Following co-electroporation of the custom designed PITX3 zinc-finger pair together with the PITX3–EGFP-specific DNA donor vector, and subsequent puromycin selection, positive hESC clones were initially detected via genomic PCR screening (Figure 1). Southern blot hybridization of these PCR positive clones with 5’ and 3’ specific probes confirmed correct targeting to exon 1 of the PITX3 locus (Figure 2), with an efficiency of 19%. Immunofluorescent images shown in Figure 3 demonstrate EGFP expression driven by the PITX3 promoter following differentiation using a published PA6 stromal cell differentiation protocol12,13. Co-localization of EGFP could be observed with markers of midbrain dopaminergic neurons such as FOXA2 (red) (Figure 3A) and tyrosine hydroxylase (TH; red) (Figure 3B), while importantly no co-localization could be observed with the GABAergic marker, gamma-aminobutyric acid (GABA; red) (Figure 3C).
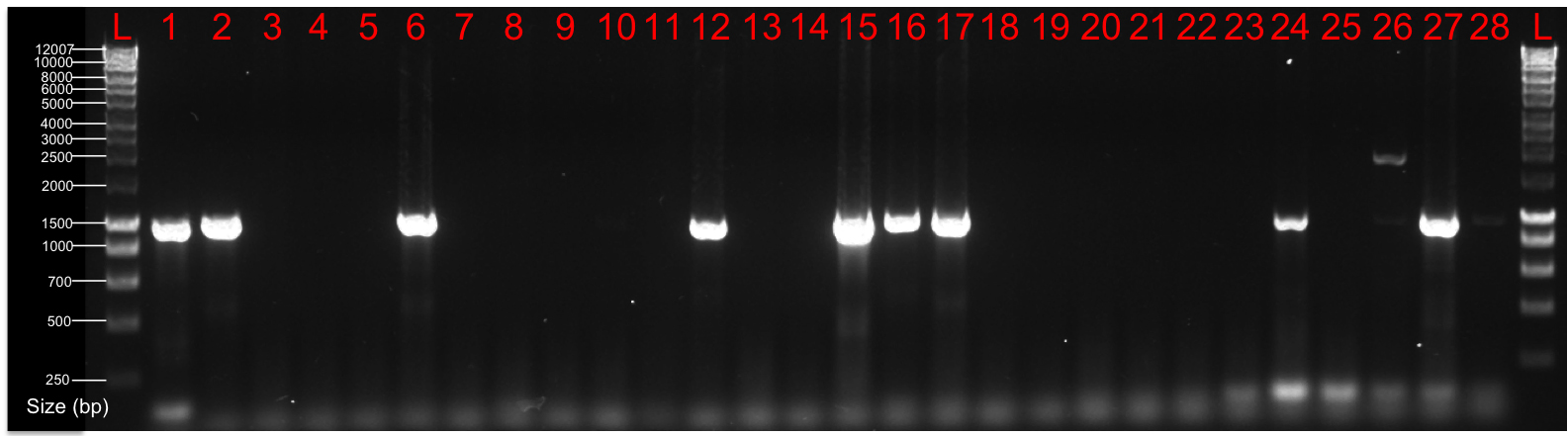
Figure 1. Preliminary genomic PCR screening. Following colony picking and expansion, preliminary genomic PCR screening detected numerous positive clones as shown by the PCR product (band at 984 bp). (Clones were screened using primers hPITX3 L arm gen. F, which lies just outside the left targeting arm, and hPITX3 L arm GFP R which is located within the EGFP gene.) Right-hand and left-hand side lanes are a 1 kb ladder. Please click here to view a larger version of this figure.

Figure 2. Validation of EGFP targeting to the hPITX3 locus using zinc finger nucleases in hESCs. (A) Schematic of the Southern blot strategy employed to validate targeting. The top panel schematic illustrates the native hPITX3 locus before targeting. The bottom panel schematic depicts the hPITX3 locus following correct insertion of the EGFP transgene. The red lines under each panel illustrate the sites at which the 5’ and 3’ Southern blot probes will bind, and correspond to the bands in Figure 2B. (B) Representative Southern blot of correctly targeted colonies selected from PCR pre-screening. Abbreviations: EGFP, enhanced green fluorescent protein; PGK, human phosphoglycerol kinase promoter; PURO, puromycin resistance gene. Other features: loxP sites, purple; polyadenylation sequence, blue, hPITX3 exons, orange. Please click here to view a larger version of this figure.
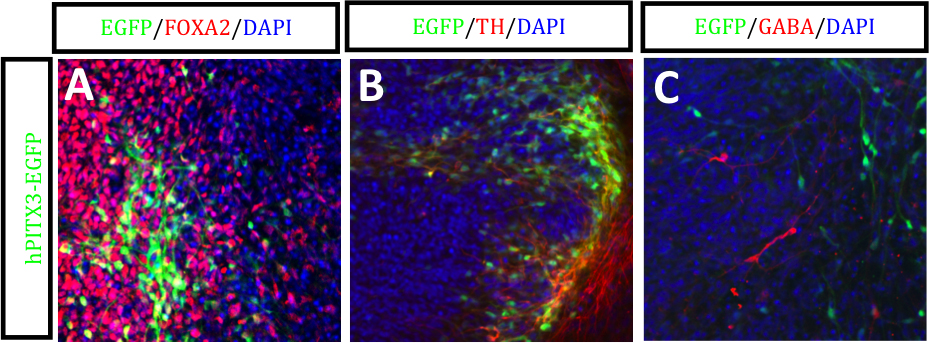
Figure 3. hPITX3-EGFP hESCs differentiated under PA6/LSB/SAG/FGF8 conditions. Immunofluorescent images of EGFP labeled with (A) FOXA2 (red), (B) TH (red) and (C) GABA (red). Counterstained with DAPI (blue). Scales bars, 50 μm. Please click here to view a larger version of this figure.
Following co-electroporation of the custom designed PITX3 zinc-finger pair together with the PITX3–EGFP-specific DNA donor vector, and subsequent puromycin selection, positive hESC clones were initially detected via genomic PCR screening (Figure 1). Southern blot hybridization of these PCR positive clones with 5’ and 3’ specific probes confirmed correct targeting to exon 1 of the PITX3 locus (Figure 2), with an efficiency of 19%. Immunofluorescent images shown in Figure 3 demonstrate EGFP expression driven by the PITX3 promoter following differentiation using a published PA6 stromal cell differentiation protocol12,13. Co-localization of EGFP could be observed with markers of midbrain dopaminergic neurons such as FOXA2 (red) (Figure 3A) and tyrosine hydroxylase (TH; red) (Figure 3B), while importantly no co-localization could be observed with the GABAergic marker, gamma-aminobutyric acid (GABA; red)(Figure 3C).
Figure 1. Preliminary genomic PCR screening. Following colony picking and expansion, preliminary genomic PCR screening detected numerous positive clones as shown by the PCR product (band at 984 bp). (Clones were screened using primers hPITX3 L arm gen. F, which lies just outside the left targeting arm, and hPITX3 L arm GFP R which is located within the EGFP gene.) Right-hand and left-hand side lanes are a 1 kb ladder. Please click here to view a larger version of this figure.
Figure 2. Validation of EGFP targeting to the hPITX3 locus using zinc finger nucleases in hESCs. (A) Schematic of the Southern blot strategy employed to validate targeting. The top panel schematic illustrates the native hPITX3 locus before targeting. The bottom panel schematic depicts the hPITX3 locus following correct insertion of the EGFP transgene. The red lines under each panel illustrate the sites at which the 5’ and 3’ Southern blot probes will bind, and correspond to the bands in Figure 2B. (B) Representative Southern blot of correctly targeted colonies selected from PCR pre-screening. Abbreviations: EGFP, enhanced green fluorescent protein; PGK, human phosphoglycerol kinase promoter; PURO, puromycin resistance gene. Other features: loxP sites, purple; polyadenylation sequence, blue, hPITX3 exons, orange. Please click here to view a larger version of this figure.
Figure 3. hPITX3-EGFP hESCs differentiated under PA6/LSB/SAG/FGF8 conditions. Immunofluorescent images of EGFP labeled with (A) FOXA2 (red), (B) TH (red) and (C) GABA (red). Counterstained with DAPI (blue). Scales bars, 50 μm. Please click here to view a larger version of this figure.
| Substance | ml/100 ml | Catalogue No. (Supplier) |
| DMEM | 89 | 10566-016 (Life Technologies) |
| Fetal Bovine Serum | 10 | 16140-071 (Life Technologies) |
| Pen/Strep | 1 | 15070-063 (Life Technologies) |
Table 1.
| Component | 100 mm dish | 60 mm dish | 6-well | 48-well | 96-well | 75cm2 flask |
| HBSS | 5 ml | 2 ml | 1 ml | 0.5 ml | 0.1 ml | 8 ml |
| Accutase | 5 ml | 2 ml | 1 ml | 0.5 ml | 0.1 ml | 8 ml |
| Dispase | 5 ml | 2 ml | 1 ml | – | – | – |
| hESC media | 10 ml | 6 ml | 3 ml | 0.5 ml | 0.2 ml | 15 ml |
| Matrigel | – | – | – | – | 0.1ml | – |
Table 2.
| Substance | ml/100 ml | Catalogue No. (Supplier) |
| DMEM/F12 | 80 | 11320-033 (Life Technologies) |
| KSR | 20 | 10828-028 (Life Technologies) |
| NEAA | 1 | 11140-050 (Life Technologies) |
| GlutaMAX | 1 | 35050-061 (Life Technologies) |
| Pen/Strep | 1 | 15070-063 (Life Technologies) |
| ß-mercaptoethanol | 0.1 | 21985-023 (Sigma-Aldrich) |
| rhFGF2 | 7 ng/ml [final] | 233-FB-025/CF (R&D Systems) |
Table 3.
| Substance | Concentration | Catalogue No. (Supplier) |
| N-Lauroylsarcosine sodium salt solution (Sarcosyl) | 0.5% (w/v) | 61747 (Sigma-Aldrich) |
| EDTA | 10.0 mM | E9884 (Sigma-Aldrich) |
| NaCl | 10.0 mM | S3014 (Sigma-Aldrich) |
| Tris-Cl (pH7.5) | 10.0 mM | T3253 (Sigma-Aldrich) |
| Proteinase K | 1.0 mg/ml | BIO-37084 (Bioline Australia) |
Table 4.
| Substance | Concentration | Catalogue No. (Supplier) |
| Tris-Cl (pH8.0) | 10.0 mM | T3253 (Sigma-Aldrich) |
| EDTA (pH8.0) | 0.1 mM | E9884 (Sigma-Aldrich) |
Table 5.
