

Summary
Denne protokol beskriver de skridt, omkostninger og nødvendige udstyr til at skabe E. coli-baseret celle ekstrakter og gennemføre in vitro- protein syntese reaktioner inden for 4 dage eller derunder. For at udnytte de fleksible karakter af denne platform for brede programmer, diskuterer vi reaktionsbetingelser, der kan tilpasses og optimeret.
Abstract
I de sidste 50 år fremstod celle-fri Protein syntese (CFPS) som en kraftfuld teknologi til at udnytte transcriptional og translationel kapaciteten af celler i et reagensglas. Længere behov at bevare levedygtigheden af cellen, og ved at fjerne den cellulære barriere, har CFPS været grundlagsforskning til nye applikationer i biomanufacturing af traditionelt udfordrende proteiner, samt programmer i rapid prototyping for metaboliske engineering, og funktionel genomforskning. Vores metoder til gennemførelse af en E. coli-baseret CFPS platform give nye brugere adgang til mange af disse programmer. Her, beskriver vi metoder til at forberede ekstrakt ved hjælp af beriget medier, forbløffet kolber og en reproducerbar metode til afstemmelige sonikering-baserede celle lysering. Denne ekstrakt kan derefter bruges til protein udtryk kan producere 900 µg/mL eller mere af super mappe grøn fluorescerende proteiner (sfGFP) i bare 5 h fra eksperimentel opsætning til dataanalyse, at passende reagens bestande har været forberedt på forhånd. Anslået start omkostninger at opnå reagenser er $4.500, som vil opretholde tusindvis af reaktioner på en anslået udgift af $0.021 pr. µg af protein produceret eller $0.019 pr. µL af reaktion. Derudover spejl protein udtryk metoder lethed af opsætningen reaktion ses i kommercielt tilgængelige systemer på grund af optimering af reagens forblandinger, på en brøkdel af prisen. For at muliggøre bruger hen til leverage den fleksible karakter af fælles fiskeripolitiks platform for brede programmer, har vi identificeret en række aspekter af den platform, der kan være tunet og optimerede afhængigt af de disponible ressourcer og protein udtryk resultater ønskede.
Introduction
Celle-fri Protein syntese (CFPS) fremstod som en teknologi, der er låst op en række nye muligheder for protein produktion, funktionel genomforskning, metabolisk engineering og mere inden for de sidste 50 år1,2. I forhold til standard i vivo protein udtryk platforme, CFPS giver tre vigtige fordele: 1) den celle-fri karakter af platformen muliggør fremstilling af proteiner, der ville være potentielt giftige eller udenlandske med celle3,4 ,5,6; 2) inaktivering af genomisk DNA og indførelsen af en skabelon DNA kodning molekylærgenetisk interesse kanal al den systemiske energi inden for produktionen af de(n) reaktion af interesse; og 3) den åbne karakter af platformen gør det muligt for brugeren at ændre og overvåge de reaktionsbetingelser og sammensætning i realtid7,8. Denne direkte adgang til reaktionen understøtter forstærkning af biologiske systemer med udvidet kemi og redox forhold for produktion af nye proteiner og tuning af metaboliske processer2,9, 10. direkte adgang også tillader bruger hen til kombinere fælles fiskeripolitiks reaktion med aktivitet assays i en single-puljen system for hurtig design-build-testen arbejdscykler,11. Kapacitet til at udføre fælles fiskeripolitiks reaktion i lille volumen dråber eller på papir-baserede enheder yderligere understøtter høj overførselshastighed discovery indsats og rapid prototyping12,13,14,15 ,16. Disse fordele og plug and play arten af systemet, CFPS entydigt har aktiveret en række bioteknologiske applikationer såsom fremstilling af proteiner, der er vanskeligt at solubly udtrykkelig i vivo17, 18,19,20, påvisning af sygdommen21,22,23, på efterspørgslen biomanufacturing18,24 ,25,26,27, og uddannelse28,29, som alle udvise fleksibilitet og nytte af den celle-fri platform.
CFPS systemer kan genereres fra en bred vifte af rå lysates fra begge prokaryote og eukaryote cellelinier. Dette giver mulighed for forskellige muligheder i systemet af valg, hver især har fordele og ulemper afhængigt af anvendelsen af interesse. CFPS systemer også varierer meget i forberedelsestid, omkostninger og produktivitet. Mest udnyttet almindeligt celle ekstrakter er produceret fra hvedekim, kanin reticulocyte, insekt celler og Escherichia coli celler, hvor sidstnævnte er den mest omkostningseffektive til dato, mens der producerer de højeste volumetriske udbytter af protein30 . Mens andre CFPS systemer kan være en fordel for deres medfødte posttranslationel modifikation maskiner, nye applikationer ved hjælp af E. coli-baserede maskiner er i stand til at bygge bro over kløften ved at generere site-specifically fosforyleres og glykosyleret proteiner på efterspørgslen31,32,33,34,35.
CFPS reaktioner kan køres i enten batch, kontinuerlig udveksling celle-fri (CECF) eller kontinuerlig flow celle-fri (CFCF) formater. Batch-formatet er et lukket system, hvis reaktion levetid er begrænset på grund af faldende mængder af reaktanter og ophobning af hæmmende biprodukter af reaktionen. CECF og CFCF metoder øge levetiden for reaktionen, og resultere dermed i øget volumetriske protein udbytter i forhold til batch reaktion. Dette opnås ved at tillade biprodukter af proteinsyntese skal fjernes fra reaktion fartøj, mens nye reaktanter leveres i løbet af reaktion2. For CFCF, kan protein af interesse også blive fjernet fra reaktionskammeret, mens i CECF, protein af interesse er fortsat i reaktionskammeret består af en semipermeabel membran36,37. Disse metoder er særligt værdifuldt i at overvinde fattige volumetriske udbytter af vanskelige at express proteiner af interesse38,39,40,41,42, 43. Udfordringer i forbindelse med gennemførelsen af CECF og CFCF tilgange er, at 1) mens de resulterer i mere effektiv brug af bio maskiner ansvarlig for transskription og translation, de kræver især større mængder reagenser, der øger samlede omkostninger og 2) de kræver mere kompleks reaktion opsætninger og specialiseret udstyr i forhold til batch format44. For at sikre tilgængelighed for nye brugere, beskrevet protokollerne heri fokus på batch format på reaktion mængder af 15 µL med specifikke henstillinger for at øge reaktion volumen til milliliter skala.
De metoder, der præsenteres heri aktiverer ikke-eksperter med grundlæggende laboratorium færdigheder (f.eks. studerende) at gennemføre cellevækst, udtrække forberedelse og batch format reaktion setup for en E. coli-baseret CFPS system. Denne tilgang er omkostningseffektive sammenlignet med kommercielt tilgængelige kits uden at ofre lethed af kit-baserede reaktion setup. Desuden, denne tilgang giver programmer i laboratoriet og i marken. Når det besluttes at implementere CFPS, bør nye brugere grundigt evaluere effektiviteten af konventionelle protein udtryk systemer opstart investeringer, som CFPS ikke kan være overlegne i hvert enkelt tilfælde. CFPS metoder beskrevet her muliggøre bruger hen til direkte gennemføre en lang række applikationer, herunder funktionel genomforskning, høj overførselshastighed afprøvning, produktion af proteiner, der er vanskelige for i vivo udtryk, som feltet applikationer, herunder biosensorer og undervisnings kits for syntetisk biologi. Yderligere programmer såsom metabolisk engineering, tuning af protein udtryk betingelser, sygdom og registrering skala-up ved hjælp af CECF eller CFCF metoder er stadig muligt, men kan kræve erfaringer med fælles fiskeripolitiks platform for yderligere ændring af reaktion betingelser. Vores metoder kombinere vækst i beriget medier og forbløffet kolber, med relativt hurtige og reproducerbare metoder til celle lysis gennem sonikering, efterfulgt af en forenklet CFPS reaktion setup, der udnytter optimeret forblandinger45. Mens de cellulære vækst metoder er blevet noget standardiseret inden for dette felt, varierer metoder til celle lysis meget. Ud over sonikering omfatter fælles lysis metoder udnyttelse af en fransk presse, en homogeniseringsapparat, perle piskeris, eller lysozym og andre biokemiske og fysiske forstyrrelser metoder46,47,48, 49. ved hjælp af vores metoder, ca. 2 mL rå celle ekstrakt er opnået per 1 L af celler. Denne mængde af celle ekstrakt kan støtte fire hundrede 15 µL CFPS reaktioner, hver producerende ~ 900 µg/mL reporter sfGFP protein fra skabelon plasmidet pJL1-sfGFP. Denne metode koster $0.021/µg af sfGFP produceret ($.019/µL af reaktion), bortset fra omkostningerne til arbejdskraft og udstyr (supplerende figur 1). Start fra bunden, denne metode kan gennemføres i 4 dage af en enkelt person og Gentag CFPS reaktioner kan være afsluttet inden for timer (figur 1). Derudover kan protokollen skaleres i volumen for større partier af reagens til brugerens behov. Vigtigere, kan protokollen præsenteres her implementeres af ikke laboratorium uddannet-eksperter som studerende, da det kun kræver grundlæggende laboratorium færdigheder. De procedurer, der er beskrevet nedenfor, og den tilhørende video er blevet specifikt udviklet for at forbedre tilgængeligheden af E. coli CFPS platform for bred anvendelse.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. medier forberedelse og cellevækst
-
Dag 1
- Streak E. coli BL21*(DE3) celler fra en glycerol lager på en LB-agar plade og Inkuber i mindst 18 timer ved 37 ° C.
- Forberede 50 mL LB medier og autoklave løsningen på en flydende cyklus for 30 min ved 121 ° C. Opbevares ved stuetemperatur.
-
Dag 2
- Forberede 750 mL 2 x YTP medier og 250 mL 0,4 M D-Glucose opløsning, som beskrevet i de supplerende oplysninger.
- Hæld 2 x YTP medierne i en autoklaveres 2,5 L forbløffet kolbe og D-Glucose løsning i en autoklaveres 500 mL glasflaske. Autoklave begge løsninger på en flydende cyklus for 30 min ved 121 ° C.
- Sikre, at begge sterile løsninger gemmes ved 37 ° C, hvis cellevækst udføres på den næste dag, at maksimere vækstrater ved podning. Ikke kombinere løsninger indtil podning.
Bemærk: Løsninger kan opbevares ved 4 ° C i 1-2 d hvis det er nødvendigt, selvom de 2 x YTP media er stærkt udsat for forurening. - Starte en overnight kultur af BL21(DE3) ved at vaccinere 50 mL af LB medier med en enkelt koloni af BL21(DE3) ved hjælp af en steriliseret loop og steril teknik til at undgå forurening.
- Placere 50 mL af BL21*(DE3) LB kultur i et 37 ° C 250 rpm ryster inkubator og vokse natten til 15-18 h.
- Forberede og sterilisere alle materialeudgiften til dage 3 og 4, herunder: to 1 L centrifuge flasker, 4 x kold 50 mL konisk rør (vejer og optage masserne af tre) og mange 1,5 mL mikrofuge rør.
-
Dag 3
- Fjerne den 50 mL overnight kultur af BL21*(DE3) i LB fra ryster rugemaskine og måle OD600 på et spektrofotometer ved hjælp af en 1:10 fortynding med LB medier. Beregning af volumen af overnight kultur nødvendigt at føje til 1 L af medier til en start OD600 af 0,1 (For eksempel, hvis en OD600 af en 1:10 fortynding læses som 0,4, podes 25 mL af ufortyndet OD600 = 4.0 overnight kultur i 1 L, 2 x YTP G).
- Fjern de varmede 2 x YTP medier og D-Glucose løsninger fra 37 ° C væksthuset sammen med 50 mL af LB kultur. Brug steril teknik, omhyggeligt hæld D-Glucose løsning i de 2 x YTP media (undgå siderne af forbløffet kolben).
Bemærk: Tilsætning af D-Glucose fuldender opskrift på 1 L, 2 x YTPG. - Opretholde steril teknik, podes på 1 L 2 x YTPG løsning med den passende mængde 50 mL kultur for at begynde 1 L kulturen på en 0,1 OD600. Umiddelbart Placer den podede 1 L kultur i et 37 ° C ryster inkubator ved 200 rpm.
- Tage de første OD600 læsning efter den første time af vækst (mellemliggende fase typisk tager 1 time). Ikke fortyndes kulturen. Fortsætte med at tage OD600 målinger ca. hver 20-30 min. indtil OD600 når 0,6.
- Ved ankomsten til OD600 = 0,6, tilsættes 1 mL 1 M IPTG (endelig koncentration i 1 L kultur = 1 mM) til 2 x YTPG kultur.
Bemærk: Ideel induktion OD600 er 0,6; men en række 0,6-0,8 er acceptabel. Induktion af IPTG er for endogene produktion af T7 RNA Polymerase (T7RNAP). - Efter induktion, måle OD600 ca. hver 20-30 min. indtil den når 3.0.
Bemærk: Køle ned centrifuge til 4 ° C i løbet af denne tid. Forberede kolde S30 buffer som beskrevet i de supplerende oplysninger. Hvis S30 buffer er forberedt på forhånd, sikre at DTT ikke føjes indtil dag i brug. - Når OD600 når 3.0 (figur 2A), hæld kulturen i et koldt 1 L centrifuge flaske i en is-vandbad. Forberede en vandfyldte 1 L centrifuge flaske lige vægt anvendes som en balance i centrifugen.
Bemærk: Absorbans værdier varierer fra instrument til instrument. Mens OD600 af høsten af BL21(DE3) ikke er en følsom variabel, anbefales det, at brugeren evaluerer og optimerer denne variabel som en fejlfinding foranstaltning. Større spektrofotometre kan resultere i relativt lavere OD600 aflæsninger i forhold til mindre kuvette-baserede spektrofotometre. - Der centrifugeres i 1 L flasker i 10 min på 5.000 x g og 10 ° C at sammenpresse cellerne.
- Langsomt hæld supernatanten og bortskaffe det efter institutionens biologisk affald procedurer. Sted pellet på is.
- Med en steril spatel, skrabe celle toerstoffet fra centrifuge flasken og overføre det til en kold 50 mL konisk slange.
- Der tilsættes 30 mL koldt S30 buffer til det koniske rør og resuspenderes celle af vortexing med korte byger (20-30 s) og hvileperioder (1 min) på is indtil fuldt genopslemmes med nogen lunser.
- Når pelleten er fuldt genopslemmes, skal du bruge en anden 50 mL konisk tube med vand som en balance og centrifugeres i 10 min på 5000 x g og 10 ° C (pre afkølet til 4 ° C).
Bemærk: Dette afslutter den 1st af 3 vasker kræves, når høsten cellerne. - Hæld supernatanten og bortskaffe det efter institutionens biologisk affald procedurer. Resuspenderes med 20-25 mL koldt S30 buffer og centrifugeres i 10 min på 5000 x g og 10 ° C (pre afkølet til 4 ° C).
Bemærk: Dette afslutter den 2nd af 3 vasker. - Igen, hæld supernatanten og bortskaffe det efter institutionens biologisk affald procedurer. Præcis 30 mL S30 buffer og vortex igen til resuspenderes tilsættes.
- Ved hjælp af de 3 pre vejes, kold 50 mL konisk reagensglas og en serologisk pipette fyldstof med en steril pipette, alikvot 10 mL resuspenderet pellet/S30 buffer blandingen i hver af de 3 koniske rør.
Bemærk: Opdele celler i 3 rør er ikke påkrævet, men dette trin resulterer i mindre celle pellets (~ 1 g) for øget bekvemmelighed på senere trin. - Centrifugeres alle rør, ved hjælp af passende saldi efter behov, for 10 min på 5000 x g og 10 ° C (pre afkølet til 4 ° C).
Bemærk: Dette fuldender det endelige wash skridt. - Hæld supernatanten og bortskaffe det efter institutionens biologisk affald procedurer. Fjern den overskydende S30 buffer ved omhyggeligt aftørring indersiden af koniske rør og hætte med en ren væv; undgå at berøre pelleten.
- Tilbagevejes rør på en Analysevægt og optage den endelige pellet vægt på hver tube.
Bemærk: Protokollen kan pause på dette tidspunkt. Pellets kan være flash frosset i flydende nitrogen og opbevares ved-80 ° C i op til et år, indtil det skal bruges til ekstrakt forberedelse.
2. rå celle ekstrakt forberedelse - dag 4
- Til ekstrakt forberedelse, holde celler kold på is under hvert trin. Der tilsættes 1 mL af kolde S30 buffer pr. 1 g af cellemasse af toerstoffet. Sikre at dithiothreitol (DTT) er blevet suppleret til S30 buffer til en slutkoncentration på 2 mM.
Bemærk: Køle ned microcentrifuge til 4 ° C i løbet af denne tid. - Resuspenderes celle af vortexing med korte byger (20-30 s) og hvileperioder (1 min) på is indtil fuldt genopslemmes. Hvis resuspension er vanskeligt, forlade pellets i isbad i 30 min til optøning.
- Overføre 1,4 mL resuspenderet celler ind i en 1,5 mL mikrofuge rør.
- Placere en 1,5 mL rør indeholdende 1,4 mL resuspenderet celler i et bad med is vand i et bægerglas. Rens med ultralyd i 45 s på efterfulgt af 59 s ud til 3 samlede cykler, med amplitude fastsat til 50%. Luk og vend rør forsigtigt blandes de off perioder. I alt, skal du levere 800-900 Jørgensen af energi til hver 1,5 mL mikrofuge rør indeholdende 1,4 mL resuspenderet celler (figur 3A & 3B).
Bemærk: Dette trin er følsomme over for sonikator type og model anvendes og skal optimeres, hvis udstyret er anderledes, end der er angivet for denne procedure. To komplementære metoder kan bruges til opskalering mængden af ekstrakt udarbejdet under dette trin: 1) flere 1,5 mL mikrofuge rør kan være sonicated parallelle, og/eller 2) større mængder kan være sonicated i koniske rør (op til 15 mL celle resuspension pr tube) , skalering mængden af energi der leveres som tidligere beskrevet 29,45. - Umiddelbart efter ultralydbehandling er komplet, tilføje 4,5 µL 1 m DTT (supplere en yderligere 2 mM DTT) i 1,4 mL lysate og invertere flere gange for at blande. Røret anbringes på is. Gentag trin 2.4 og 2.5 for enhver yderligere rør af resuspenderede celler, før du fortsætter til centrifugering.
- Microcentrifuge prøver på 18.000 x g og 4 ° C i 10 min (figur 3 c).
- Der afpipetteres supernatanten i en ny 1,5 mL mikrofuge tube. Lave ikke forstyrre pelleten; Det er bedre at efterlade nogle supernatanten for at opretholde renhed end for at forstyrre pelleten i bestræbelser på at maksimere afkastet.
- Inkuber supernatanten fra det forrige trin på 250 rpm og 37 ° C i 60 minutter af tape rør til at den rystende platform af inkubator (dette er afstrømning reaktion).
- Microcentrifuge prøver på 10.000 x g og 4 ° C i 10 min.
- Fjern supernatanten uden at forstyrre pelleten og overføre det til en ny tube. Oprette mange 100 µL delprøver af ekstrakt til opbevaring.
Bemærk: Protokollen kan blive standset her, og ekstraktet kan være flash frosset i flydende nitrogen og opbevares ved-80 ° C i op til et år, indtil det skal bruges til CFPS reaktioner. Mindst 5 fryse-tø cykler kan være gennemgået uden bekostning at udtrække produktivitet (figur 4).
3. celle-gratis Protein syntese Batch Format reaktioner
- Tø løsninger A og B, DNA skabelon, BL21(DE3) ekstrakt (hvis frosne), T7RNAP og en alikvot af vand af Molekylær renhed.
Bemærk: CFPS reaktion skabelon kan findes i de Supplerende oplysninger. Løsninger A og B opskrifter er fastsat i de Supplerende oplysninger og svarer til bestemte koncentrationer for talrige reagenser til at støtte PANOx-SP baseret energisystem til CFPS. Hver reagens og acceptabel variation i disse reagens koncentrationer, der kan understøtte CFPS rolle har været fast besluttet på50. En T7RNAP rensning protokol kan findes i de supplerende oplysninger51. Supplerende T7RNAP kan øge volumetriske udbyttet, men er ikke nødvendigt, hvis T7RNAP er induceret under cellevækst. Plasmid DNA skabelon (pJL1-sfGFP) kan forberedes ved hjælp af en maxiprep kit med to vasker ved wash buffer til sættet, efterfulgt af en efterbehandling DNA-oprydning ved hjælp af en PCR rensning kit (figur 2B). Lineære DNA skabeloner kan også bruges i CFPS reaktioner. - Mærke den nødvendige mængde af mikrofuge rør behov for CFPS reaktioner.
Bemærk: Reaktioner kan udføres i forskellige størrelser, fartøj, men et mindre fartøj kan mindske volumetriske protein udbytter (figur 2 c). Optrapning af en reaktion i samme størrelse fartøjet kan også reducere volumetriske udbytter, som en funktion af faldende ilt udveksling, på grund af et fald i areal til volumen-forholdet. Når stigende reaktion volumen over 100 μL, anbefales det at bruge flad bund godt plader 31,37,52. - Tilføje 2,2 µL af opløsning A, 2.1 µL af løsning B, 5 µL af BL21*(DE3) ekstrakt, 0,24 μg af T7RNAP (16 μg/mL endelige koncentration), 0,24 ng af DNA skabelon (16 ng/mL endelige koncentration), og vand for at bringe de endelige mængden til 15 µL.
Bemærk: Vortex løsninger A og B ofte under installationen af reaktion at undgå sedimentation af komponenter og sikre, at hver reaktion får en homogen delprøve af hver løsning. Undgå vortexing ekstrakt, i stedet invertere røret til mix. - Efter alle reagenser er blevet føjet til reaktionen, mix hver tube af pipettering op og ned eller forsigtigt vortexing samtidig med at den endelige reaktionsblandingen er kombineret i en enkelt 15 µL perle i bunden af 1,5 mL mikrofuge tube.
- Placer hver reaktion til 37 ° C inkubator uden ryster for 4 h, eller 30 ° C natten over.
Bemærk: Vellykket reaktioner kan kvalitativt vurderes visuelt baseret på den grønne farve af sfGFP produktet inden for fælles fiskeripolitiks reaktionsblandingen (figur 3D). Udtryk for protein af interesse kan også bekræftes af SDS-PAGE (supplerende figur 2).
4. kvantificering af Reporter Protein, [sfGFP]
- Indlæse 48 µL af 0,05 M HEPES, pH 8, i hver godt behov for kvantificering (normalt udføres i tre eksemplarer pr. reaktion tube).
- Fjerne reaktioner fra inkubator. Pipetteres op og ned for at blande hver reaktion, derefter overføre 2 µL af reaktion i de 48 µL af 0,05 M HEPES, pH 8. Pipette op og ned igen i godt at blande.
- Når alle reaktioner er indlæst og blandet, 96 godt pladen anbringes i fluorometer og måle sfGFP slutpunkt fluorescens.
Bemærk: Excitations- og bølgelængder for sfGFP fluorescens kvantificering er 485 nm og 510 nm, henholdsvis. - Brug en tidligere genererede standardkurve, bestemme [sfGFP] fra opnåede fluorescens aflæsninger.
Bemærk: For at generere en standardkurve for sfGFP koncentration versus fluorescens intensitet anvisninger i supplerende oplysninger (supplerende figur 3). Brugerne bliver nødt til at etablere en standardkurve for deres instrument, eftersom instrumentet følsomhed kan variere.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Vi har præsenteret en sonikering-baserede E. coli ekstrakt forberedelse protokol, der kan gennemføres over en fire-dages span, med figur 1 viser den proceduremæssige fordeling over hver dag. Der er formbarhed til de skridt, der kan udfyldes hver dag med forskellige midlertidigt stop punkter, men vi har fundet denne arbejdsproces til at være den mest effektive til at udføre. Derudover både celle pellets (trin 1.3.18) og fuldt forberedt ekstrakt (trin 2.10) er stabil ved-80 ° C i mindst et år, tillade bruger hen til skabe større lagre af hvert gemme til brug på et senere tidspunkt17. Ikke kun er uddraget stabil over lange perioder, men ekstrakt kan også gennemgå mindst fem fryse tø cykler uden betydelige tab af produktivitet (figur 4). Dette giver mulighed for større delprøver af ekstraktet opbevares i flere anvendelser, hvis fryseren opbevaringsplads er begrænset. Vi anbefaler imidlertid flere mindre delprøver (~ 100 μL) ekstrakt når det er muligt.
Med hver ny ekstrakt forberedelse anbefales det, at brugeren udfører en magnesium titrering for at bestemme den optimale mængde af magnesium for partiet ekstrakt. Brugere kan kvantificere batch til batch variationer i samlede proteinkoncentration celle ekstrakt af Bradford assay. For højere udfører ekstrakter, vi typisk ser total protein koncentrationer på 30-50 mg/mL, og inden for dette interval der er ingen indlysende sammenhæng mellem total protein koncentrationer og celle ekstrakt ydeevne. Vi anbefaler derfor, at brugerne tune magnesium koncentrationer i overensstemmelse hermed for at sikre, at protein- og nukleinsyre funktionalitet er maksimeret for hvert ekstrakt parti. Magnesium er vigtigt for korrekt DNA replikation, transskription og translation, men overdreven niveauer kan være til skade for disse processer53. For at vise denne afhængighed, vi har udført en co titrering af magnesium og udtrække volumen for at bestemme den optimale kombination, der minimerer mængden af ekstrakt nødvendige, samtidig med at en produktiv reaktion (figur 5). Fra dette eksperiment anbefaler vi at bruge 5 µL af ekstrakt og 10 mM Mg2 + for ekstrakt med en samlede proteinindhold på 30 mg/mL, for at opnå mere end 1.000 μg/mL af sfGFP.
Vores erfaring med CFPS har også tilladt os at finde trin inden for den protokol, der kan varieres uden skade for den samlede produktivitet i systemet, og andre, der er integreret til en højt ydende CFPS system. Især, de endelige OD600 af celle høst påvirker ikke væsentligt det endelige output af fælles fiskeripolitiks reaktion, og celler kan realistisk høstes overalt fra 2,7-4.0 OD600. Dette er den tidlige eksponentielle fase af vækst hvor ribosomet koncentration per celle er den højeste og den translationel maskiner er de mest aktive støtter hurtige vækst. Denne bemærkning giver brugerne fleksibilitet til at optimere deres egne procedurer. Vi anbefaler høst på omkring 3,0 OD600 for at fange cellerne på en OD600 tættere til 3.3 af tid høst er komplet (figur 2A). Variabler, der påvirker CFPS udbytter omfatter skabelon DNA kvalitet, reaktion fartoejsstoerrelse og de relative mængder af celle ekstrakt og magnesium ion i reaktionen. Vi har fundet DNA kvalitet har bemærkelsesværdige batch til batch variationer. For at løse dette, anbefales det, at brugere rense DNA via en midi eller maxi prep, efterfulgt af en yderligere DNA oprydning trin enten på DNA oprensning kolonne bruges i maxiprep, eller efter rensning ved hjælp af en ekstra DNA oprydning kit. Dette forbedrer reproducerbarhed i DNA kvalitet til CFPS reaktioner og resultater i mere robust protein produktion (figur 2B). Reaktion fartøj påvirker også volumetriske udbytter, således at protein produktion af identiske reaktion opsætninger i varierende fartøjer diskenhederne kan variere op til 40%. Det har været teoretiserede at boost i volumetriske høstudbyttet observeret i større fartøjer på grund af en øget areal af reaktionsblandingen, giver mulighed for bedre udveksling af ilt (figur 2 c), og andre har yderligere styrket volumetriske udbyttet ved at køre CFPS reaktioner i store fladbundede plader, som vi anbefaler for reaktioner over 100 µL17,31,37,52.

Figur 1 : Tidslinje for kultur vækst, produktion af celle ekstrakt, opsætning og kvantificering af CFPS reaktioner. Brugeren kan gennemføre CFPS platform for deres forskning applikationer gennem denne fire-dages arbejdsgang. Reagens repræsenterer den primære tid og omkostninger investering for den første runde af dette eksperiment og mindskes betydeligt efter reagenser bestande er etableret. Derudover kan celle pellets og forberedt celle ekstrakt opbevares i over et år på-80 ° C, så brugeren kan begynde tidslinje på forskellige trin for hurtigere resultater. Brugeren kan også pause på forskellige trin for at ændre tidslinje for denne arbejdsproces. Venligst klik her for at se en større version af dette tal.

Figur 2 : Kan ændres betingelserne for CFPS og effekter på volumetriske reaktionen udbytter. A. ekstrakt produktivitet sammenligning baseret på høst BL21(DE3) celler på forskellige OD600 aflæsninger. Baseret på dette plot, anbefaler vi høst på en OD600 af 3.3 at producere mindst 1000 µg/mL af target protein. Reaktioner blev udført på en 15 μl skala i 1,5 mL mikrofuge rør. B. sammenligningen af to DNA maxiprep vask protokoller med og uden efter rensning DNA-oprydninger. pJL1-sfGFP plasmider undergik en maxiprep med et eller to vasker efterfulgt af en efter rensning oprydning af PCR rensning kit. For at opnå ~ 900 µg/mL af protein udtryk, foreslår vi, at udføre en efter rensning DNA oprydning uanset antallet af maxiprep vasker. Reaktioner blev udført på en 15 μl skala i 1,5 mL mikrofuge rør. C. 15 µL CFPS reaktioner udført i forskellige fartøjer, der spænder fra 2 mL 0,6 mL mikrofuge rør. "Neg" repræsenterer en negativ kontrol hvor ingen DNA skabelonen blev tilføjet til reaktionen. Alle fejllinjer udgør 1 standardafvigelse af tre uafhængige reaktioner for hver betingelse, som hver var kvantificeret i tre eksemplarer. Venligst klik her for at se en større version af dette tal.

Figur 3 : Nøglen proceduremæssige opsætninger og resultater for at skabe produktive uddrag. A. korrekt opsætning af sonikering isvand bad for køling af prøven, mens varmeudvikling under sonikering. B. 1,5 mL mikrofuge rør indeholdende resuspenderet celle pellet før (venstre) og post (højre) sonikering. Den resulterende lysate skal vise en mørkere nuance i forhold til resuspenderet celle pellet. C. korrekt adskillelse af supernatanten og pellet celle lysate efter 18.000 x g centrifugering. D. CFPS reaktioner efter 4 h inkubation ved 37 ° C. 1,5 mL mikrofuge tube til højre (vellykket reaktion) viser synlige fluorescens af sfGFP reporter protein på ~ 900 μg/mL. Negativ kontrol røret på venstre, mangler template DNA og simulerer en mislykket reaktion, viser en klar løsning med ingen fluorescens. Venligst klik her for at se en større version af dette tal.

Figur 4 : Ændring i protein udtryk over 5 fryse-tø cykler til CFPS uddrag. Ekstrakt fremstillet af den samme vækst gennemgik fem fryse optøningscykler via flydende kvælstof flash frysning efterfulgt af optøning på køl. Ingen væsentlige ændringer i uddrag produktivitet for at udtrykke sfGFP blev set over de fem fryse-tø cykler. Reaktioner blev udført på en 15 μl skala i 1,5 mL mikrofuge rør. "Neg" repræsenterer en negativ kontrol hvor ingen DNA skabelonen blev tilføjet til reaktionen. Alle fejllinjer udgør 1 standardafvigelse af tre uafhængige reaktioner for hver betingelse, som hver var kvantificeret i tre eksemplarer. Venligst klik her for at se en større version af dette tal.

Figur 5 : CFPS for reaktioner med varierende [Mg2+] og udtrække diskenheder versus [sfGFP]. [Mg2 +] varierede fra 8 mM til 14 mM med 2 mM-trin og udtrække diskenheder varierede fra 3 µL til 7 µL med 1 µL intervaller. Farve kode repræsenterer [sfGFP] fremstillet af høj (rød) til lav (lilla). For at maksimere reagens effektivitet samtidig opretholde høje protein produktion, vi anbefaler at bruge 5 µL ekstrakt og 10 mM Mg2+ for ekstrakter, der har en samlet proteinindhold ~ 30 mg/ml, som bestemmes af Bradford assay. Oprindelige punkter til at generere contour plottet var baseret off slutpunkt fluorescens af tre uafhængige reaktioner for hver betingelse, som hver blev målt i tre eksemplarer. Reaktioner blev udført på en 15 μl skala i 1,5 mL mikrofuge rør. Venligst klik her for at se en større version af dette tal.
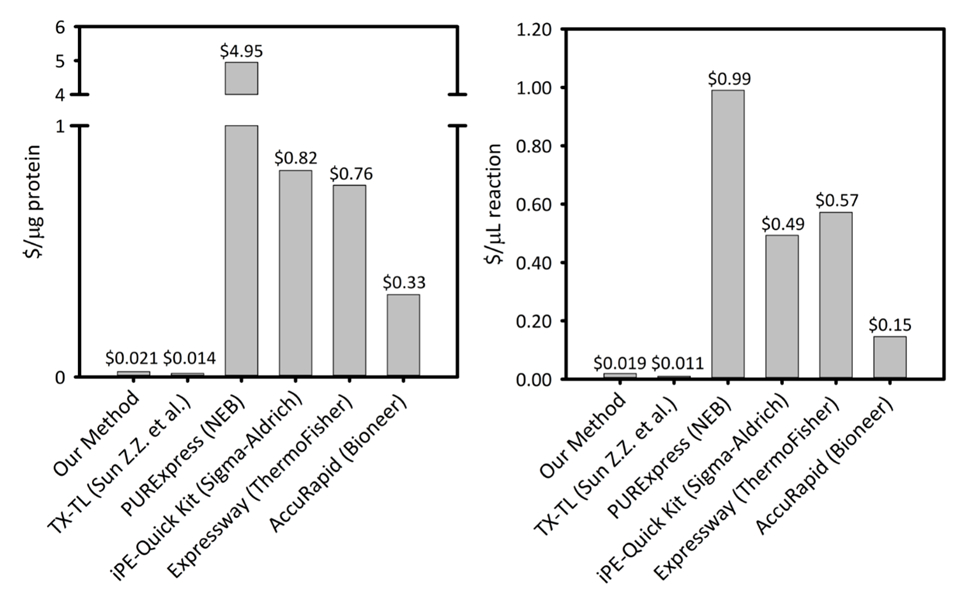
Supplerende figur 1: koster pr. mikrogram af protein produceret og microliter reaktion på tværs af seks celle-fri protein syntese platforme. Vores platform er sammenlignet med fem forskellige celle gratis protein syntese kits/platforme med varierende produktivitet og priser. Vores sonikering baseret CFPS platform er mere omkostningseffektive i begge $/ µg af protein og $/ µL af reaktion end de fleste kommercielle kits og giver lethed, hvormed et kit til reaktion setup, mens de resterende omkostninger sammenlignes med andre akademiske CFPS platforme. Venligst klik her for at se en større version af dette tal.
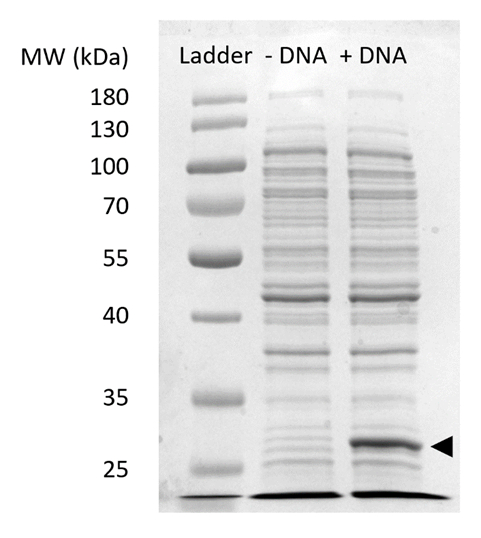
Supplerende figur 2: SDS-PAGE sfGFP udtryk i CFPS. Celle-gratis protein syntese reaktioner med (+ DNA) og uden (-DNA) DNA skabelon for sfGFP blev kørt på en 12% SDSPAGE gel til at demonstrere udtryk for sfGFP observeret på 27 kDa (sort pil). Traditionelle SDS-PAGE teknikker blev brugt. Hver prøve læsset på gelen inkluderet 18 µg af total protein baseret på Bradford assay kvantificering af total protein i cellen uddrag. Baseret på fluorescens intensitet målinger og vores standardkurven, vi anslår, at den "+ DNA" lane indeholder 0,42 µg for sfGFP. For at få disse prøver, køres CFPS reaktioner på en 15 µL skala i 1,5 mL mikrofuge rør producerer volumetriske udbytter i overensstemmelse med figur 3 c. Venligst klik her for at se en større version af dette tal.
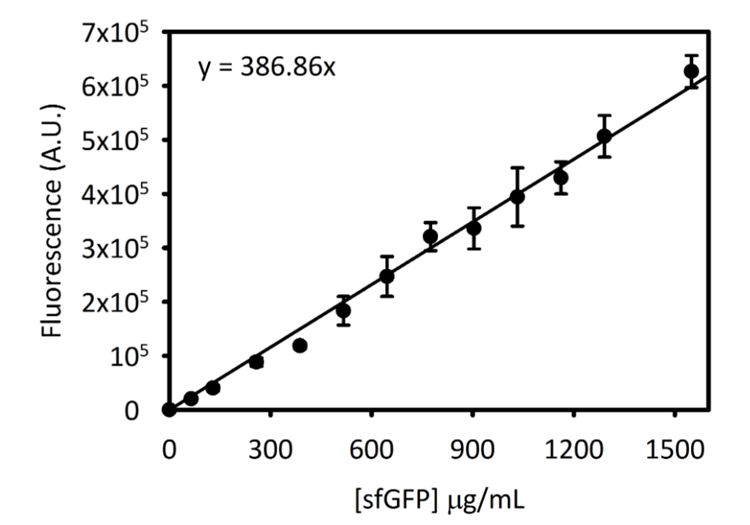
Supplerende figur 3: Standardkurven for sfGFP på Cytation 5. Denne kurve blev bestemt ved hjælp af de metoder, der er skitseret ovenfor. Alle data, der indsamles for dette manuskript blev konverteret fra slutpunkt fluorescens aflæsninger til [sfGFP] i µg/mL ved hjælp af denne standardkurven. Venligst klik her for at se en større version af dette tal.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Celle-fri proteinsyntesen fremstod som en kraftfuld og aktivering teknologi for en lang række applikationer lige fra biomanufacturing til rapid prototyping af biokemiske systemer. Bredden af programmer understøttes af kapacitet til at overvåge, manipulere, og øge cellulære maskiner i realtid. På trods af den ekspanderende effekt af denne platformteknologi, brede tilpasning er forblevet langsomt tekniske nuancer i gennemførelsen af metoderne. Gennem denne indsats tilstræber vi at give enkelhed og klarhed for at etablere denne teknologi i nye labs. Mod denne ende, vores protokol for en E. coli-baseret celle-fri protein syntese platform kan opnås inden for en opstart af fire dage ved laboratorium uddannet ikke-eksperter, som studerende (figur 1). Desuden, når en bestand af reagenser og ekstrakt er produceret, kan efterfølgende CFPS batch reaktioner oprette, rugede, og kvantificeres i bare 5 h. En enkelt, 1 L cellevækst kan resultere i nok ekstrakt for fire hundrede 15 µL CFPS reaktioner, mens enkelt batch præparater af de andre celle-fri reagenser kan understøtte tusindvis af reaktioner. Reagens præparater kan også skaleres, hvis en endnu større lager er nødvendig. CFPS reaktioner kan være setup i en høj overførselshastighed måde, ved hjælp af en 96-brønd plade eller PCR rør til afprøvning af en række betingelser i parallel. Volumetriske udbyttet vil falde når ved hjælp af mindre fartøjer som det ses i figur 2 c. CFPS reaktioner kan også skaleres fra microliters til snesevis af milliliter af samlede reaktion lydstyrke for at øge det samlede proteinindhold afkast for en enkelt betingelse. Når optrapning volumen, er de primære overvejelser at volumetriske reaktionen udbytter falde som det overflade område til volumen-forholdet på reaktionen aftager37,52. For at skala-up samtidig opretholde lignende volumetriske udbytter af protein udtryk, brugere skal opdele reaktion volumen i talrige reaktion fartøjer og/eller øge fartøjernes størrelse. For reaktionen skalaer spænder fra 15 µL - 100 µL i volumen, anbefales talrige 15 µL reaktioner i parallel. Reaktioner på over 100 µL i volumen, 24-godt-bundplade er anbefalet og 12-godt plader anbefales til reaktion mængder overstiger 600 µL. Sådanne bindinger af reaktion diskenheder og fartøjer giver sammenhæng i volumetriske reaktionen udbytter på skala-up17,31,37,52. Skalere op ud over disse mængder kan opnås ved at udnytte flere brønde af pladen i parallel. Brug dette format, kan reaktionen skaleres til over 10 mL samlede volumen. Optimere kombinationen reaktion volumen-reaktion fartøj kan støtte applikationer af biomanufacturing uden at ofre produktiviteten i reaktionen.
Når du udfører denne protokol, er der et par vigtige overvejelser, der påvirker volumetriske reaktionen udbytter samt indikatorer tilknyttet dårligt udfører ekstrakt. For at sikre ordentlig lysis og forhindre denaturering af funktionelle omskrivning/oversættelse maskiner, er det vigtigt at afbøde den varme, der produceres under lysering. Fordyb celle resuspension i en is vandbad under sonikering til hurtigt sprede hede under ultralydbehandling (figur 3A). En indikator for effektiv celle lysis er fremkomsten af en mørkere udseende af cellen lysate i forhold til pre-sonicated prøver (figur 3B). For brugeren fleksibilitet er sonikator og sonde vist i figur 3A tilpasningsdygtige til en vifte af diskenheder fra 100 µL til 15 mL resuspenderet celler. For at opnå dette, kan brugeren justere antallet af joule leveret til lysering af den ønskede mængde af celler. Derudover kan større mængder af ekstrakt være forberedt gennem to komplementære metoder. Brugere kan sonikeres flere rør i parallel og/eller der sonikeres større mængder af celle resuspension, skalering mængden af energi proportionalt med mængden som tidligere beskrevet29,45. Endnu et skridt, der angiver ekstrakt kvalitet er centrifugering skridt efter celle lysering. Indlæg celle lysis, vi anbefaler centrifugering ved 18.000 x g til at give en klar opdeling mellem supernatanten (omskrivning/oversættelse maskiner, fragmenterede genomisk DNA, som ikke længere funktioner på skabelon omskrivning/oversættelse) og pellet ( uønskede cellulære komponenter såsom cellemembranen og bundfaldne proteiner) (figur 3 c). Vi har fundet at centrifugering på 18.000 x g forbedrer adskillelse, hvilket resulterer i bedre reproducerbarhed i forhold til ture ved lavere hastigheder som 12.000 x g. For nemheds skyld anbefaler vi at bruge en tabel-top nedkølet centrifuge, i stand til at opnå et minimum af 12.000 x g. Dette trin er også almindeligvis udføres på 30.000 x g, som bør overvejes, hvis det relevante udstyr er tilgængelige54,55,56,57,58, 59 , 60. Uddrag ydeevne påvirkes ikke af centrifugering hastigheder på dette trin, da der opnås korrekt adskillelse. Når du fjerner den ønskede supernatanten, er det bedst at undgå overskyet materialer, der findes på grænsen mellem supernatanten og pellet, da denne forurening vil reducere produktiviteten af ekstraktet. Sigter for renhed af supernatanten resulterer i mere produktive ekstrakter og er værd den reducerede mængde ekstraktrumfanget fremstillet for nye brugere.
Det er vigtigt at bemærke, at mens de metoder, vi har præsenteret er reproducerbare og kan udføres af forskere med minimal ekspertise, der kan være batch til batch og reaktion til reaktion variation. Dette kan tilskrives variation af lysate efter ultralydbehandling61proteom sammensætning. Den batch til batch variation, at vi har observeret er generelt formindsket ved tilskud med T7RNAP og optimering af magnesium koncentrationer. Eksogene tilføjelsen af T7RNAP er udbredt blandt CFPS reaktioner til støtte for optimal protein udtryk, og vi finder, at have to kilder af T7RNAP - endogene udtryk i BL21*(DE3) og den supplerende T7RNAP til en endelig koncentration på 16 µg/mL - forbedrer batch til batch reproducerbarhed for nye brugere45,46. Med erfaring, kan brugere ændre deres eksperimenter for at bruge kun en enkelt kilde til T7RNAP, hvis det ønskes. Kvantificering af total proteinindholdet af et nyt parti ekstrakt og passende justering af Mg2 + koncentration kan også bidrage til at mindske batch til batch variationer i volumetriske protein udtryk udbytter. Variationer i protein udtryk kan også være på grund af forskelle i størrelse og struktur af protein af interesse, codon brugen af genet og dens tilsvarende ribosomet bindingssted af gen af interesse, samt typen udtryk vektor, der anvendes62 ,63. Af disse grunde, kan nogle proteiner ikke udtrykke samt model protein sfGFP, hvilket resulterer i reducerede volumetriske udbytter fra CFPS reaktioner.
Begrænsninger af den præsenterede CFPS teknik omfatter, det ikke kan være direkte egnet til alle anvendelser af celle-fri, såsom metabolisk teknik og tuning af udtryk betingelser, uden yderligere ændringer til protokollerne. Men vi mener, at denne protokol vil danne grundlag for etablering af fælles fiskeripolitiks platform i nye laboratorier og levere ikke-eksperter med evnen til at gennemføre indledende celle-fri reaktioner i deres labs. Efter indledende implementering, kan forskere eksperimentere med platformen til at gøre deres egne ændringer for mere specifikke applikationer baseret på anden litteratur på området.
Den fælles fiskeripolitiks platform omkostninger $0.021/µg protein (bortset fra udgifter til arbejdskraft og udstyr), at gøre vores system til konkurrencedygtige priser med kommercielle kits uden at gå på kompromis lethed af reaktion setup. Vurderinger af sammenlignende omkostninger pr. µL af reaktion viser lignende tendenser (supplerende figur 1). Vi anslår start omkostninger for at være ~ $4.500 for alle reagenser, og en yderligere $3.200 for specialiseret udstyr, såsom en sonikator. Person timer at fuldføre denne procedure er anslået til at være ~ 26 h for alle reagens prep fra jorden op. Men når store lagre af reagenser har været forberedt, krav på arbejdskraft mindskes væsentligt. Derudover som er erfaringerne med platformen, vi anbefaler optrapning af størrelsen af cellevækst, uddrag forberedelse og reagens til at maksimere tidseffektivitet. Da start omkostninger, vi anbefaler CFPS platform for programmer i syntetisk biologi, høj overførselshastighed bestræbelser, og protein udtryk betingelser, der er uforenelig med traditionelle protein udtryk platforme skyldes er i strid med cellen biokemi og levedygtighed begrænsninger. I disse specialiserede sager hvor den ønskede teknik er aktiveret ved fælles fiskeripolitiks platform, er større udgifter til CFPS over i vivo udtryk berettiget.
Fortsat udvikling af fælles fiskeripolitiks platform forventes at give bredere nytte hen til bioteknologi bestræbelser som den metaboliske engineering af enzymatisk stier, produktion og karakterisering af traditionelt genstridig proteiner, nonstandard aminosyre vedtægter og unaturlige protein udtryk, stratificeret medicin fremstilling og udvide sig ud over laboratoriet ind i klasseværelset for STEM Uddannelse64,65,66. Disse bestræbelser vil støttes yderligere af de igangværende indsats for detaljerede karakterisering af fælles fiskeripolitiks platform. En bedre forståelse af sammensætningen af celle ekstrakt vil føre til fortsatte raffinement mod forbedret reaktion udbytter og fleksibilitet i reaktion betingelser61,67,68.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer, at de har ingen konkurrerende finansielle interesser eller andre interessekonflikter.
Acknowledgments
Forfatterne vil gerne anerkende Dr. Jennifer VanderKelen, Andrea Laubscher, og Tony Turretto for teknisk support, Wesley Kao, Layne Williams og Christopher Hight til nyttige diskussioner. Forfatterne også anerkende finansieringen støtte fra Bill og Linda Frost fond, Center for programmer i bioteknologis Chevron bioteknologi anvendt forskning Endowment Grant, Cal Poly forskning, akademisk og kreative aktiviteter Grant Program (RSCA 2017), og National Science Foundation (NSF-1708919). MZL anerkender California State University Graduate Grant. MCJ anerkender hær forskning Office W911NF-16-1-0372, giver National Science Foundation MCB-1413563 og MCB-1716766, Air Force forskning laboratorium Center af Excellence Grant FA8650-15-2-5518, Defense trussel reduktion agenturet Grant HDTRA1-15-10052/P00001, David og Lucile Packard Foundation, Camille Dreyfus lærer-Scholar Program, Institut for energi BER Grant DE-SC0018249, menneskelige grænser Science Program (RGP0015/2017), DOE Joint Genome Institute ETOP Grant, og Chicago biomedicinsk konsortium med støtte fra Searle midler på Chicago Fællesskabet Trust for støtte.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth | ThermoFisher | 12795027 | |
| Tryptone | Fisher Bioreagents | 73049-73-7 | |
| Yeast Extract | Fisher Bioreagents | 1/2/8013 | |
| NaCl | Sigma-Aldrich | S3014-1KG | |
| Potassium Phosphate Dibasic | Sigma-Aldrich | 60353-250G | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P9791-500G | |
| D-Glucose | Sigma-Aldrich | G8270-1KG | |
| KOH | Sigma-Aldrich | P5958-500G | |
| IPTG | Sigma-Aldrich | I6758-1G | |
| Mg(OAc)2 | Sigma-Aldrich | M5661-250G | |
| K(OAc) | Sigma-Aldrich | P1190-1KG | |
| Tris(OAc) | Sigma-Aldrich | T6066-500G | |
| DTT | ThermoFisher | 15508013 | |
| tRNA | Sigma-Aldrich | 10109541001 | |
| Folinic Acid | Sigma-Aldrich | F7878-100MG | |
| NTPs | ThermoFisher | R0481 | |
| Oxalic Acid | Sigma-Aldrich | P0963-100G | |
| NAD | Sigma-Aldrich | N8535-15VL | |
| CoA | Sigma-Aldrich | C3144-25MG | |
| PEP | Sigma-Aldrich | 860077-250MG | |
| K(Glu) | Sigma-Aldrich | G1501-500G | |
| NH4(Glu) | MP Biomedicals | 02180595.1 | |
| Mg(Glu)2 | Sigma-Aldrich | 49605-250G | |
| Spermidine | Sigma-Aldrich | S0266-5G | |
| Putrescine | Sigma-Aldrich | D13208-25G | |
| HEPES | ThermoFisher | 11344041 | |
| Molecular Grade Water | Sigma-Aldrich | 7732-18-5 | |
| L-Aspartic Acid | Sigma-Aldrich | A7219-100G | |
| L-Valine | Sigma-Aldrich | V0500-25G | |
| L-Tryptophan | Sigma-Aldrich | T0254-25G | |
| L-Phenylalanine | Sigma-Aldrich | P2126-100G | |
| L-Isoleucine | Sigma-Aldrich | I2752-25G | |
| L-Leucine | Sigma-Aldrich | L8000-25G | |
| L-Cysteine | Sigma-Aldrich | C7352-25G | |
| L-Methionine | Sigma-Aldrich | M9625-25G | |
| L-Alanine | Sigma-Aldrich | A7627-100G | |
| L-Arginine | Sigma-Aldrich | A8094-25G | |
| L-Asparagine | Sigma-Aldrich | A0884-25G | |
| Glycine | Sigma-Aldrich | G7126-100G | |
| L-Glutamine | Sigma-Aldrich | G3126-250G | |
| L-Histadine | Sigma-Aldrich | H8000-25G | |
| L-Lysine | Sigma-Aldrich | L5501-25G | |
| L-Proline | Sigma-Aldrich | P0380-100G | |
| L-Serine | Sigma-Aldrich | S4500-100G | |
| L-Threonine | Sigma-Aldrich | T8625-25G | |
| L-Tyrosine | Sigma-Aldrich | T3754-100G | |
| Fisherbrand Premium Microcentrifuge Tubes: 2.0 mL | Fisher Scientific | 05-408-138 | |
| Fisherbrand Premium Microcentrifuge Tubes: 1.5 mL | Fisher Scientific | 05-408-129 | |
| Fisherbrand Premium Microcentrifuge Tubes: 0.6 mL | Fisher Scientific | 05-408-120 | |
| PureLink HiPure Plasmid Prep Kit | ThermoFisher | K210007 | |
| Ultrasonic Processor | QSonica | Q125-230V/50Hz | 3.175 mm diameter probe |
| Avanti J-E Centrifuge | Beckman Coulter | 369001 | |
| JLA-8.1000 Rotor | Beckman Coulter | 366754 | |
| 1L Centrifuge Tube | Beckman Coulter | A99028 | |
| Tunair 2.5L Baffeled Shake Flask | Sigma-Aldrich | Z710822 | |
| Microfuge 20 | Beckman Coulter | B30134 | |
| New Brunswick Innova 42/42R Incubator | Eppendorf | M1335-0000 | |
| Cytation 5 | BioTek | ||
| Strep-Tactin XT Starter Kit | IBA | 2-4998-000 | |
| pJL1-sfGFP | Addgene | 69496 | |
| BL21(DE3) | New England BioLabs |
References
- Jiang, L., Zhao, J., Lian, J., Xu, Z. Cell-free protein synthesis enabled rapid prototyping for metabolic engineering and synthetic biology. Synthetic and Systems Biotechnology. 3 (2), 90-96 (2018).
- Carlson, E. D., Gan, R., Hodgman, C. E., Jewett, M. C. Cell-free protein synthesis: Applications come of age. Biotechnology Advances. 30 (5), 1185-1194 (2012).
- Watanabe, M. Cell-Free Protein Synthesis for Structure Determination by X-ray Crystallography. Methods in molecular biology (Clifton, N.J). 607, 149-160 (2010).
- Martemyanov, K. A., Shirokov, V. A., Kurnasov, O. V., Gudkov, A. T., Spirin, A. S. Cell-Free Production of Biologically Active Polypeptides: Application to the Synthesis of Antibacterial Peptide Cecropin. Protein Expression and Purification. 21 (3), 456-461 (2001).
- Renesto, P., Raoult, D. From genes to proteins: in vitro expression of rickettsial proteins. Annals of the New York Academy of Sciences. 990, 642-652 (2003).
- Xu, Z., Chen, H., Yin, X., Xu, N., Cen, P. High-Level Expression of Soluble Human b-Defensin-2 Fused With Green Fluorescent Protein in Escherichia coli Cell-Free System. Applied Biochemistry and Biotechnology. 127 (1), 053-062 (2005).
- Baumann, A. In-situ observation of membrane protein folding during cell-free expression. PLoS ONE. 11 (3), 1-15 (2016).
- Wang, Y., Percival, Y. H. P. Cell-free protein synthesis energized by slowly-metabolized maltodextrin. BMC Biotechnology. 9, 1-8 (2009).
- Whittaker, J. W. Cell-free protein synthesis: the state of the art. Biotechnology Letters. 35 (2), 143-152 (2013).
- Martin, R. W. Cell-free protein synthesis from genomically recoded bacteria enables multisite incorporation of noncanonical amino acids. Nature Communications. 9 (1), 1203 (2018).
- Kwon, Y. C., Song, J. K., Kim, D. M. Cloning-Independent Expression and Screening of Enzymes Using Cell-Free Protein Synthesis Systems. Methods in Molecular Biology. 1118, 97-108 (2014).
- Chappell, J., Jensen, K., Freemont, P. S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Research. 41 (5), 3471-3481 (2013).
- Takahashi, M. K. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods. 86, 60-72 (2015).
- Karim, A. S., Jewett, M. C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metabolic Engineering. 36, 116-126 (2016).
- Dudley, Q. M., Anderson, K. C., Jewett, M. C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synthetic Biology. 5 (12), 1578-1588 (2016).
- Pardee, K. Paper-based synthetic gene networks. Cell. 159 (4), 940-954 (2014).
- Zawada, J. F. Microscale to manufacturing scale-up of cell-free cytokine production-a new approach for shortening protein production development timelines. Biotechnology and Bioengineering. 108 (7), 1570-1578 (2011).
- Sullivan, C. J. A cell-free expression and purification process for rapid production of protein biologics. Biotechnology Journal. 11 (2), 238-248 (2016).
- Li, J. Cell-free protein synthesis enables high yielding synthesis of an active multicopper oxidase. Biotechnology Journal. 11 (2), 212-218 (2016).
- Heinzelman, P., Schoborg, J. A., Jewett, M. C. pH responsive granulocyte colony-stimulating factor variants with implications for treating Alzheimer's disease and other central nervous system disorders. Protein Engineering Design and Selection. 28 (10), 481-489 (2015).
- Pardee, K. Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell. 165 (5), 1255-1266 (2016).
- Slomovic, S., Pardee, K., Collins, J. J. Synthetic biology devices for in vitro and in vivo diagnostics. Proceedings of the National Academy of Sciences. 112 (47), 14429-14435 (2015).
- Gootenberg, J. S. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 356 (6336), 438-442 (2017).
- Pardee, K. Portable, On-Demand Biomolecular Manufacturing. Cell. 167 (1), 248-259 (2016).
- Karig, D. K., Bessling, S., Thielen, P., Zhang, S., Wolfe, J. Preservation of protein expression systems at elevated temperatures for portable therapeutic production. Journal of the Royal Society, Interface. 14 (129), (2017).
- Smith, M. T., Berkheimer, S. D., Werner, C. J., Bundy, B. C. Lyophilized Escherichia coli-based cell-free systems for robust, high-density, long-term storage. BioTechniques. 56 (4), 186-193 (2014).
- Hunt, J. P., Yang, S. O., Wilding, K. M., Bundy, B. C. The growing impact of lyophilized cell-free protein expression systems. Bioengineered. 8 (4), 325-330 (2017).
- Stark, J. C. BioBitsTM Bright: A fluorescent synthetic biology education kit. Science Advances. 4 (8), (2018).
- Huang, A. BioBitsTM Explorer: A modular synthetic biology education kit. Science Advances. 4 (8), (2018).
- Zemella, A., Thoring, L., Hoffmeister, C., Kubick, S. Cell-Free Protein Synthesis: Pros and Cons of Prokaryotic and Eukaryotic Systems. ChemBioChem. 16 (17), 2420-2431 (2015).
- Oza, J. P. Robust production of recombinant phosphoproteins using cell-free protein synthesis. Nature Communications. 6 (1), 8168 (2015).
- Zemella, A. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Scientific Reports. 8 (1), 8514 (2018).
- Jaroentomeechai, T. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nature Communications. 9 (1), 2686 (2018).
- Kightlinger, W. Design of glycosylation sites by rapid synthesis and analysis of glycosyltransferases. Nature Chemical Biology. 14 (6), 627-635 (2018).
- Schoborg, J. A. A cell-free platform for rapid synthesis and testing of active oligosaccharyltransferases. Biotechnology and Bioengineering. 115 (3), 739-750 (2018).
- Chekulayeva, M. N., Kurnasov, O. V., Shirokov, V. A., Spirin, A. S. Continuous-Exchange Cell-Free Protein-Synthesizing System: Synthesis of HIV-1 Antigen Nef. Biochemical and Biophysical Research Communications. 280 (3), 914-917 (2001).
- Hong, S. H. Improving Cell-Free Protein Synthesis through Genome Engineering of Escherichia coli Lacking Release Factor 1. ChemBioChem. 16 (5), 844-853 (2015).
- Endo, Y., Otsuzuki, S., Ito, K., Miura, K. Production of an enzymatic active protein using a continuous flow cell-free translation system. Journal of Biotechnology. 25 (3), 221-230 (1992).
- Volyanik, E. V., Dalley, A., Mckay, I. A., Leigh, I., Williams, N. S., Bustin, S. A. Synthesis of Preparative Amounts of Biologically Active Interleukin-6 Using a Continuous-Flow Cell-Free Translation System. Analytical Biochemistry. 214 (1), 289-294 (1993).
- Martin, G. A., Kawaguchi, R., Lam, Y., DeGiovanni, A., Fukushima, M., Mutter, W. High-yield, in vitro protein expression using a continuous-exchange, coupled transcription/ translation system. BioTechniques. 31 (4), 948-950 (2001).
- Stech, M., Quast, R. B., Sachse, R., Schulze, C., Wüstenhagen, D. A., Kubick, S. A Continuous-Exchange Cell-Free Protein Synthesis System Based on Extracts from Cultured Insect Cells. PLoS ONE. 9 (5), e96635 (2014).
- Quast, R. B., Sonnabend, A., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield cell-free synthesis of human EGFR by IRES-mediated protein translation in a continuous exchange cell-free reaction format. Scientific Reports. 6 (1), 30399 (2016).
- Thoring, L., Dondapati, S. K., Stech, M., Wüstenhagen, D. A., Kubick, S. High-yield production of "difficult-to-express" proteins in a continuous exchange cell-free system based on CHO cell lysates. Scientific Reports. 7 (1), 11710 (2017).
- Hoffmann, M., Nemetz, C., Madin, K., Buchberger, B. Rapid translation system: A novel cell-free way from gene to protein. Biotechnology annual review. 10, 1-30 (2004).
- Kwon, Y. C., Jewett, M. C. High-throughput preparation methods of crude extract for robust cell-free protein synthesis. Scientific Reports. 5 (1), 8663 (2015).
- Katsura, K. A reproducible and scalable procedure for preparing bacterial extracts for cell-free protein synthesis. Journal of Biochemistry. 162 (June), 357-369 (2017).
- Fujiwara, K., Doi, N. Biochemical preparation of cell extract for cell-free protein synthesis without physical disruption. PLoS ONE. 11 (4), 1-15 (2016).
- Shrestha, P., Holland, T. M., Bundy, B. C. Streamlined extract preparation for Escherichia coli-based cell-free protein synthesis by sonication or bead vortex mixing. BioTechniques. 53 (3), 163-174 (2012).
- Krinsky, N. A Simple and Rapid Method for Preparing a Cell-Free Bacterial Lysate for Protein Synthesis. PLoS ONE. 11 (10), (2016).
- Jewett, M. C., Swartz, J. R. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering. 86 (1), 19-26 (2004).
- Swartz, J. R., Jewett, M. C., Woodrow, K. A. Cell-Free Protein Synthesis With Prokaryotic Combined Transcription-Translation. Recombinant Gene Expression. (267), 169-182 (2004).
- Voloshin, A. M., Swartz, J. R. Efficient and scalable method for scaling up cell free protein synthesis in batch mode. Biotechnology and Bioengineering. 91 (4), 516-521 (2005).
- Vernon, W. B. The role of magnesium in nucleic-acid and protein metabolism. Magnesium. 7 (5-6), 234-248 (1988).
- Pratt, J. M. Transcription and Translation: A Practical Approach. , IRL Press. New York. (1984).
- Kim, D. M., Kigawa, T., Choi, C. Y., Yokoyama, S. A Highly Efficient Cell-Free Protein Synthesis System from Escherichia coli. European Journal of Biochemistry. 239 (3), 881-886 (1996).
- Shin, J., Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. Journal of Biological Engineering. 4 (1), 8 (2010).
- Zubay, G. In Vitro Synthesis of Protein in Microbial Systems. Annual Review of Genetics. 7 (1), 267-287 (1973).
- Kigawa, T. Preparation of Escherichia coli cell extract for highly productive cell-free protein expression. Journal of Structural and Functional Genomics. 5 (1/2), 63-68 (2004).
- Liu, D. V., Zawada, J. F., Swartz, J. R. Streamlining Escherichia Coli S30 Extract Preparation for Economical Cell-Free Protein Synthesis. Biotechnology Progress. 21 (2), 460-465 (2008).
- Yang, W. C., Patel, K. G., Wong, H. E., Swartz, J. R. Simplifying and streamlining Escherichia coli-based cell-free protein synthesis. Biotechnology Progress. 28 (2), 413-420 (2012).
- Foshag, D. The E. coli S30 lysate proteome: A prototype for cell-free protein production. New Biotechnology. 40 (Pt B), 245-260 (2018).
- Caschera, F. Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology. 2 (2), 97-104 (2017).
- Chizzolini, F., Forlin, M., Yeh Martín, N., Berloffa, G., Cecchi, D., Mansy, S. S. Cell-Free Translation Is More Variable than Transcription. ACS Synthetic Biology. 6 (4), 638-647 (2017).
- Hong, S. H., Kwon, Y. C., Jewett, M. C. Non-standard amino acid incorporation into proteins using Escherichia coli cell-free protein synthesis. Frontiers in Chemistry. 2, 34 (2014).
- Sawasaki, T., Ogasawara, T., Morishita, R., Endo, Y. A cell-free protein synthesis system for high-throughput proteomics. Proceedings of the National Academy of Sciences of the United States of America. 99 (23), 14652-14657 (2002).
- Dudley, Q. M., Karim, A. S., Jewett, M. C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnology Journal. 10 (1), 69-82 (2015).
- Garcia, D. C. Elucidating the potential of crude cell extracts for producing pyruvate from glucose. Synthetic Biology. 3 (1), 1-9 (2018).
- Hurst, G. B. Proteomics-Based Tools for Evaluation of Cell-Free Protein Synthesis. Analytical Chemistry. 89 (21), 11443-11451 (2017).

{kind=link}
{kind=link}
{kind=link}