The present protocol is a modified digestion protocol published by Link et al., 20076 with a shorter digestion time (45 min maximum) due to mechanical disaggregation with an automated multichannel pipette. In addition, the procedure is more standardized, minimizes degradation of surface markers on different lymph node stromal cells and allows the handling of more than one sample at the same time.
Collagenase IV and Collagenase D in Links protocol6 and current protocol or Collagenase P and Dispase in Fletchers protocol13 maintain lymph node stromal cells viability and spare the digestion of several surface markers including CD45, gp38 and CD31. To test the current protocol, axillary, brachial and inguinal lymph nodes were removed and digested to isolate their stromal cells. Analysis of CD45, gp38 and CD31 expression demonstrated the presence of TRC, LEC, BEC and DN cells isolated from C57BL/6 mice (Figure 1, gating strategy).
Mechanical stress might reduce the viability of lymph node stromal cells during digestion. Comparing the viability of isolated subpopulations of stromal cells using the three different protocols, it was found that the viability was higher than 95% in the Fletcher protocol, but lower for both Current protocol and Link protocol (Figure 2A), although Current protocol show better survival in the BEC subpopulation then Link protocol. The total cells number recovered post digestion of the lymph node stromal cell subsets was slightly higher in Current protocol compared to Link protocol and Fletcher protocol (Figure 2A). These results demonstrate that the Current protocol maintains viability of lymph node stromal cells similar to published protocols. The major differences between the three protocols are listed in Table 1.
Surface markers are required to characterize lymph node stromal cells by flow cytometry and to isolate them by cell sorting. TRCs and LECs isolated via Current, Link and Fletcher protocols were compared for I-Ab, CD140a, CD80, PD-L1 and CD40 expression. After isolating stromal cells by all three protocols, we found the expression of I-Ab, CD80, CD140a, PD-L1, and CD40 was higher upon digestion with CP and LP in both TRCs and LECs (Figure 2B). These results suggest that degradation of some surface molecules with Collagenase IV and D are less strong than with Collagenase P and Dispase.
Taken together, the current protocol includes a short digestion combined with automated mechanical disaggregation to minimize surface marker degradation of viable lymph node stromal cells.

Figure 1. TRC, LEC, BEC, and DN cells staining, gating, and quantification. Lymph nodes from C57BL/6 mice were digested following the current protocol and stained with CD45, gp38 and CD31, Live/Dead to define TRC, LEC, BEC, and DN cells by flow cytometry. Data show representative results of the staining. Click here to view larger image.
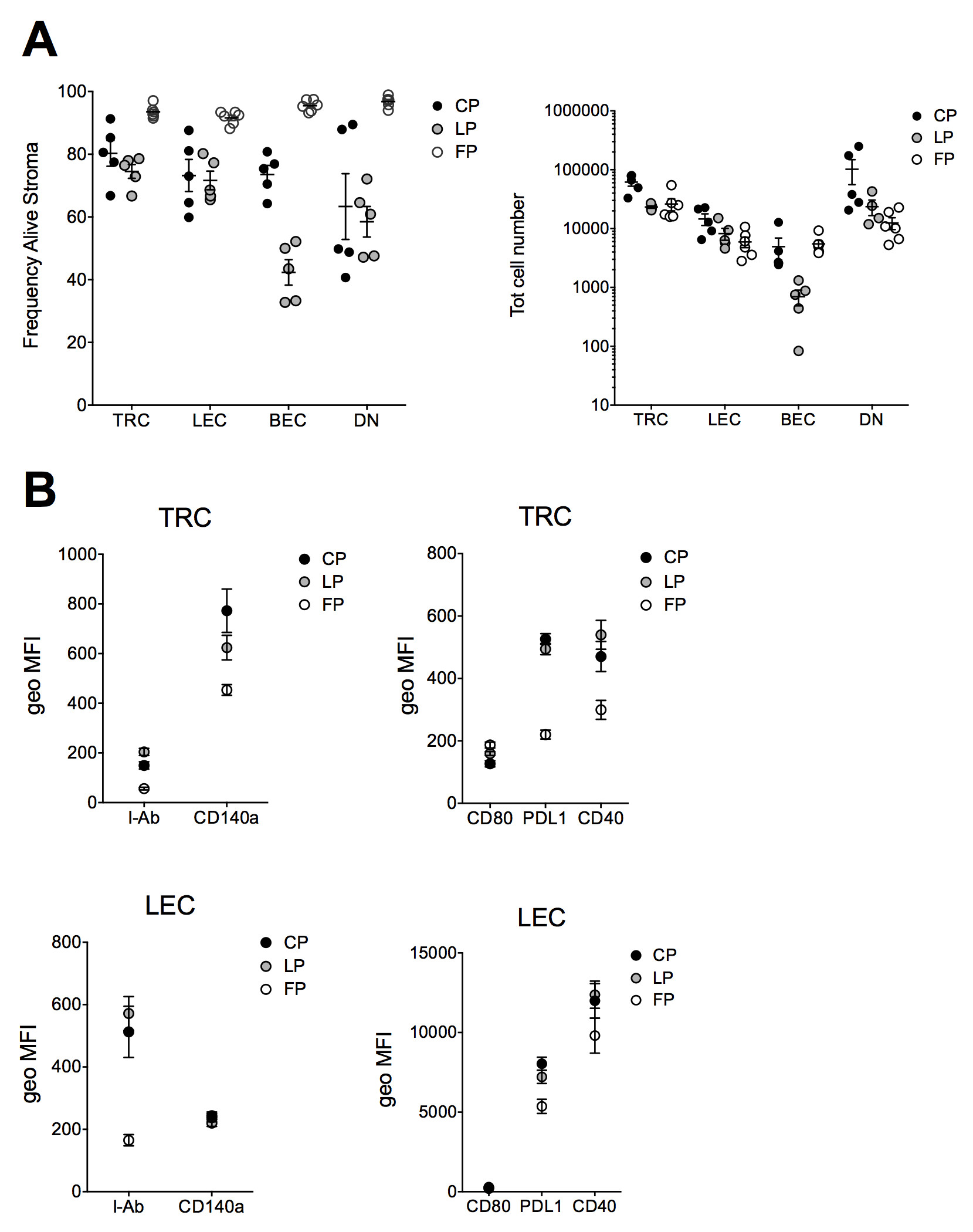
Figure 2. Viability and surface marker expression of lymph node stromal cells. Lymph nodes from C57BL/6 mice were digested following the Current (CP), Link’s (LP) and Fletcher’s protocol (FP) and stained with CD45, gp38 and CD31 to define TRC, LEC, BEC and DN cells by flow cytometry. A) Viability and total cell number of all lymph node stromal cell subpopulations after Live/Dead staining (n ≥ 5, pooled data of 2 independent experiments, bars represent SEM). B) Analysis of surface expression of I-Ab (APC-Cy7), CD140a (PerCP-Cy5.5), CD80 (PerCP-Cy5.5), PD-L1 (PE-Cy7) and CD40 (PE-Cy7) on TRCs and LECs (n = 6 for FP and n = 11 for LP and CP, pooled data of 2 independent experiments, bars represent SEM). Geometric MFI auto-fluorescence values for LEC-APC-Cy7: 324 ± 97, LEC-PECy7: 163 ± 82.5, LEC-PerCpCy5.5: 156 ± 36, TRC-APC-Cy7: 349 ± 152, TRC-PECy7: 117 ± 54, TRC-PerCP-Cy5.5: 121 ± 45. Click here to view larger image.
| Current Protocol-Enzymes | Current-protocol | Link Protocol-enzymes | Link-protocol | Fletcher Protocol-enzymes | Fletcher-protocol |
| Collagenase IV, Dnase I | 30 min, 37 °C, stirring | Collagenase IV, DNAse I | 30 min, 37 °C | Collagenase P, Dispase, DNAse I | 20 min, 37 °C, invert every 5 min |
| Collagenase D, Dnase I | 5 min, 37 °C, stirring, resuspend | Collagenase D, DNAse I | 20 min, 37 °C | Collagenase P, Dispase, DNAse I | 10 min, 37 °C, resuspend 30 sec |
| 10 min, 37 °C, stirring | Collagenase D, DNAse I | 37 °C, resuspend every 10 min until complete digestion | Collagenase P, Dispase, DNAse I | 37 ° C, resuspend every 5 min until complete digestion | |
| automated mechanical resuspension |
Table 1. Short summaries of CP, LP, FP enzymes used and protocols.
