לחזור צבירת טאו רקומביננטי זוהו ממסכי תפוקה גבוהה ובדיקות ממוקדות 18,40,41,48. יעילות של תרכובות אלה לצבירת p-טאו עדיין לא הובהרה. במסכים אלה בוצעו במצב סופני על ידי מחלק תמהיל צבירה משותף ללא צבע לבארות multiplate בודדות עם תרכובות שונות. בדרך כלל לאחר הדגירה O / N, THT או THS מתווסף למדידת הקרינה, חושף את הכח המעכב של תרכובות רבות. הגישה עם צבעים שהוזכרו לעיל ועל ידי אל רנקין ואח. 49 טרם התאגדו למסכי תפוקה גבוהה. כעת, עם טאו hyperphosphorylated זמין למחקרים הקינטית ותרופות של לזווגהיווצרות חוט סליל, גילוי התרופות למחלת אלצהיימר עשויה לקדם רחוק יותר.
לבסוף, ראוי לציין כי המחקר של צבירת p-טאו הוא בעל חשיבות קריטית ללא רק tauopathies, אלא גם יכול להשפיע על אוכלוסייה רחבה עוד יותר. לדוגמא, ישנם דיווחים כי סבכי neurofibrillary הם לגילוי בחלק מהחולים של אנצפלופתיה הטראומטית הכרונית, כגון נגנים מקצועיים אמריקאים כדורגל ומתאגרפי 50-52. מתאם דומה דווח גם לפגיעה מוחית טראומטית בודדת או חוזר על עצמו כולל חיילים 53. פרוטוקולים שתוארו בעבודה זו עשויים לעזור לגילוי ופיתוח התרופות חדשות מיקוד אגרגטים p-טאו בתאים עצביים.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q.
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S.
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M.
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B.
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
