Optogenetic Stimulation of the Auditory Nerve
Summary
Cochlear implants (CIs) enable hearing by direct electrical stimulation of the auditory nerve. However, poor frequency and intensity resolution limits the quality of hearing with CIs. Here we describe optogenetic stimulation of the auditory nerve in mice as an alternative strategy for auditory research and developing future CIs.
Abstract
Direct electrical stimulation of spiral ganglion neurons (SGNs) by cochlear implants (CIs) enables open speech comprehension in the majority of implanted deaf subjects1-6. Nonetheless, sound coding with current CIs has poor frequency and intensity resolution due to broad current spread from each electrode contact activating a large number of SGNs along the tonotopic axis of the cochlea7-9. Optical stimulation is proposed as an alternative to electrical stimulation that promises spatially more confined activation of SGNs and, hence, higher frequency resolution of coding. In recent years, direct infrared illumination of the cochlea has been used to evoke responses in the auditory nerve10. Nevertheless it requires higher energies than electrical stimulation10,11 and uncertainty remains as to the underlying mechanism12. Here we describe a method based on optogenetics to stimulate SGNs with low intensity blue light, using transgenic mice with neuronal expression of channelrhodopsin 2 (ChR2)13 or virus-mediated expression of the ChR2-variant CatCh14. We used micro-light emitting diodes (µLEDs) and fiber-coupled lasers to stimulate ChR2-expressing SGNs through a small artificial opening (cochleostomy) or the round window. We assayed the responses by scalp recordings of light-evoked potentials (optogenetic auditory brainstem response: oABR) or by microelectrode recordings from the auditory pathway and compared them with acoustic and electrical stimulation.
Introduction
According to the World Health Organization, 360 million people worldwide suffer from hearing loss. In deaf subjects, direct electrical stimulation of SGNs by CIs enable open speech comprehension in the majority of them1,2,4,5. Even though CIs have been implanted in more than 200,000 people, therefore being the most successful neuroprosthesis, sound encoding driven by the current cochlear implants is limited. CIs are based on electrical stimulation by a certain number of electrodes where each one activates a tonotopic region of the auditory nerve thus bypassing the dysfunctional sensory organ of Corti in the cochlea. Normal hearing listeners can discriminate more than 2,000 frequencies, however today's CIs use only up to 12-22 frequency channels4. This is due to widespread current flow from each stimulating electrode7,9, activating a large number of SGNs that represent many different sound frequencies8,15. This limitation can be improved using multipolar stimulation but at the expense of higher power consumption16,17. Their output dynamic range for sound intensity is also limited, typically below 6-20 dB4,18. For these reasons, improving frequency and intensity resolution are important objectives for increasing CI performance to ameliorate speech recognition in noisy environments, prosody comprehension and music perception.
A different option to stimulate the auditory nerve is optical stimulation. Light can be conveniently focused to target a small SGN population, promising better spatial confinement, increasing frequency resolution and also widening dynamic range, resulting in a better intensity resolution. Indeed, cochlear stimulation with infrared light has shown excellent frequency resolution in animal models10,11,19. One disadvantage of this kind of stimulation is that it requires higher energies than electrical stimulation10,11. Moreover, concerns about the ability of the method to directly stimulate auditory neurons have been raised12,20.
As an alternative to infrared stimulation, we employ optogenetics to render SGNs light sensitive. Optogenetics is a novel approach that combines genetic and optical techniques to non-invasively and specifically control cells with high temporal precision (reviews21-23). The currently most frequently used modality employs the expression of the microbial channelrhodopsin 2 (ChR2) gene of Chlamydomonas reinhardtii and variants thereof, encoding a light-gated cation channel24. ChR2 is a 7-transmembrane-helix protein that, when transduced into neurons and activated by blue light, acts as non-selective cation channel, thus depolarizing the cells24-27. ChR2 has been well characterized24,28-31 and many variants have been developed to modify action spectrum, gating and permeability properties32,33. The aim of our work is to establish cochlear optogenetics for the activation of the auditory pathway. We note that the optogenetic approach to stimulate the auditory nerve requires genetic manipulation of the spiral ganglion for the expression of channelrhodopsin. Working with mice and rats allows the use of available transgenic animals13,34,35, which provide expression of channelrhodopsin with little variability along the tonotopic axis and across animals36. Combining conditional alleles37 with appropriate Cre-lines provides for cell-specific expression. Gene transfer into the spiral ganglion of other animals requires the use of virus such as adeno-associated virus that is a standard approach in optogenetics38 and that we showed to work well in mice36. Genetic manipulation and expression of transgenes encoding alien proteins bear risks for adverse effects such as immune responses and/or proliferation, compromised condition or even death of genetically manipulated cells. For the purpose of this demonstration we use transgenic mice expressing ChR2 in spiral ganglion neurons under the Thy-1 promotor13 to optically stimulate the auditory pathway. We note that other channelrhodopsin variants can be used for the same purpose as we demonstrated using virus-mediated transfer of the variant CatCh14 into SGNs39.
While cochlear optogenetics requires genetic manipulation, it offers molecular tuning for optimized SGN stimulation and promises improved frequency and intensity resolution when compared to electrical stimulation. Optogenetic stimulation of the auditory pathway is highly relevant for hearing research. For example, it promises advances in studies of the activity-dependent refinement of tonotopy during development, in the analysis of the requirement for spectral integration in sound localization and of the extent of interaction between frequency-specific afferent projections in the central auditory system.
Protocol
All experiments presented in this work were conducted with the ethical standards defined by the German law for the protection of experimental animals. The University of Goettingen board for animal welfare and the animal welfare office of the state of Lower Saxony approved the experiments.
1. Preparation of µLED-stimulator
- For µLEDs, first prepare the µLED-stimulator. Use blue LEDs with 200 by 200 μm active surface (µLED, see Materials Table).
- Solder wires to the µLED. Then, encapsulate the μLED and the connections using epoxy glue. Leave the device overnight to let the epoxy cure.
- For mechanical stability and electrical insulation funnel the wires through a thin glass capillary, use 1 mm outer diameter borosilicate capillaries and seal the junction with epoxy (Figure 1D) such that the capillary can be mounted on a mechanical manipulator.
NOTE: There is a trade-off of making the capsule thick enough to get a good electrical isolation (avoiding stimulation artifacts), but thin enough to let the LED move freely inside the bulla and to locate it onto the cochleostomy. It is a good practice to prepare more than one LED. - To confirm good electrical isolation, first prepare a Petri dish with 0.9% NaCl and dip 3 bare ended copper wires into the liquid. This serves to simulate an animal with ABR electrodes. Connect the wires to the ABR-amplifier and from there to an oscilloscope.
- Then, connect the LED to the stimulator and immerse it into the solution until it is totally covered. Drive the LED with pulses (see step 3.1.1) at the maximum allowed current for longer than 20 min and check for stimulation artifacts. If they appear, discard the LED and try a new one.
2. Surgical Procedure
- Use 4-10 week-old transgenic mice expressing ChR2 under the Thy1.2 promoter (line 9 in Wang et al.13). To manipulate the animal and perform the surgery, always use clean and well identified surgical tools.
- Anesthetize animals with an i.p. injection of a mixture of urethane (1.32 mg/kg), xylazine (5 mg/kg), and buprenorphine 0.1 µg/g diluted in 0.9% sterile NaCl solution and place the mouse onto a heating plate for maintaining body temperature at 37 °C. With this dose, anesthesia typically lasts for the whole experimental protocol.
- Beginning 10 min after initial anesthesia induction check for signs of arousal by the paw withdrawal reflex. In case of movements, apply maintenance doses of urethane (.66 mg/kg) and buprenorphine (0.05 µg/g) diluted in 0.9% sterile NaCl solution (without Xylazine). Again check the paw withdrawal reflex and proceed only if the reflex is absent.
- Carefully shave the retroauricular area in preparation of the incision. Use a retroauricular approach to reach the bulla (air-containing middle ear cavity).
- With a micro forceps carefully dissect and/or remove part of the neck muscles, e.g., platysma, sternocleidomastoid, and scalenes.
- Find and expose the bulla (Figure 1A). Identify the bulla as a bony structure with a characteristic spherical shape with a ring on the bulla surface that indicates the insertion of the tympanic membrane (Figure 1A). Make a hole with the tip of an insulin needle just below this ring. Start there, remove the bone that covers the bulla with a fine Rongeur (gnawer) or microforceps in such a way that the promontorium of the cochlea is exposed (Figure 1B).
- For the cochleostomy, gently shave off the bony capsule to open a small window (cochleostomy: ~500-800 μm), taking care to leave intact the membranous labyrinth (Figure 1B). Perform the cochleostomy on the second cochlear turn as it is far enough from the stapedial artery and from the apex. Opening the apex could produce rupture of the cochlea including a fracture of the modiolus.
- For a round window insertion of an optical fiber or µLED-implant to scala tympani, carefully enlarge the round window niche by scratching the bony niche with the tip of a sharp scalpel (a blade number 11 is a good option). Always use a new blade or scalpel. Be very careful as the stapedial artery runs very close to the round window (Figure 1B).
3. Optical Stimulation Using Two Approaches
- For transcochlear stimulation, use µLEDs or a fiber coupled laser.
- Mount the capillary carrying the µLED on a mechanical manipulator and carefully position the µLED onto the cochleostomy. Use oABR, elicited by 3-10 msec long stimulation from a current source (typically 5-40 mA at 2.7 V) at 1-5 Hz, as a means to optimize the position and orientation of the µLED.
- For laser stimulation, use a 473 nm continuous wave laser and a 250 µm optical fiber (see Materials). Couple the optical fiber to a blue laser source that ideally offers fast analogue control of power (else use an acousto-optical device or a fast shutter to define the optical stimulus). Using a micromanipulator, locate the tip of the fiber directly onto the cochleostomy and fix it there using dental cement (transcochlear stimulation).
- For intracochlear stimulation, insert the fiber through the round window into the scala tympani. Then use 3-10 msec laser pulses of 10-30 mW (power of fiber output) at 1-5 Hz to elicit oABR. The large amplitude of oABR enables the experimenter to observe oABR evoked by single stimuli on the oscilloscope for optimizing the position and orientation of the emitter (µLED or fiber) using oABR amplitude as a read-out.
- Once good oABR have been achieved, fix the fiber with cyanoacrylate glue or dental cement and wait until the glue is solid.
NOTE: It is normal to observe some liquid coming out from the cochlea. However to avoid problems with the polymerization of the cement, it is advisable to dry up the region using fine cotton wicks.
4. Recordings of oABR
- Insert needle electrodes underneath the pinna, on the vertex and on the back near the legs and connect them to the amplifier.
- Amplify the difference potential between vertex and mastoid subdermal needles. Sample at a rate of 50 kHz and filter (1-10,000 Hz). Visualize the difference potential on an oscilloscope ideally near the recording setup for optimizing the position of the optical stimulator. Use signal averaging (50-1,000x) and store the data on a computer for offline analysis.
5. Recordings of Optically Evoked Local Field Potentials
- Place the animal into a stereotactic frame. Using forceps centrally pull the skin overlying the cranium and perform a median incision with scissors roughly extending from between the eyes to the point where the neck muscles attach to the skull.
- Reflect the skin laterally. Cut the periosteum lengthwise along the sagittal suture and remove it sideward by gently rubbing the exposed cranial bone with a cotton swab.
- Position a custom made metal plate onto the exposed skull leaving approximately 1 mm space between the caudal end of the plate and lambda. With a 0.7 mm diameter drill carefully drill a hole into the parietal bone.
- Gently screw in a 0.85 mm diameter screw – to serve as reference for potentials recorded from the inferior colliculus (IC) – holding the screw with forceps as long as the screw is not securely fastened. Ensure good contact between screw and dura without penetrating the brain. Glue the screw and the metal plate onto the skull with cyanoacrylate glue.
- Loosen the neck muscles gently from the skull. Pull the neck muscles caudally for circa 1-2 mm. Identify the intersection of the superior sagittal sinus (SSS) and the transverse sinus (TS). If necessary, increase the transparency of the skull by moistening the bone with physiological NaCl solution.
- Identify the IC in mice as lying directly caudal to the SSS and TS intersection. After identification of the location of the IC slowly and carefully perform a trepanation extending ca. 0.5 mm rostrally to 1.5 mm caudally to the TS and roughly 3 mm laterally.
- Avoid any injury to the SSS and TS as these would preclude a successful experiment. Here, test whether the trepanned bone has become loose by softly pushing it. Using forceps slowly lift off the bone.
- With an appropriate adaptor attach a multichannel array and headstage onto a micromanipulator. To prevent destroying the multichannel recording array during insertion into the IC, carefully remove the dura with a bent tip of a small hypodermic needle starting close to a bone edge.
- Remove the animal from the stereotaxic frame. Fix the metal plate onto a bar to remain stereotaxic fixation. Position the recording probe over the IC and insert it under microscopic control such that the top most channel is just visible at the surface.
- Amplify the difference potential between the individual channels of the recording array and the reference screw in the parietal bone, sample at a rate of 32 kHz, low-pass filter (300 Hz) and store on a computer for offline analysis.
- All animals should be humanely euthanized at the end of the procedure prior to anesthetic recovery in accordance with relevant euthanasia guidelines. Suitable methods of euthanasia may include cervical dislocation or CO2 inhalation.
Representative Results
An optimal cochleostomy is critical and increases the probability of a successful experiment. This means the window is regular, small, and there is no injury of the internal cochlear structures. For example, bleeding indicates damage of the stria vascularis. A good example is presented in Figure 1B.
Using ChR2-transgenic mice, ChR2 is expressed in the SGNs within the cochlea (Figure 1C). Blue light illumination, either by µLED or laser, elicits large oABR, which differ from acoustic ABR (aABR) in amplitude and waveform. oABR (Figure 1F) have larger amplitudes than aABR (Figure 1E) but they are comparable to electrical ABR (eABR, Figure 1G). This can be interpreted as oABR and eABR reflecting recruitment of more SGNs and/or a more synchronized activation of SGNs than for sound-evoked aABR.
To elicit eABR a rodent electrical CI (MED-EL) was inserted into the round window. We used biphasic current pulses (80 μsec phase duration, 20 μsec interphase duration, 500 μA, 6 Hz stimulation rate) to stimulate and averaged responses to 100 stimulus repetitions. Stimulation with 2 glass-insulated tungsten electrodes (1 inside and 1 outside of the cochlea) is also possible.
It is important to note that oABR are not detectable in wild type animals, after inhibition of voltage-gated sodium channels or after sacrificing the animals39. These control experiments render heating as a mechanism underlying optically evoked ABRs unlikely.
We verified propagation of the activity through the auditory pathway by extracellular recordings in the central auditory system. For an example, optogenetic stimulation of the cochlea elicited also neural activity in the auditory midbrain (inferior colliculus, Figure 2).
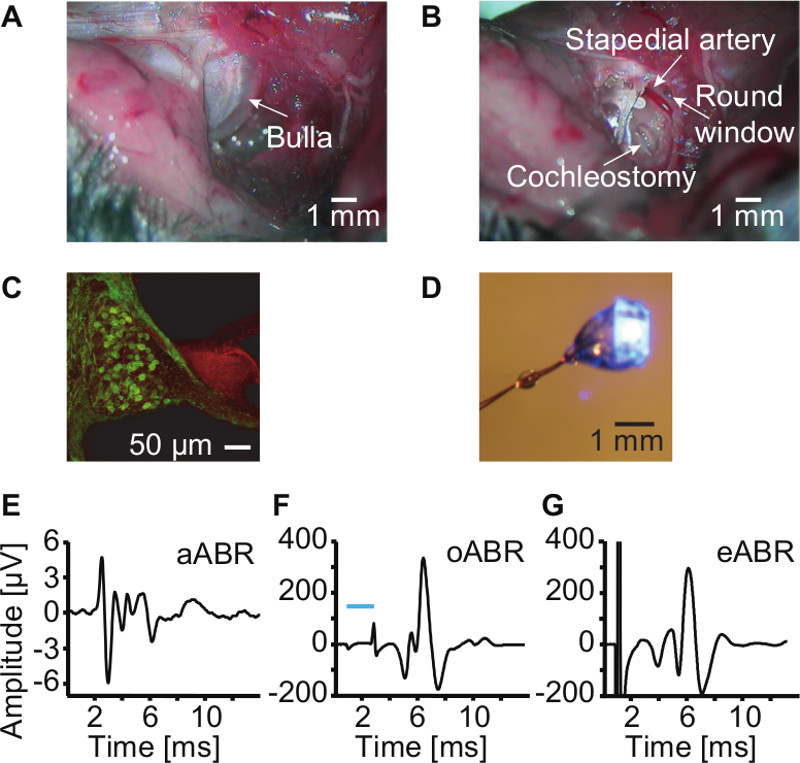
Figure 1: Surgical procedure, µLED preparation, and oABR recording. (A) Dissecting the neck muscles exposes the bulla; observe its characteristic spherical shape and the ring where the eardrum inserts into the bone, which can be used as a landmark. Scale bar 1 mm. (B) Opening of the bulla will expose the promontorium. The cochleostomy was performed on the second cochlear turn. Other landmarks are indicated, among them the stapedial artery. Scale bar 1 mm. (C) ChR2-YFP expression in SGNs. Immunolabeling for GFP (green) and phalloidin-AF-568 (red). Scale bar 50 µm. (D) µLED, wired but not yet funneled into the capillary. Scale bar 1 mm. (E), (F), and (G) representative examples of aABR (click, 20 Hz, 80 dB), oABR (2 msec, 1 Hz, 4 mW/mm2) induced by LED stimulation and eABR (900 μA, 20 Hz) via glass-insulated tungsten electrodes. Please click here to view a larger version of this figure.
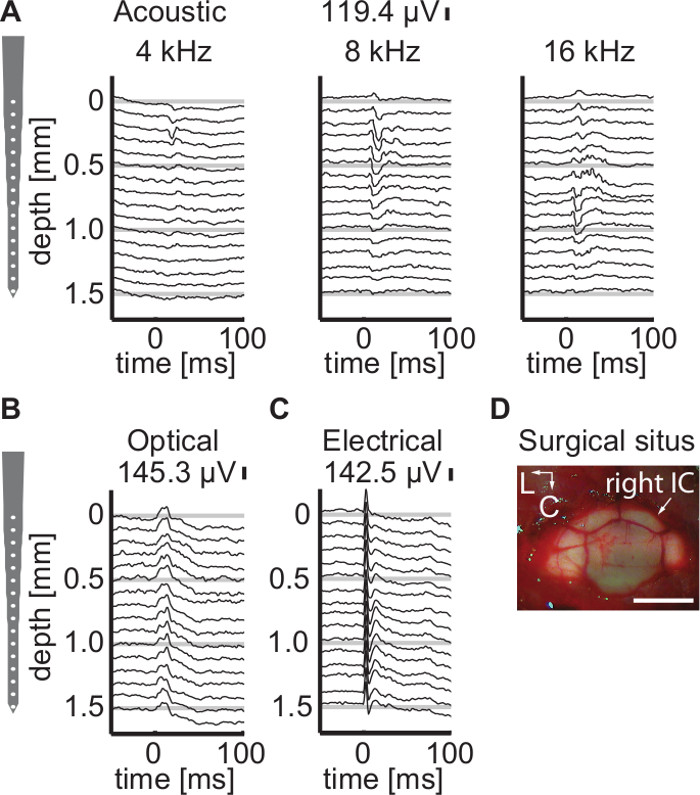
Figure 2: Single trial local field potentials recorded from the IC after acoustic, optogenetic, and electrical cochlear stimulation. A schematic representation of the multichannel recording probe is drawn on the left side of each panel. (A) Single presentations of pure tones with various frequencies (80 dB SPL) lead to a distinguishable gradient of activity with lower frequencies leading to more superficial activation than higher frequencies. This observation is used to evaluate successful insertion of the multichannel recording array into the IC. (B) Optical stimulation (24 mW, 6 msec duration) and (C) Electrical stimulation (biphasic, 80 μsec phase duration, 20 μsec interphase duration, 500 μA) also led to local field potentials in the IC often exhibiting comparable response amplitudes. (D) Surgical situs including both IC as well as a portion of the cerebellum. Scale bar 5 mm. Please click here to view a larger version of this figure.
Discussion
The described experiments demonstrate the optogenetic stimulation of the SGNs, and can, in principle, also be used to stimulate inner and/or outer hair cells, provided the expression of opsins. These experiments require much patience and care. As mentioned before, the most critical steps are a good cochleostomy/round window insertion as well as an appropriate position and orientation of the light source.
There are limitations with optogenetic stimulation when using ChR2. In our case oABR amplitude increases with light intensity and pulse duration but decreases when rate of stimulation increases15. Response latency also depends upon light intensity, decreasing when the intensity of the stimulus increases. We propose that these responses are the result of the kinetics of the ChR2 and the number of channels expressed in the SGNs. In addition, ChR2 can cause extra spikes, failures and membrane potential summation at high spike rates27,40. There have been continuous efforts to overcome these limitations32,33. We have recently reported the use of virus-mediated expression of the ChR2-variant CatCh14 in SGNs, which reduced the light requirement and allowed spiking to higher rates of stimulation39. Most recently the Boyden laboratory reported on Chronos, a channelrhodopsin with faster kinetics and high light sensitivity41.
We propose that optogenetic stimulation of the auditory pathway can contribute to auditory research and, in the future, to cochlear prosthetics. We consider that it can improve speech recognition and perception of prosody and music. Translation of this approach into the clinic requires many critical developments such as the optimization of channelrhodopsins, the development of multichannel optical stimulation technology for an optical cochlear implant as well as the demonstration of biosafety for optical stimulation and long-term genetic manipulation of SGNs.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was supported by the German Federal Ministry of Education and Research (Bernstein Focus for Neurotechnology grant 01GQ0810, to T. Moser, and MED-EL Germany); the German Research Foundation through the Center for Nanoscale Microscopy and Molecular Physiology of the Brain (FZT 103, T. Moser) and through the SFB889, to N. Strenzke and T. Moser).
Materials
| Urethane | Sigma Aldrich | U2500-100G | Anesthetic |
| Xylazine HCl | RXV | Sedative and analgesic | |
| Buprenorphine | Reckitt Benckiser | Analgesic | |
| Dumont #5 Forceps | Fine Science Tools | 11251-10 | It is used to hold hard tissue, e.g. bone or materials. Never use them to hold soft delicated tissue |
| Dumont #5 – Fine Forceps | Fine Science Tools | 11254-20 | Only to be used to hold soft tissue |
| Fine Scissors – Sharp | Fine Science Tools | 14060-09 | To open the skin and help with the muscle dissection |
| Lempert Rongeurs | Fine Science Tools | 16004-16 | They are very useful to easily remove the bone from the bulla |
| 473-nm laser | Changchun New Industries | MLL-III473 | 100 mW solid state 473 nm laser |
| Laser driver | Changchun New Industries | DPSSL MLL 100 mW | TTL operated laser driver |
| 250 µm optical fiber | Any comercial ; e.g. Thorlabs | M42L05 | |
| Acousto-optical modulator | Crystal Technology, Inc. | PCAOM VIS | Control the amount of light coupled into the fiber from the laser |
| Controller for Acousto-optical modulator | Crystal Technology, Inc. | 160T1-8SAR-24-0.8 | Control the acousto-optic modulator |
| Solo2 laser power & energy meter | Gentec-EO | Used to measure light intensity of the LED and the fiber coupled laser | |
| Blue µLED | Cree | C470UT200 | It is necessary to build several μLED devices because easily get damaged or the isolation is not good enough |
| TDT System | Tucker-Davis Technologies | RZ6-A-P1 | It can be used any system for stimulus generation presentation and data acquisition |
| Single-shank, 16-channel silicon probe | Neuronexus | a1x16-5mm-100-177-CM16LP | These are fragile devises, must be handled carefully and cleaned after use |
| Omnidrill | World Precision Instruments | 503598 | Perform craniotomy for IC recordings and reference screw implantation |
| Micro Drill Steel Burrs | any commercial; e.g. Fine Science Tools | 19007-07 | |
| Self tapping bone screw | any commercial; e.g. Fine Science Tools | 19010-10 | Reference screw |
| Micromanipulator | any commercial; e.g. Luigs+NeumannInVivo Unit Junior 4 axis | Positioning of recording probe |
References
- Rubinstein, J. T. Paediatric cochlear implantation: prosthetic hearing and language development. Lancet. 360 (9331), 483-485 (2002).
- Middlebrooks, J. C., Bierer, J. A., Snyder, R. L. Cochlear implants: the view from the brain. Current opinion in neurobiology. 15 (4), 488-493 (2005).
- Clark, G. M. The multiple-channel cochlear implant: the interface between sound and the central nervous system for hearing, speech, and language in deaf people-a personal perspective. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 361 (1469), 791-810 (2006).
- Zeng, F. G., Rebscher, S., Harrison, W., Sun, X., Feng, H. Cochlear implants: system design, integration, and evaluation. IEEE reviews in biomedical engineering. 1, 115-142 (2008).
- Wilson, B. S., Dorman, M. F. Cochlear implants: a remarkable past and a brilliant future. Hearing research. 242 (1-2), 3-21 (2008).
- Moore, D. R., Shannon, R. V. Beyond cochlear implants: awakening the deafened brain. Nature neuroscience. 12 (6), 686-691 (2009).
- Shannon, R. V. Multichannel electrical stimulation of the auditory nerve in man. II. Channel interaction. Hearing research. 12 (1), 1-16 (1983).
- Fishman, K. E., Shannon, R. V., Slattery, W. H. Speech recognition as a function of the number of electrodes used in the SPEAK cochlear implant speech processor. Journal of speech, language, and hearing research: JSLHR. 40 (5), 1201-1215 (1997).
- Kral, A., Hartmann, R., Mortazavi, D., Klinke, R. Spatial resolution of cochlear implants: the electrical field and excitation of auditory afferents. Hearing research. 121 (1-2), 11-28 (1998).
- Izzo, A. D., Suh, E., Pathria, J., Walsh, J. T., Whitlon, D. S., Richter, C. P. Selectivity of neural stimulation in the auditory system: a comparison of optic and electric stimuli. Journal of biomedical. 12 (2), 021008 (2007).
- Richter, C. P., Rajguru, S. M., et al. Spread of cochlear excitation during stimulation with pulsed infrared radiation: inferior colliculus measurements. Journal of neural engineering. 8 (5), 056006 (2011).
- Teudt, I. U., Maier, H., Richter, C. P., Kral, A. Acoustic events and “optophonic” cochlear responses induced by pulsed near-infrared laser. IEEE transactions on bio-medical engineering. 58 (6), 1648-1655 (2011).
- Wang, H., et al. High-speed mapping of synaptic connectivity using photostimulation in Channelrhodopsin-2 transgenic mice. Proceedings of the National Academy of Sciences of the United States of America. 104 (19), 8143-8148 (2007).
- Kleinlogel, S., Feldbauer, K., et al. Ultra light-sensitive and fast neuronal activation with the Ca2+-permeable channelrhodopsin CatCh. Nature neuroscience. 14 (4), 513-518 (2011).
- Friesen, L. M., Shannon, R. V., Baskent, D., Wang, X. Speech recognition in noise as a function of the number of spectral channels: comparison of acoustic hearing and cochlear implants. The Journal of the Acoustical Society of America. 110 (2), 1150-1163 (2001).
- Donaldson, G. S., Kreft, H. A., Litvak, L. Place-pitch discrimination of single- versus dual-electrode stimuli by cochlear implant users (L). The Journal of the Acoustical Society of America. 118 (2), 623-626 (2005).
- Srinivasan, A. G., Shannon, R. V., Landsberger, D. M. Improving virtual channel discrimination in a multi-channel context. Hearing research. 286 (1-2), 19-29 (2012).
- Zeng, F. G., et al. Speech dynamic range and its effect on cochlear implant performance. The Journal of the Acoustical Society of America. 111 (1 Pt 1), 377-386 (2002).
- Matic, A. I., Walsh, J. T., Richter, C. P. Spatial extent of cochlear infrared neural stimulation determined by tone-on-light masking. Journal of biomedical. 16 (11), 118002 (2011).
- Verma, R., Guex, A. A., et al. Auditory responses to electric and infrared neural stimulation of the rat cochlear nucleus. Hearing research. 310, 69-75 (2014).
- Fenno, L., Yizhar, O., Deisseroth, K. The development and application of optogenetics. Annual review of neuroscience. 34, 389-412 (2011).
- Hegemann, P., Nagel, G. From channelrhodopsins to optogenetics. EMBO molecular medicine. 5 (2), 173-176 (2013).
- Packer, A. M., Roska, B., Häusser, M. Targeting neurons and photons for optogenetics. Nature neuroscience. 16 (7), 805-815 (2013).
- Nagel, G., et al. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proceedings of the National Academy of Sciences of the United States of America. 100 (24), 13940-13945 (2003).
- Nagel, G., Szellas, T., Kateriya, S., Adeishvili, N., Hegemann, P., Bamberg, E. Channelrhodopsins: directly light-gated cation channels. Biochemical Society transactions. 33 (Pt 4), 863-866 (2005).
- Nagel, G., Brauner, M., Liewald, J. F., Adeishvili, N., Bamberg, E., Gottschalk, A. Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Current biology: CB. 15 (24), 2279-2284 (2005).
- Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G., Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nature neuroscience. 8 (9), 1263-1268 (2005).
- Bamann, C., Kirsch, T., Nagel, G., Bamberg, E. Spectral characteristics of the photocycle of channelrhodopsin-2 and its implication for channel function. Journal of molecular biology. 375 (3), 686-694 (2008).
- Ritter, E., Stehfest, K., Berndt, A., Hegemann, P., Bartl, F. J. Monitoring light-induced structural changes of Channelrhodopsin-2 by UV-visible and Fourier transform infrared spectroscopy. The Journal of biological chemistry. 283 (50), 35033-35041 (2008).
- Berndt, A., Prigge, M., Gradmann, D., Hegemann, P. Two open states with progressive proton selectivities in the branched channelrhodopsin-2 photocycle. Biophysical journal. 98 (5), 753-761 (2010).
- Kato, H. E., et al. Crystal structure of the channelrhodopsin light-gated cation channel. Nature. 482 (7385), 369-374 (2012).
- Hegemann, P., Möglich, A. Channelrhodopsin engineering and exploration of new optogenetic tools. Nature methods. 8 (1), 39-42 (2011).
- Mattis, J., Tye, K. M., et al. Principles for applying optogenetic tools derived from direct comparative analysis of microbial opsins. Nature methods. 9 (2), 159-172 (2012).
- Arenkiel, B. R., Peca, J., et al. In Vivo Light-Induced Activation of Neural Circuitry in Transgenic Mice Expressing Channelrhodopsin-2. Neuron. 54 (2), 205-218 (2007).
- Tomita, H., Sugano, E., et al. Visual Properties of Transgenic Rats Harboring the Channelrhodopsin-2 Gene Regulated by the Thy-1.2 Promoter. PLoS ONE. 4 (11), e7679 (2009).
- Hernandez, V. H., et al. Optogenetic stimulation of the auditory pathway. The Journal of clinical investigation. 124 (3), 1114-1129 (2014).
- Madisen, L., Mao, T., et al. A toolbox of Cre-dependent optogenetic transgenic mice for light-induced activation and silencing. Nature Neuroscience. 15 (5), 793-802 (2012).
- Yizhar, O., Fenno, L. E., Davidson, T. J., Mogri, M., Deisseroth, K. Optogenetics in neural systems. Neuron. 71 (1), 9-34 (2011).
- Hernandez, V. H., et al. Optogenetic stimulation of the auditory pathway. The Journal of clinical investigation. 124 (3), 1114-1129 (2014).
- Gunaydin, L. A., Yizhar, O., Berndt, A., Sohal, V. S., Deisseroth, K., Hegemann, P. Ultrafast optogenetic control. Nature neuroscience. 13 (3), 387-392 (2010).
- Klapoetke, N. C., Murata, Y., et al. Independent optical excitation of distinct neural populations. Nature methods. 11 (3), 338-346 (2014).
