

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
פרוטוקולים אלה יסייעו למשתמשים לחקור את חילוף החומרים של אנרגיה מיטוכונדריאלית בספרואידים שמקורם בתאי סרטן תלת-ממדיים באמצעות ניתוח שטף חוץ-תאי של סוסון ים.
Abstract
אגרגטים תאיים תלת-ממדיים (3D), המכונים ספרואידים, הפכו בשנים האחרונות לחזית תרבית התאים במבחנה . בניגוד לתאי התרבות כחד-שכבתיים דו-ממדיים חד-תאיים (תרבית דו-ממדית), תרבית תאים כדורית מקדמת, מווסתת ותומכת בארכיטקטורה תאית פיזיולוגית ובמאפיינים הקיימים ב-vivo, כולל ביטוי של חלבוני מטריצה חוץ-תאית, איתות תאים, ביטוי גנים, ייצור חלבונים, התמיינות והתפשטות. חשיבותה של תרבית תלת-ממדית הוכרה בתחומי מחקר רבים, כולל אונקולוגיה, סוכרת, ביולוגיה של תאי גזע והנדסת רקמות. בעשור האחרון פותחו שיטות משופרות לייצור ספרואידים ולהערכת תפקודם המטבולי וגורלם.
מנתחי שטף חוץ-תאי (XF) שימשו כדי לחקור את תפקוד המיטוכונדריה במיקרוטיסים תלת-ממדיים כגון ספרואידים באמצעות צלחת לכידת איים XF24 או מיקרו-פלטה ספרואידית XFe96. עם זאת, פרוטוקולים ברורים ואופטימיזציה של בדיקת חילוף החומרים של אנרגיה מיטוכונדריאלית בספרואידים באמצעות טכנולוגיית XF לא תוארו בפירוט. מאמר זה מספק פרוטוקולים מפורטים לבדיקת חילוף החומרים של אנרגיה מיטוכונדריאלית בספרואידים תלת-ממדיים בודדים באמצעות מיקרו-פלטות ספרואידיות עם מנתח XF XFe96. באמצעות קווי תאים סרטניים שונים, טכנולוגיית XF מוכחת כמי שמסוגלת להבחין בין נשימה תאית בספרואידים תלת-ממדיים בגדלים שונים לא רק בגדלים שונים, אלא גם בנפחים שונים, מספרי תאים, תכולת DNA וסוגים שונים.
ריכוזי התרכובת האופטימליים של אוליגומיצין, BAM15, רוטנון ואנטימיצין A משמשים לבדיקת פרמטרים ספציפיים של חילוף חומרים של אנרגיה מיטוכונדריאלית בספרואידים תלת-ממדיים. מאמר זה דן גם בשיטות לנרמול נתונים המתקבלים מספרואידים ומתייחס לשיקולים רבים שיש לקחת בחשבון בעת חקירת חילוף החומרים של הספרואידים באמצעות טכנולוגיית XF. פרוטוקול זה יסייע להניע מחקר במודלים מתקדמים של ספרואידים במבחנה .
Introduction
ההתקדמות במודלים חוץ גופיים במחקר הביולוגי התקדמה במהירות במהלך 20 השנים האחרונות. מודלים כאלה כוללים כיום אופנות של איברים על שבב, אורגנואידים וספרואידים תלת-ממדיים של מיקרוטיסואה, שכולם הפכו למוקד נפוץ לשיפור התרגום בין מחקרי in vitro ו-in vivo. השימוש במודלים מתקדמים במבחנה, במיוחד ספרואידים, משתרע על פני מספר תחומי מחקר, כולל הנדסת רקמות, מחקר בתאי גזע, סרטן וביולוגיה של מחלות 1,2,3,4,5,6,7, ובדיקות בטיחות, כולל טוקסיקולוגיה גנטית 8,9,10, טוקסיקולוגיה של ננו-חומרים11, 12,13,14, ובדיקות בטיחות ויעילות של תרופות 8,15,16,17,18,19.
מורפולוגיה תקינה של תאים היא קריטית לפנוטיפ ולפעילות ביולוגית. גידול תאים לכדי ספרואידים תלת-ממדיים של מיקרוטיסואה מאפשר לתאים לאמץ מורפולוגיה, פונקציה פנוטיפית וארכיטקטורה, הדומים יותר לאלה שנצפו in vivo אך קשים ללכידה באמצעות טכניקות קלאסיות של תרביות תאים חד-שכבתיות יותר. הן in vivo והן in vitro, תפקוד התאים מושפע ישירות מהמיקרו-סביבה התאית, שאינה מוגבלת לתקשורת תאית ותכנות (למשל, תצורות צומת תאים-תאים, הזדמנויות ליצור נישות תאים); חשיפת תאים להורמונים ולגורמי גדילה בסביבות המיידיות (למשל, חשיפה לציטוקין תאי כחלק מתגובה דלקתית); הרכב של מטריצות פיזיקליות וכימיות (למשל, האם תאים גדלים בפלסטיק בתרבית רקמה נוקשה או בסביבת רקמה אלסטית); והכי חשוב, כיצד חילוף החומרים התאי מושפע מתזונה וגישה לחמצן, כמו גם מעיבוד של תוצרי פסולת מטבולית כגון חומצה לקטית.
ניתוח שטף מטבולי הוא דרך רבת עוצמה לבחון את חילוף החומרים התאי בתוך מערכות מוגדרות במבחנה . באופן ספציפי, טכנולוגיית XF מאפשרת ניתוח של שינויים חיים בזמן אמת בביו-אנרגיה תאית של תאים ורקמות שלמים. בהתחשב בכך שאירועים מטבוליים תוך-תאיים רבים מתרחשים בסדר גודל של שניות עד דקות, גישות תפקודיות בזמן אמת הן בעלות חשיבות עליונה להבנת שינויים בזמן אמת בשטף המטבולי התאי בתאים שלמים וברקמות במבחנה.
מאמר זה מספק פרוטוקולים לטיפוח קווי תאים שמקורם בסרטן A549 (אדנוקרצינומה של הריאות), HepG2/C3A (קרצינומה הפטוצלולרית), MCF-7 (אדנוקרצינומה של השד) ו-SK-OV-3 (אדנוקרצינומה שחלתית) כמו במודלים ספרואידיים תלת-ממדיים במבחנה תוך שימוש בגישות של צבירה כפויה (איור 1). הוא גם (i) מתאר בפירוט כיצד לחקור את חילוף החומרים של אנרגיה מיטוכונדריאלית של כדוריות תלת-ממדיות בודדות באמצעות מנתח Agilent XFe96 XF, (ii) מדגיש דרכים לייעל את מבחני ה-XF באמצעות ספרואידים תלת-ממדיים בודדים, ו-(iii) דן בשיקולים חשובים ובמגבלות של בדיקת חילוף החומרים הספרואידי התלת-ממדי באמצעות גישה זו. והכי חשוב, מאמר זה מתאר כיצד נאספים מערכי נתונים המאפשרים חישוב של קצב צריכת החמצן (OCR) כדי לקבוע זרחון חמצוני ובכך את תפקוד המיטוכונדריה בספרואידים תאיים. למרות שלא נותח עבור פרוטוקול זה, קצב ההחמצה החוץ-תאית (ECAR) הוא פרמטר נוסף הנמדד לצד נתוני OCR בניסויי XF. עם זאת, ECAR מתפרש לעתים קרובות בצורה גרועה או שגויה ממערכי נתונים של XF. אנו מספקים פרשנות לגבי המגבלות של חישוב ECAR בעקבות גישות בסיסיות של יצרן הטכנולוגיה.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

איור 1: זרימת עבודה גרפית ליצירת ספרואידים תאיים, ניתוח שטף חוץ-תאי ובדיקות במורד הזרם. ארבעה קווי תאים סרטניים עברו תרבית סלקטיבית כחד-שכבתיים (A), נותקו מצלוחיות תרביות רקמה, ונזרעו לתוך מיקרו-פלטות חיבור אולטרה-נמוכות בעלות 96 בארות ליצירת ספרואידים (B). קרצינומה של ריאה A549, קרצינומה בכבד HepG2/C3A, אדנוקרצינומה שחלתית SK-OV-3 ותאי קרצינומה של השד MCF-7 נזרעו ב-1 × 103-8 × 103 תאים/באר וגדלו עד 7 ימים כדי ליצור ספרואידים בודדים ולייעל את צפיפות הזריעה הספרואידית ואת זמן הטיפוח על ידי תצפית רציפה ומדידות פלנימטריות. לאחר שנוצרו, ספרואידים בודדים נשטפו לתוך מדיום XF ללא סרום ונזרעו בקפידה לתוך מיקרו-לוחות בדיקה ספרואידיים, מצומדים מראש עם פולי-D-ליזין (C). הספרואידים היו נתונים לניתוח שטף חוץ-תאי באמצעות מנתח XFe96 תוך שימוש במספר פרוטוקולים כדי לטפל ב: (1) גודל ספרואידי אופטימלי לתגובת נשימה מיטוכונדריאלית בסיסית; (2) טיטרציה אופטימלית של מעכבי נשימה מיטוכונדריאליים; (3) אופטימיזציה של מיקום הספרואידים בתוך בארות מיקרו-פלטה. (D) ניתוחים לאחר XF, מיקרוסקופיית ניגודיות פאזה וכימות DNA ספרואידי שימשו לנורמליזציה של נתונים ולבדיקות אחרות במבחנה במורד הזרם. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
1. טיפוח קווי תאים סרטניים כספרואידים תלת-ממדיים במבחנה
| קו סלולרי | תיאור | מדיום תרבותי | מקור |
| A549 | קו תאי קרצינומה של הריאות | RPMI 1640 | אוסף אירופי של תרביות תאים מאומתות (ECACC) |
| נתרן פירובט (1 מ"מ) | |||
| פניצילין- סטרפטומיצין - (100 U/mL - 100 מ"ג/מ"ל) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | קו תאי קרצינומה בכבד, נגזרת קלונלית של קו תאי ההורה HepG2 | DMEM | אוסף תרביות רקמה אמריקאי (ATCC) |
| פניצילין- סטרפטומיצין - (100 U/mL - 100 מ"ג/מ"ל) | |||
| 10 % (v/v) FBS | |||
| MCF7 | קו תאי אדנוקרצינומה של השד | RPMI 1640 | אוסף אירופי של תרביות תאים מאומתות (ECACC) |
| נתרן פירובט (1 מ"מ) | |||
| פניצילין- סטרפטומיצין - (100 U/mL - 100 מ"ג/מ"ל) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | קו תאי אדנוקרצינומה שחלתית | RPMI 1640 | אוסף אירופי של תרביות תאים מאומתות (ECACC) |
| נתרן פירובט (1 מ"מ) | |||
| פניצילין- סטרפטומיצין - (100 U/mL - 100 מ"ג/מ"ל) | |||
| 10 % (v/v) FBS | |||
| רכיב | מדיום בדיקת RPMI (נפח סופי של 50 מ"ל) | ||
| מדיום בסיס | סוסון ים אג'ילנט XF RPMI, pH 7.4 | ||
| גלוקוז (1 M ציר סטרילי) | 11 mM (תמיסת מלאי של 0.55 מ"ל) | ||
| L-גלוטמין (200 mM מלאי סטרילי) | 2 mM (0.5 מ"ל של תמיסת מלאי) | ||
| נתרן פירובט (ציר סטרילי של 100 mM) | 1 mM (0.5 מ"ל של תמיסת מלאי) |
טבלה 1: מדיה של קווי תאים סרטניים והרכבי מדיה XF.
- תרבית את כל קווי התאים באמצעות טכניקת תרבית רקמה אספטית סטנדרטית ואישר שהם נקיים ממיקופלסמה באמצעות ערכת בדיקה מתאימה.
- תרבית את קווי התאים בבקבוקי תרבית רקמה T75 או שווה ערך, באמצעות המדיום המומלץ (טבלה 1). תרבית את קווי התא למפגש של 65-80% והעברתם באופן קבוע עד למקסימום של 25 מעברים.
- יש לשטוף פעמיים את צלוחיות תרבית התאים במי מלח (DBPS) שעברו DBPS של דולבקו.
- נתקו את התאים מהצלליות עם 3 מ"ל של מגיב הדיסוציאציה של התא (ראו טבלת החומרים) למשך 5 דקות בטמפרטורה של 37 מעלות צלזיוס ואישרו את הניתוק על ידי מיקרוסקופיה.
- שאפו בעדינות את תרחיף התא המנותק כדי להבטיח תרחיף חד-תאי ולהשבית את ריאגנט הדיסוציאציה של התא עם 7 מ"ל של מדיום תרבית רקמה שלם.
- לאסוף את התאים על ידי צנטריפוגה ב 300 × g במשך 5 דקות, להשליך את supernatant, ולהחיות את התאים בתווך שלם.
- ספרו את התאים באמצעות המוציטומטר או מונה תאים אוטומטי והצמידו את צפיפות התאים הרצויה לזריעה.
הערה: כדי לזרוע צלחת שלמה של 96 בארות ב-100 μL/well ב-4 ×-103 תאים/באר, התאים צריכים להיות מחוברים ל-4 ×-104 תאים/מ"ל בנפח מומלץ של 12 מ"ל. - נקו את תרחיף התא למאגר סטרילי ופיצו 100 μL של תרחיף התא לכל באר של מיקרו-לוחית דוחה תאים באמצעות פיפטטור רב-ערוצי.
הערה: יש לזרוע רק את 60 הבארות הפנימיות של מיקרו-לוחית ואת השאר למלא ב-DPBS. פעולה זו תיצור מחסום אידוי, תבטיח הומוגניות כדורית על פני הצלחת ותמזער את השפעות קצה הצלחת. - מיקרו-לוחות ספרואידים של צנטריפוגה ב-300 × גרם במשך 15 דקות כדי לאלץ את התאים לאגרגטים רופפים.
- הדגירה של הצלחות בטמפרטורה של 37 מעלות צלזיוס, 5% CO2 למשך 3 ימים לפחות כדי להבטיח היווצרות כדורית.
- בצע מיקרוסקופיה של ניגודיות פאזה תוך שימוש בשיטות מעבדה סטנדרטיות כדי לעקוב אחר צמיחתם של ספרואידים. מלאו את מדיום תרביות התאים כל 3 ימים או פעמיים בשבוע על ידי ביצוע חילופי חצי נפח בינוניים.
2. בדיקת חילוף חומרים של אנרגיה מיטוכונדריאלית של כדוריות בודדות באמצעות טכנולוגיית שטף חוץ-תאי (XF)
- הכנת בדיקה (יום אחד לפני כן)
- בדוק את הכדאיות הספרואידית באמצעות מיקרוסקופ אור הפוך עם ניגודיות פאזה בהגדלה של פי 4 כדי להבטיח מבנה ספרואידי שלם, מורפולוגיה ואחידות כוללת בין הדגימות.
- הוסיפו לחות למחסנית החיישן.
- Aliquot ~ 20 מ"ל של calibrant לתוך צינור חרוטי.
- מניחים את הצינור החרוטי המכיל את הקליברנט באינקובטור שאינו CO2 37 °C למשך הלילה.
- הסר את התוכן מתוך ערכת הבדיקה.
- הסר את מחסנית החיישן מלוח השירות והנח אותה במהופך על משטח העבודה שליד לוח השירות.
- Pipette 200 μL של ddH2O סטרילי לתוך כל באר של לוחית השירות של מחסנית החיישן באמצעות פיפטה P300 רב-ערוצית.
- הניחו את מחסנית החיישן על גבי לוחית השירות.
- בדקו שמפלס המים בכל באר גבוה מספיק כדי להטביע את בדיקות החיישן.
- העבר את מחסנית החיישן המורכבת לחממה שאינה CO2 37 °C והשאר אותה למשך הלילה.
הערה: ניתן לבצע שלב זה 12-72 שעות לפני תחילת הבדיקה.
- מעיל כדורי בדיקה מיקרו-לוחית
- באמצעות טכניקות אספטיות, הוסיפו 30 μL/באר של תמיסת Poly-D-Lysine סטרילית (0.1 מ"ג/מ"ל) למיקרו-פלטה הספרואידית והדגמו אותה למשך 30 דקות בטמפרטורת החדר.
- שאפו את התמיסה מכל באר של המיקרו-פלטה הספרואידית, הפכו את הצלחת והקשו אותה בחוזקה על נייר טישו כדי להסיר כל תמיסה שיורית.
- שטפו את הצלחת פעמיים עם 200 μL/באר של ddH2O סטרילי.
- לאחר הכביסה הסופית, הפכו את המיקרו-פלטה והקשו אותה בחוזקה על נייר טישו כדי להסיר את שאריות המים.
- אפשרו לצלחת להתייבש באוויר למשך 30 דקות לפני השימוש או האחסון שלה ב-4 מעלות צלזיוס לשימוש עתידי.
הערה: יש לצפות את המיקרו-פלטה של הבדיקה הספרואידית בדבק מולקולרי כדי להבטיח שהספרואידים מקובעים בתחתית המיקרו-פלטה. ללא דבק מולקולרי, ספרואידים יכולים להתנתק ולהפריע לתוצאות הבדיקה. דבקים מולקולריים אחרים יכולים לשמש גם כחלופה לפולי-D-ליזין עבור לוחות מקדימים. ניתן לאחסן צלחות מוכנות מראש בטמפרטורה של 4 מעלות צלזיוס, אך יש להשאירן לשיווי משקל לטמפרטורת החדר לפני תחילת הבדיקה.
- הכן XF Assay בינוני
- הכן מדיום XF RPMI, כמפורט בטבלה 1, ומסנן סטרילי עם מסנן מזרק 0.22 מיקרומטר
- הכנת בדיקה (שעה אחת לפני הבדיקה)
- יש להמתין מראש את המבחן הנוסף של XF RPMI בינוני ל-37 מעלות צלזיוס.
- לפני המלחמה את המיקרו-פלטה של בדיקת הספרואידים המצופה באינקובטור שאינו CO2 37 °C או באמבטיה יבשה.
- הכן את מחסנית החיישן.
- הוציאו את הצינור החרוטי המכיל את הקליברנט ואת מחסנית החיישן מחממת האוויר.
- הסר את מחסנית החיישן מלוח השירות והנח אותה במהופך על משטח העבודה.
- באמצעות פיפטה רב-ערוצית P300, שאפו את המים מלוח השירות והשליכו אותם.
- יוצקים את תמיסת הקליברנט למאגר ריאגנטים סטרילי ומוסיפים 200 μL/באר של הקליברנט המתוכנן מראש ללוח השירות באמצעות פיפטה רב-ערוצית P300.
- הרימו את מחסנית החיישן והניחו אותה בחזרה על גבי לוח השירות, מה שמבטיח שהחיישנים שקועים היטב בקליברנט.
- העבר את מחסנית החיישן המורכבת בחזרה לחממה שאינה CO2 37 °C עד שיהיה מוכן לטעון את פתרונות הזרקת היציאה.
- לשטוף את הספרואידים עם מדיום הבדיקה.
- הסר את צלחת תרבית הספרואידים מחממת 37 °C, 5% CO2 והתבונן בספרואידים מתחת למיקרוסקופ כדי להבטיח את שלמותם לפני שלבי העברת הספרואידים.
- טען את כל הבארות של הלוח הספרואידי עם 180 μL / באר של מדיום בדיקה מוכן מראש, כולל כל בארות תיקון רקע.
- ממלאים חלקית צלחת פטרי 7 ס"מ ב-3 מ"ל של מדיום הבדיקה.
- באמצעות פיפטה רב-ערוצית עמוסה בקצות פיפטה רחבים, מעבירים את הספרואידים מצלחת התרבית בת 96 הבארות לכלי פטרי בקוטר 7 ס"מ על ידי הצבת הפיפטטור בנפח שאיפה של 10-50 מיקרול'.
- כדוריות זרעים לתוך מיקרו-לוחית הבדיקה הספרואידית המצופה מראש.
- באמצעות מיקרוסקופ דיסקציה ומנגנון תיבת אור, מעבירים את הספרואידים מצלחת פטרי למיקרו-לוחית הבדיקה הספרואידית כמפורט להלן.
- הגדר את הנפח של פיפטטור בעל ערוץ יחיד המצויד בקצה פיפטה רחב ל-20 μL ושאף בזהירות ספרואיד יחיד. מניחים את הקצה ישירות במרכז כל באר של מיקרו-לוחית הבדיקה הספרואידית ומאפשרים לכוח הכבידה להכניס ספרואיד יחיד למרכז כל באר, כלומר, אל תגרשו שום מדיום מקצה הפיפטה ותאפשרו לפעולה נימית למשוך את הספרואיד מקצה הפיפטה. כדי לאשר את האלוטציה, ניתן לטפטף את תכולת הפיפטטור בחזרה לתוך צלחת פטרי 7 ס"מ מתחת למיקרוסקופ.
הערה: אלוטציה של ספרואיד יחיד לוקחת בדרך כלל 15-30 שניות בהתאם לגודל/צפיפות הספרואידים. במהלך תקופה זו, אין להסיר את הצינור. כל בארות תיקון רקע צריכות להיות נקיות מספרואידים ולהכיל רק מדיום בדיקה. תחת המיקרוסקופ, לאשר את המיקום של כל ספרואיד. כל ספרואיד צריך להיות ממוקם באופן אידיאלי במרכז כל באר. - לאחר שכל הספרואידים הועברו למיקרו-לוחית הבדיקה של הספרואידים, העבירו את הצלחת לחממה שאינה CO2 בטמפרטורה של 37 מעלות צלזיוס למשך שעה לפחות 1 שעות לפני הבדיקה.
- הגדר את הנפח של פיפטטור בעל ערוץ יחיד המצויד בקצה פיפטה רחב ל-20 μL ושאף בזהירות ספרואיד יחיד. מניחים את הקצה ישירות במרכז כל באר של מיקרו-לוחית הבדיקה הספרואידית ומאפשרים לכוח הכבידה להכניס ספרואיד יחיד למרכז כל באר, כלומר, אל תגרשו שום מדיום מקצה הפיפטה ותאפשרו לפעולה נימית למשוך את הספרואיד מקצה הפיפטה. כדי לאשר את האלוטציה, ניתן לטפטף את תכולת הפיפטטור בחזרה לתוך צלחת פטרי 7 ס"מ מתחת למיקרוסקופ.
- באמצעות מיקרוסקופ דיסקציה ומנגנון תיבת אור, מעבירים את הספרואידים מצלחת פטרי למיקרו-לוחית הבדיקה הספרואידית כמפורט להלן.
3. הכנה וטעינה של תרכובות למחסנית החיישן למבחני XF
| אסטרטגיית הזרקה | מתחם (יציאה) | נפח התחלתי של מיקרו-וול XFe96 (μL) | ריכוז באר סופית רצויה | נפח יציאה (μL) | נפח מיקרווול XFe96 סופי לאחר ההזרקה (μL) | ריכוז מלאי עבודה |
| 1 | אוליגומיצין (אנ') | 180 | 3 מ"ג/מ"ל | 20 | 200 | 30 מיקרוגרם/מ"ל |
| רוטנונה (ב) | 200 | 2 מיקרומטר | 20 | 220 | 22 מיקרומטר | |
| אנטימיצין A (B) | 200 | 2 מיקרומטר | 20 | 220 | 22 מיקרומטר | |
| 2 | BAM15 (A) | 180 | 5 מיקרומטר | 20 | 200 | 50 מיקרומטר |
| רוטנונה (ב) | 200 | 2 מיקרומטר | 20 | 220 | 22 מיקרומטר | |
| אנטימיצין A (B) | 200 | 2 מיקרומטר | 20 | 220 | 22 מיקרומטר |
טבלה 2: ריכוזי תרכובות מיטוכונדריאליות לבדיקת חילוף החומרים של אנרגיה מיטוכונדריאלית של ספרואידים תלת-ממדיים בודדים באמצעות מנתח XFe96.
- הכן ריכוזי מלאי עבודה של כל תרכובת כפי שצוין בטבלה 2 באמצעות מדיום בדיקת XF RPMI מלא, מוכן מראש.
- כוון את לוחית המחסנית (המוצמדת ללוח השירות) מבחינת עמודה, 1-12 משמאל לימין.
- אם אתה משתמש במדריך טעינה, הנח אותו על גבי לוחית המחסנית בהתאם להליך הטעינה היטב, לדוגמה, אם נטען תחילה יציאה A, ודא ש- A גלוי בפינה השמאלית העליונה של המדריך.
- העבר את פתרון העבודה של כל תרכובת למאגר מתאים, ובאמצעות פיפטה רב-ערוצית מכוילת P100, מחלק 20 μL לכל היציאות המתאימות. יש לחזור על הפעולה עבור כל תרכובת לתוך היציאות הנותרות.
הערה: אם לא נעשה שימוש ביציאות כלשהן בלוח מחסנית החיישן, ניתן להשאיר אותן ריקות או מלאות במדיום בדיקה. אם נעשה שימוש רק בבחירה של אות יציאה מסוימת, ודא שהיציאות האחרות המתאימות לאות זו נטענות במדיום בדיקה; אחרת, אוויר יוזרק לתוך הבאר, תוך פגיעה בתוצאות בארות אלה. - לאחר טעינת היציאה, הסר את מדריכי טעינת הצלחת (אם נעשה בהם שימוש) והכן את המנתח לטעינת מחסנית החיישן.
הערה: אם הבדיקה אינה מופעלת מיד לאחר טעינת היציאות, הנח את המכסה בחזרה על מחסנית החיישן והחזיר את הצלחת לחממת האוויר של 37 °C עד שיהיה מוכן לטעינה למכונה.
4. תכנון בדיקה, אסטרטגיות הזרקה ורכישת נתונים
- הפעלת הבדיקה
- הפעל את המנתח והתחבר לבקר (מחשב).
הערה: ניתן לאמת זאת על-ידי מצב חיבור המכשיר בחלונית הווידג'טים של תוכנת Wave Controller. - נווט אל דף התבניות בתוכנת WAVE, מצא את קובץ תבנית הבדיקה של הניסוי ולחץ פעמיים כדי לפתוח אותו.
הערה: אם תבנית הבדיקה אינה מופיעה בתצוגה תבניות , ייבא את קובץ התבנית לתיקיית התבנית מכונן רשת משותף או מכונן הבזק מסוג USB. - כדי להתחיל את הבדיקה, לחץ על הכרטיסיה הפעל Assay .
הערה: אם הגדרות הקבוצה הוקצו כהלכה בתוך מפת הלוח, הבדיקה תהיה מוכנה להפעלה כפי שמצוין על ידי הסימון הירוק בצד ימין של הדף. בשלב זה, ניתן להזין כל מידע נוסף בדף סיכום הבדיקה או בדף שנותר ריק; המשך לשלב הבא. בשל החדירה המאוחרת של אפננים מיטוכונדריאליים בספרואידים תלת-ממדיים של מיקרוטיסואה (איור 2), השתמש במידע פרוטוקול המדידה המתואר בטבלה 3.
- הפעל את המנתח והתחבר לבקר (מחשב).
| תקופת המדידה | מספר הזרקה ויציאה | פרטי המדידה | משך התקופה (h:min:s) |
| כיול | לא ישים | מנתחי XF מבצעים תמיד כיול זה כדי לוודא שהמדידות מדויקות | 00:20:00 (זה ממוצע ויכול להשתנות בין מכונות) |
| שוויון נפש | לא ישים | שיווי משקל מתרחש לאחר כיול והוא מומלץ. | 00:10:00 |
| בזלת | לא ישים | מחזורים = 5 | 00:30:00 |
| מיקס = 3:00 | |||
| המתן = 0:00 | |||
| מידה = 3:00 | |||
| אוליגומיצין / BAM15 | הזרקה 1 (יציאה A) | מחזורים = 10 | 01:00:00 |
| מיקס = 3:00 | |||
| המתן = 0:00 | |||
| מידה = 3:00 | |||
| רוטנון + אנטימיצין A | הזרקה 2 (יציאה B) | מחזורים = 10 | 01:00:00 |
| מיקס = 3:00 | |||
| המתן = 0:00 | |||
| מידה = 3:00 | |||
| סה"כ זמן: | 03:00:00 | ||
טבלה 3: הגדרת פרוטוקול לבדיקת חילוף חומרים של אנרגיה מיטוכונדריאלית של ספרואידים תלת-ממדיים בודדים באמצעות מנתח XFe96.
- לחץ על התחל הפעל כדי להעלות את תיבת הדו-שיח שמירת מיקום .
- הזן את מיקום השמירה של קובץ התוצאה, והנח את מחסנית החיישן המורכבת על המגש התרמי המופיע מהדלת בצד המנתח. המתן עד שהמגש התרמי ייפתח באופן אוטומטי והמסך יציג את הודעת לוח השירות של Load Calibrant . לפני ביצוע ההנחיות על המסך, ודא i) התאמה נכונה של מחסנית החיישן בלוח השירות, ii) המכסה מוסר ממחסנית החיישן, ו- iii) כיוון נכון של מחסנית החיישן בלוח השירות.
- בצע את הפקודות שעל המסך כדי להפעיל כיול מחסנית חיישן.
הערה: הזמן שלוקח להשלים את הכיול הוא בערך 10-20 דקות (לבדיקות ב-37 מעלות צלזיוס). - לאחר כיול מחסנית החיישן, טען את המיקרו-פלטה הספרואידית לתוך המנתח על-ידי ביצוע ההוראות שעל המסך בבקר הגלים כדי להתחיל את שלב שיווי המשקל של 12 דקות.
הערה: תיבות ירוקות עם קרציות לבנות מציינות כיול 'טוב' עבור באר זו. אם בארות כלשהן לא יצליחו לספק כיול 'טוב', הן יסומנו בקופסה אדומה ובצלב לבן. יש לציין בארות כאלה ולהחריג אותן מכל ניתוח לאחר השלמת הבדיקה באמצעות כרטיסיית בדיקת השינוי . - המתן עד שהמנתח יתחיל באופן אוטומטי לרכוש מדידות בסיסיות לאחר שהמכונה השלימה את שלב שיווי המשקל (כמתואר בפרוטוקול המכשיר).
- כדי להשלים את הניסוי, בצע את הפקודות שעל המסך בבקר WAVE.
הערה: לאחר שהמיקרו-פלטה הספרואידית הוסרה מהמנתח, השליכו את מחסנית החיישן והניחו בצד את הלוחית הספרואידית לניתוח נוסף במידת הצורך (למשל, כימות דנ"א דו-גדילי (ds). אם המיקרו-פלטה אינה נדרשת לניתוח נוסף, ניתן להשליך אותה יחד עם מחסנית החיישן. - המתן עד שתיבת הדו-שיח של הבדיקה תופיע ותציג את התוצאות או חזור לתצוגת התבניות .
5. אסטרטגיות לנורמליזציה וניתוח נתונים - נורמליזציה לאחר הבדיקה ומבחנים במורד הזרם (שלבים אופציונליים)
- נורמליזציה של נתונים
- כדי לנרמל את נתוני הספרואידים, עיין בסדרת הפרוטוקולים הרלוונטיים לאסטרטגיות נורמליזציה של נתונים לחישוב הגודל והנפח של הספרואידים ולכימות dsDNA במבחנים ספרואידיים. אלה נכללו כקבצים משלימים; ראה קובץ משלים 1 וקובץ משלים 2.
- ניתוח נתונים
- כדי לייצא נתונים לאחד ממחוללי הניתוח האוטומטיים, בצע את פקודות ייצוא הנתונים בבקר WAVE ובחר את מחולל הייצוא התואם לסוג הבדיקה. לחלופין, ייצא את קובץ הנתונים והעלה אותו לניתוח סוסון ים.
הערה: החיסרון של מחוללי דוחות וניתוח סוסון ים הוא שניתוח נתונים מוגבל לאופן שבו מתוכנן מבחן ה- XF ואינו מאפשר לקחת ממוצעים על פני מחזורי מדידה. ייצוא ידני של מערכי נתונים מתוכנת המכשיר מאפשר העדפת משתמש בהקשר זה. בהתחשב בכך שאסטרטגיית ההזרקה להערכת נשימה מיטוכונדריאלית של ספרואידים תלת-ממדיים תהיה ככל הנראה שונה מזו של בדיקת 'MitoStress' טיפוסית, פותחו סדרה של תבניות גיליון אלקטרוני כדי לסייע בניתוח מערכי נתונים אלה, ספציפיים לתרביות תאים תלת-ממדיות, ויסופקו על פי בקשה. קבצי תבניות נתונים אלה יספקו נתונים על הפרמטרים הנשימתיים המיטוכונדריאליים העיקריים המפורטים והמוסברים באיור 2. - כדי לנתח את הנתונים, ייצא את הנתונים כדוח גיליון אלקטרוני מתוכנת בקר WAVE והשתמש בתבנית גיליון אלקטרוני עצמאית לניתוח.
- כדי לייצא נתונים לאחד ממחוללי הניתוח האוטומטיים, בצע את פקודות ייצוא הנתונים בבקר WAVE ובחר את מחולל הייצוא התואם לסוג הבדיקה. לחלופין, ייצא את קובץ הנתונים והעלה אותו לניתוח סוסון ים.

איור 2: מתארים סכמטיים לפרמטרים הנגזרים מניתוח נתוני שטף חוץ-תאי. קיצור: OCR = שיעור צריכת חמצן. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
כדי להשיג ספרואידים קומפקטיים מעוצבים היטב, כל קו תאים הותאם בנפרד לצפיפות הזריעה ולמשך הטיפוח (איור 3). קווי תאים A549, HepG2/C3A ו-SK-OV-3 יצרו בתחילה אגרגטים רופפים שלא התקדמו לכדורואידים עגולים עם היקפים מוגדרים בבירור עד לאחר 7 ימים בתרבית. לעומת זאת, תאי MCF-7 יכולים ליצור ספרואידים תוך 3 ימים. היה מתאם ברור בין צפיפות הזריעה הראשונית של התאים לבין נפח הספרואידים לאחר תקופת התרבית עבור כל המודלים הספרואידיים. גודל הספרואידים והמורפולוגיה הותאמו לצפיפות הזריעה. המורפולוגיה והמחזוריות החלו לרדת עם הגדלת גודל הספרואידים בכל המודלים. אסטרטגיות זריעה עבור קווי תאים הותאמו ב-4 ×-103 תאים/באר עבור תאי A549 ו-SK-OV-3; תאי HepG2/C3A עברו בעבר אופטימיזציה במקומות אחרים ל-1 × 103 תאים/באר, ותאי MCF-7 שימשו ב-4 × 103 תאים/באר בכל הבדיקות. באסטרטגיות זריעה אופטימליות, נפח הספרואידים היה בין 5.46 × 107 μm3 (SK-OV-3) ו-1.45 × 108 מיקרומטר3 (A549) (איור 3B). לכל סוגי הספרואידים היה מתאם ליניארי בין צפיפות הזריעה הראשונית לבין נפח הספרואידים כאשר ל-A549 ול-HepG2/C3A היו ערכי R2 של 0.957 ו-0.947, בהתאמה. נפחי הספרואידים MCF-7 ו-SK-OV-3 נמצאו שניהם כבעלי מתאם גדול יותר עם צפיפות הזריעה הראשונית, R2=0.977 (איור 3A).
מעגליות ספרואידית חושבה באמצעות פלנימטריית תמונה בתוך תוכנת ניתוח FIJI תוך שימוש בקטרים הספרואידים הארוכים והקצרים. לסימטריה ספרואידית מושלמת הייתה מעגליות = 1.0; סטייה מ-1.0 הצביעה על אובדן מעגליות (איור 3C). מעגליות הייתה גדולה יותר בספרואידים MCF-7 מאשר במודלים אחרים שבהם המעגליות נשמרה בין 0.83 ל-0.9 בכל צפיפויות הזריעה. לשם השוואה, ההיקף החיצוני של הספרואידים SK-OV-3 לא היה כפי שהוגדר בבירור, ונפח הספרואידים היה קטן משמעותית גם לאחר 7 ימים בתרבית, והניב ספרואידים עם מעגליות מקסימלית של 0.61 בצפיפות זריעה של 4 × 103/well. כמו כן נמצא שתאי HepG2/C3A יוצרים ספרואידים הדוקים ומעוצבים היטב עם מורפולוגיה אחידה על פני שטח הפנים של כל הספרואידים, כאשר העגוליות נשמרה על 0.79 עבור תאים שנזרעו ב-1 × 103 תאים/באר. נראה כי תאי A549 עקבו אחר מגמה שבה מעגליות ומורפולוגיה של ספרואידים שופרו עם צפיפות הזריעה; עם זאת, המעגליות לא הייתה גדולה מ-0.63 בצפיפות ששימשה בניסויים אלה.
הנשימה המיטוכונדרית הבסיסית חושבה כ-OCR שנמדדה מספרואידים שנזרעו ב-1 × 103, 2 × 103, 4 × 103, או 8 × 103 תאים/באר במיקרו-לוחות של תרבית ספרואידית חיבור אולטרה-נמוכה (איור 3D). עבור כל סוגי הספרואידים, זיהוי תווים אופטי (OCR) גדל עם הגודל הספרואידי והיה מתואם באופן ליניארי לנפח הספרואידים עם R2 הגבוה ביותר בספרואידים MCF-7 ב-0.988 והנמוך ביותר בספרואידים SK-OV-3 ב-0.744 (איור 3E). ה-OCR שנמדד היה שונה סטטיסטית בין כל קבוצות הניסוי. ל-A549 היה ה-OCR הנמוך ביותר, והוא השיג רק 18 pmol/min/well בגודל הספרואידי הגדול ביותר (איור 3D). לעומת זאת, הספרואידים MCF-7 הניבו זיהוי תווים אופטי (OCR) דומה בגודל הספרואיד הקטן ביותר לאחר 3 ימים בלבד בתרבית, והגיעו ל-OCR בסיסי מרבי של 53 pmol/min/well עבור גודל הספרואיד הגדול ביותר (איור 3D). HepG2/C3A הניבו נתוני זיהוי תווים אופטי (OCR) התואמים מאוד את גודל הספרואידים ואת המורפולוגיה שלהם. בספרואידים HepG2/C3A שנזרעו מ-1 ×-103 תאים/באר, ה-OCR הבסיסי הגיע לממוצע של 15 pmol/min/well, ועלה למקסימום של 52 pmol/min/well בספרואידים הגדולים ביותר (איור 3D). זיהוי תווים אופטי (OCR) בספרואידים SK-OV-3 היה משמעותי רק בין ספרואידים שגדלו מ-4 × 103 תאים/באר ו-8 × 103 תאים/באר, עם הבדל קטן שנראה בספרואידים שגדלו בין 1 ×10 3, 2 × 103, או 4 × 103 תאים. למרות הבדלי הגודל, נתוני ה-OCR היו דומים מאוד בין הספרואידים HepG2/C3A ו-MCF-7 בכל נקודות הגודל. יחסית לגודל הספרואידים (μm3), OCR הבסיסי של כדורי MCF-7 היה דומה לזה של כדוריות HepG2/C3A שגדלו במשך 7 ימים מ-1,000 תאים לכל באר.

איור 3: פרמטרי גדילה כדוריים קובעים את הנשימה המיטוכונדרית הבסיסית. (A) הספרואידים A549, HepG2/C3A, MCF-7 ו-SK-OV-3 הותאמו לצפיפות הזריעה וצמיחתם מנוטרת ב-1 × 103, 2 × 103, 4 × 103 ו-8 × 103 תאים/באר בכל אחד מהפוטומיקרוגרפים, משמאל למעלה ועד ימין למטה, בהתאמה; סרגלי קנה מידה = 500 מיקרומטר. (B) גודל הספרואיד חושב באמצעות נתונים פלנימטריים מפוטומיקרוגרפים שנאספו והושווה באמצעות סטטיסטיקת המתאם של פירסון; קווים מנוקדים מייצגים את ההתפלגות של רווח בר-סמך של 95%. (C) המורפולוגיה הספרואידית הושוותה על ידי חישוב המעגליות. (D) OCR נמדד פי 5, ולאחר מכן נוסף רוטנון-אנטימיצין A כדי להסביר את קצב הנשימה הלא-מיטוכונדריאלי באמצעות אנלייזר Agilent Seahorse XFe96. בסיס OCR נמדד כ- OCR - OCRr/a הושווה בין צפיפות הזריעה (D) לנפח הספרואידי (E). הנתונים הם ממוצעים ±-SEM מ-5-8 שכפולי באר לכל סוג ספרואיד וצפיפות זריעת תאים. קיצורים: OCR = שיעור צריכת חמצן; OCRבסיסי = נשימה מיטוכונדריאלית בסיסית; OCRr/a = OCR לאחר הוספת רוטנון-אנטימיצין A. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
קורס הריכוז והזמן לחשיפה לאפנונים נשימתיים בניתוח XF הוא שלב קריטי באופטימיזציה של הבדיקה. תרכובות אפנן הנשימה - אוליגומיצין, BAM15, תערובת של רוטנון-אנטימיצין A-או בקרת רכב DMSO הוזרקו ברצף דרך יציאות הזרקת מחסנית החיישן לתוך בארות מיקרו-פלטה המכילות את הספרואידים MCF-7 (איור 4A). ארבעה מחזורי מדידה הושלמו כדי לקבוע את ממוצע ה-OCR הבסיסי של 30-40 pmol/min/well עבור כל קבוצות הדגימה. במשך שארית משך הבדיקה, מודולטורים נשימתיים נוספו ברצף כל 5 מחזורי מדידה כדי להשיג ריכוזי באר סופיים של 0.5 מיקרומטרM (0.5 מיקרוגרם/מ"ל אוליגומיצין) לאחר הזרקה 1; 2.0 מיקרומטר (2 מיקרוגרם/מ"ל אוליגומיצין) לאחר הזרקה 2; 5 מיקרומטר (5 מיקרוגרם/מ"ל אוליגומיצין) לאחר הזרקה 3; ולבסוף, ריכוז באר מקסימלי של 11 מיקרומטר (11 מיקרוגרם/מ"ל אוליגומיצין) לאחר התוספת הרציפה הרביעית. הספרואידים MCF-7 לא הגיבו לבקרת הרכב לאורך כל הניסוי (איור 4A). זיהוי תווים אופטי (OCR בסיסי) השתנה מיד לאחר הזריקה הראשונה של כל תרכובת בהתאמה בריכוז הנמוך ביותר של 0.5 מיקרומטר או 0.5 מיקרוגרם/מ"ל אוליגומיצין (איור 4B). זיהוי תווים אופטי (OCR) בספרואידים מסוג MCF-7 הורד עם אוליגומיצין מ-41 pmol/min/well ל-23 pmol/min/well לאחר 5 מחזורי מדידה לאחר הזרקה ראשונה של 0.5 מיקרוגרם/מ"ל (איור 4B).
בתגובה ל-0.5 מיקרומטר BAM15, ה-OCR הוגדל מ-33 ל-41 pmol/min/הרבה לפני הזריקה השנייה (איור 4C). באופן יחסי, השילוב של רוטנון בתוספת אנטימיצין A הוריד את ה-OCR מ-37 ל-13 pmol/min/הרבה לפני הזריקה השנייה (איור 4D). עקבות קינטיים חשפו עוד ירידה ליניארית מתמדת (אוליגומיצין ורוטנון-אנטימיצין A) או עלייה (BAM15) ב-OCR. עבור כל משטרי המינון המורכבים, OCR במצב יציב הושג תוך 10-12 מחזורי מדידה מלאים (60-72 דקות) בריכוז כולל של 2 μM BAM15, 2 μM רוטנון, 2 μM אנטימיצין A ו-2 מיקרוגרם/מ"ל אוליגומיצין (איור 4A). קצב צריכת החמצן הגיע לרמה של כ-19 פמול/דקה/באר (אוליגומיצין), 52 פמול/דקה/באר (BAM15) ו-10 pmol/min/well (רוטנון-אנטימיצין A) (איור 4A). להגברת הריכוז המורכב של אוליגומיצין, BAM15 או רוטנון בתוספת אנטימיצין A לא הייתה השפעה ברורה על זיהוי תווים אופטי (OCR), שנשאר קבוע לאורך כל שארית הבדיקה. נתונים אלה מראים כי יש לקחת בחשבון הן את ריכוז התרכובות והן את מהלך הזמן של החשיפה לתרכובות אפנן הנשימה לצורך אופטימיזציה של הבדיקה בעת שימוש בספרואידים תלת-ממדיים.

איור 4: טיטרציה של תרכובות אפנן הנשימה כצעד חשוב לאופטימיזציה של ניתוח שטף חוץ-תאי. (A) כדוריות MCF-7 נזרעו ב-4 × 103 תאים/בארות ותרביתם במשך 3 ימים לפני שהונחו בבארות של מיקרו-פלטת בדיקה ספרואידית המכילה XF RPMI ונחקרה עבור מודולטורים מיטוכונדריאליים ± OCR באמצעות מנתח XFe96. זיהוי תווים אופטי (OCR) נמדד פי 5, ולאחר מכן נוספו טיטרציות של בקרת רכב, אוליגומיצין (B), BAM15 (C) או רוטנון-אנטימיצין A (D) כדי לעכב את הסינתאז ATP המיטוכונדריאלי, לקבוע את יכולת הנשימה המרבית או לקבוע קצב נשימה לא מיטוכונדריאלי, בהתאמה. הריכוז של כל אפנן מיטוכונדריאלי הוגדל על פני 4 אסטרטגיות הזרקת טיטרציה בודדות (0.5 μM, 1.5 μM, 3 μM ו-6 μM; היחידות עבור אוליגומיצין הן מיקרוגרם/מ"ל) כדי לקבוע OCR מקסימלי במצב יציב בתגובה לריכוז תרכובת אופטימלי. זיהוי תווים אופטי (OCR) נמדד עבור 5 מחזורי מדידה בין כל הזרקה. הנתונים הם ממוצעים ± SEM מ 5-8 משוכפלים היטב בודדים. קיצור: OCR = שיעור צריכת חמצן. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
אחד היתרונות העיקריים של טכנולוגיית XF הוא היכולת לחקור את תפקוד המיטוכונדריה בתאים ורקמות שלמות. כדי לבחון היבטים ספציפיים של תפקוד המיטוכונדריה בתאים וברקמות, מווספנים מיטוכונדריאליים מתווספים ברצף לבארות של המיקרו-לוחית של הדגימה דרך 4 יציאות ההזרקה הזמינות במחסנית החיישן. הרצף הטיפוסי של מודולטורים המשמשים לבדיקת פרמטרים מיטוכונדריאליים במבחני XF הם אוליגומיצין, פרוטונופור (למשל, FCCP או BAM15), ושילוב של רוטנון בתוספת אנטימיצין A, אשר מתווספים ברצף כדי לעכב את הסינתאז ATP המיטוכונדריאלי, לקבוע את יכולת הנשימה המקסימלית, ונכונים לקצב הנשימה הלא-מיטוכונדריאלי, בהתאמה. רצף טיפוסי זה של תוספות אפנן מכונה מבחן MitoStress על ידי יצרן טכנולוגיית הבדיקה. בהתחשב בכך שאוליגומיצין יכול לעכב נשימה לא מגורה על-ידי לכידה בחלק מהחד-שכבתיים של התאים20, בחנו זאת עם ספרואידים תלת-ממדיים שמקורם בסרטן על-ידי מדידת OCR (OCRmax) מגורה-לא-מגורה(OCR) לפני (יחיד) ואחרי הזרקת אוליגומיצין (רציפה) (איור 5A-D). זיהויתווים אופטי (OCR max) לא הוגבל באופן משמעותי על-ידי הוספת אוליגומיצין בספרואידים שנוצרו מ-HEPG2/C3A או מ-SK-OV-3 (איור 5E ואיור 5G). עם זאת, מספר ה-OCRהמרבי הופחת באופן משמעותי בספרואידים A549 ו-MCF-7 לאחר הזרקה רציפה של BAM15 לאחר אוליגומיצין בהשוואה ל-OCRmax שהושגה מהזרקה בודדת של BAM15 (איור 5F ואיור 5H). אלא אם כן ידוע אחרת, מומלץ אפוא להשתמש בבארות נפרדות לטיפול עם אוליגומיצין ו uncoupler, עם תוספת סופית של רוטנון ואנטימיצין A בעת חקירת חילוף החומרים של אנרגיה מיטוכונדריאלית של ספרואידים 3D. גישה זו עדיין מאפשרת חישוב של כל הפרמטרים המיטוכונדריים כמו במבחן MitoStress טיפוסי שבו תרכובות מתווספות ברצף.

איור 5: הזרקה בודדת או רציפה של תרכובות נשימה מיטוכונדריאליות. ספרואידים שמקורם בתאי סרטן של MCF-7, HEPG2/C3A, SK-OV-3 ו-A549 הוכנסו לבארות של מיקרו-לוחית XFe96 ספרואידית ב-XF RPMI ונבדקו לאיתור OCR באמצעות מנתח סוסון הים האג'ילנט XFe96. זיהוי תווים אופטי (OCR) נמדד פי 5, ולאחר מכן 2 μg/mL אוליגומיצין (הזרקה יציאה A: עקבות ירוקים) או 5 μM BAM15 (הזרקה יציאה A: עקבות כחולים או יציאת הזרקה B: עקבות ירוקים) כדי לעכב את ה-ATP סינתאז המיטוכונדריאלי ולקבוע את יכולת הנשימה המקסימלית, בהתאמה. נתוני זיהוי תווים אופטי (OCR) קינטיים מבוטאים כ- % בסיסי (A-D). קיבולת הנשימה המקסימלית (OCRmax) חושבה כגורם של OCR בסיסי על ידי המשוואה: OCRmax = OCRBAM15 / OCRבסיסי. OCRמקסימום התקבל מממוצעי OCR על פני מחזורי מדידה 8-10 לאחר הזרקת BAM15 עם (פסים ירוקים) וללא (פסים כחולים) אוליגומיצין. הנתונים הם ממוצעים ± SEM מ 3-8 בודדים משוכפלים היטב על פני מיקרו-לוחית הבדיקה הספרואידית. קיצורים: OCR = שיעור צריכת חמצן. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
תוך שימוש בצפיפויות אופטימליות של זריעת תאים, ריכוזי תרכובות, אסטרטגיית הזרקה ותקופת מחזור מדידה שנקבעו בניסויי אופטימיזציה אלה (טבלה 3), פיתחנו פרוטוקול מפורט לבדיקה מדויקת של הנשימה המיטוכונדרית הבסיסית:OCR בסיסי (איור 6A), נשימת זרחון ADP: OCRADP (איור 6B), נשימת דליפה: OCRomy (איור 6C), יעילות צימוד (איור 6D ), קיבולת נשימתית מקסימלית: OCRmax (איור 6E), וקיבולת נשימה רזרבית:OCR רזרבי (איור 6F) באמצעות ספרואידים תלת-ממדיים שמקורם בסרטן.

איור 6: בדיקת זיהוי תווים אופטי (OCR) עם טכנולוגיית XF כדי לבסס את חילוף החומרים של אנרגיה מיטוכונדריאלית של ספרואידים שמקורם בסרטן. ספרואידים שמקורם בתאי סרטן של MCF-7, HEPG2/C3A, SK-OV-3 ו-A549 הונחו בבארות של מיקרו-לוחית בדיקה ספרואידית ב-XF RPMI ונבדקו לאיתור זיהוי תווים אופטי (OCR) באמצעות מנתח סוסון הים האג'ילנט XFe96. זיהוי תווים אופטי (OCR) נמדד פי 5, ולאחר מכן נוספו 2 μg/mL אוליגומיצין, או 5 μM BAM15, ו-RA כדי לעכב את הסינתאז ATP המיטוכונדריאלי, לקבוע את יכולת הנשימה המקסימלית ולחשב את קצב הנשימה הלא-מיטוכונדריאלי, בהתאמה. (A) הנשימה המיטוכונדריאלית הבסיסית (OCRbasal) חושבה כממוצע של זיהוי תווים אופטי (OCR) מ-3 מחזורי המדידה שלפני הזרקת יציאה A. (B) יעילות הצימוד של זרחון חמצוני הוערך על ידי ביטוי OCRADP (OCR בסיסי -דליפת OCR) כאחוזמבסיס OCR. (C) נשימת הזרחון של ADP (OCRADP) נמדדה כ-OCR רגיש לאוליגומיצין, שחושב מה-OCR הממוצע על פני מחזורי מדידה 11-13 לפני הזרקת BAM15. (D) נשימתאומי דליפה (דליפת OCR) נמדדה כחוסר רגישות OCR לאוליגומיצין, המחושב מממוצע ה-OCR הממוצע על פני מחזורי מדידה 11-13. (E) קיבולת הנשימה המרבית (OCRmax) נמדדה כממוצע ה-OCR המרבי שנמדד לאחר הזרקת BAM15. (F) קיבולת הנשימה הרזרבית חושבה על ידי הבעתרזרבי OCR (OCRמקסימום - OCRבסיסי) כאחוז מבסיס OCR. OCR לאחר הזרקת רוטנון-אנטימיצין A (OCRr/a) הופחתה מכל השיעורים כדי לתקן עבור OCR שאינו מיטוכונדריאלי. הנתונים הם ממוצעים ±-SEM מ-3-8 משוכפלים היטב בודדים על פני הלוח הספרואידי XFe96. קיצורים: OCR = שיעור צריכת חמצן; RA = 2 μM rotenone-2 μM antimycin A. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
כדורי MCF-7 שגדלו מ-4 × 103 תאים/הרבה יותר מ-3 ימים שימשו כמודל לקביעת העברה, מיקום וניתוח אופטימליים בתוך מיקרו-לוחות בדיקה כדוריים. באמצעות ממדים שסופקו עבור המיקרו-פלטה הספרואידית מהיצרן, משטח הבאר פוצל לשלושה אזורי-אזור למיקום ספרואידי אופטימלי (איור 7A), כאשר אזור 1 הודגש כאזור האופטימלי במרכז הבאר. בעזרת צנרת זהירה באמצעות קצות פיפטה רחבים, הועברו הספרואידים ללוחות הספרואידים והתפזרו באופן אקראי על פני משטחי הבאר על ידי אלוטיון הכבידה (איור 7B). כאשר הספרואידים הועברו בזהירות באמצעות אלוטיון כוח הכבידה, ניתן היה למצוא את רוב הספרואידים בדרך כלל באזורים 1-2 של המיקרו-לוחית, תוך שימוש בטכניקות ההעברה המומלצות מהיצרן. במקום שבו הספרואידים הוצאו מהקצה של הפיפטה על ידי שאיפה, הספרואידים הוצבו לעתים קרובות מעבר לאזורים אלה ולא ניתן היה לראותם באמצעות מיקרוסקופיה.
כדי להשוות את מיקומי המיקום של הספרואידים, כדוריות MCF-7 הועברו למיקרו-לוחות הבדיקה של הספרואידים באזורים המיועדים 1-3 או מחוץ לאזור (איור 7A). 4 הבארות האלה היו במעקב באמצעות ניסוי קינטי OCR בנקודת ההתחלה ולאחר הוספת אוליגומיצין, BAM15 או רוטנון-אנטימיצין A (איור 7C). זיהוי תווים אופטי (OCR) חושב מממוצע של שלוש קריאות מחזוריות לפני כל זריקה (איור 7B). זיהוי תווים אופטי (OCR) נמדד באופן קינטי במשך 200 דקות ב-4 הבארות שנבחרו (איור 7C) ובתיקון הבסיס (איור 7D). במקומות שבהם הספרואידים הוצבו באזור 3 או מחוץ לאזור, ה-OCRs הבסיסיים היו נמוכים משמעותית מהספרואידים שהוצבו באזורים 1 ו-2 (איור 7C). גם ההשפעות של תרכובות הנשימה אוליגומיצין, BAM15 ורוטנון-אנטימיצין A היו שונות באופן דרמטי בין ספרואידים הממוקמים באזורים 1 ו-2 בהשוואה לאזור 3 ולאזורים מחוץ לאזור. עלייה ב-OCR נצפתה עם אוליגומיצין בספרואידים הממוקמים באזור 3 או מחוץ לאזור (איור 7E). יתר על כן, ספרואידים שהוצבו באזור 3 או מחוץ לאזור חוו תגובה גבוהה מדי ל-BAM15 עם זיהוי תווים אופטי גבוה יותר מהבסיס לאחר הזרקת רוטנון-אנטימיצין A (איור 7E). למרות עלייה של כמעט פי שניים ב-OCR הבסיסי (איור 7C) עם ספרואידים הממוקמים באזור 2 לעומת אזור 1, השינויים המתקפלים בתגובה לכל תרכובות הנשימה היו דומים מאוד (איור 7E), מה שמרמז על כך שהבדלים ב-OCR הבסיסי בין ספרואידים הממוקמים באזורים 1 או 2 לא צפויים להיות תוצאה של מיקום בתוך הבאר.

איור 7: מיקום של ספרואידים בתוך המיקרו-פלטה של הבדיקה הספרואידית מכתיב אפקטים של OCR בסיסי ומודולטור מיטוכונדריאלי באמצעות טכנולוגיית XF. הספרואידים MCF-7 נזרעו ב-4 × 103 תאים/באר ותרבית במשך 3 ימים לפני שהוכנסו לבארות של המיקרו-פלטה הספרואידית המכילה XF RPMI ונבדקו עבור OCR ± מודולטורים מיטוכונדריאליים באמצעות מנתח סוסיון הים האג'ילנט XFe96. (A) פוטומיקרוגרפים של מיקומי אזור ספרואידים במיקרו-לוחות בדיקה כדוריים לאחר משך הבדיקה; סרגל קנה מידה = 500 מיקרומטר ו- OCR שנלכדו מבארות מקבילות לאורך זמן המבוטאות כ- pmol / min-1 / well-1 (B) או % בסיסי (C). (D) אפקטים של אפנן מיטוכונדריאלי של כדוריות MCF-7 הממוקמות באזורים שונים בתוך מיקרו-לוחית הבדיקה הספרואידית; נתונים המבוטאים כשינוי קיפול מבזלת. (E) דוגמה למעקב קינטי המדגיש אילו מדידות נתוני OCR (עיגולים אדומים) משמשות לחישוב התגובה של כל אפנן מיטוכונדריאלי עבור נתונים המוצגים ב- E. הנתונים המוצגים הם מתגובות טובות בודדות. קיצור: OCR = שיעור צריכת חמצן. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
קריטריוני הבחירה לרקע הם בעלי חשיבות גבוהה; השימוש בבארות החיצוניות ביותר לתיקון רקע אינו מייצג את כל בארות המיקרו-פלטה, מה שעלול להוביל להסקת הנחות נתונים שגויות ולמסקנות נתונים שגויות עקב השפעות קצה על פני המיקרו-לוחית הספרואידית. כדי להעריך תצפית זו, נעשה שימוש בספרואידים MCF-7 כדי להשוות את הליכי תיקון הבדיקה כדי לגזור ערכי OCR בתגובה להוספת בקרת רכב, אוליגומיצין, BAM15 או רוטנון-אנטימיצין A (איור 8). כל התרכובות הנשימתיות הניבו את פרופילי ה-OCR הקינטיים הצפויים עבור התרכובות שנבחרו, וחשפו קצב נשימה בסיסי קבוע ממוצע של 20-30 pmol/min/well (איור 8A). עם זאת, כאשר נתוני הבדיקה נותחו באמצעות הבארות החיצוניות ביותר לתיקון טמפרטורת הרקע (A1, A12, H1 ו- H12), הערכים שנחשפו עבור OCR לאחר הוספת תרכובות נשימה היו נמוכים במיוחד; זיהוי תווים אופטי (OCR) הניב ערכים שליליים עבור רוטנון-אנטימיצין A. בתגובה לתצפיות אלה, בוצע ניתוח חלופי באמצעות סדרה של בארות ריקות, המפוזרות באופן אקראי על פני המיקרו-לוחית הספרואידית, כבארות תיקון טמפרטורה ברקע (איור 8B). כאשר הוחל תיקון רקע חלופי, כל ההשפעות המורכבות היחסיות על זיהוי תווים אופטי (OCR) היו זהות בשתי קבוצות הניתוח; עם זאת, ערכי זיהוי תווים אופטי (OCR) מוחלטים עלו בכ-10 pmol/min/well (איור 8). נתונים אלה מדגישים את העוצמה והחשיבות של תיקון טמפרטורת הרקע במיקרו-לוחות בדיקה של ספרואידים ומדגישים את החשיבות של אופטימיזציה של המשתמש לניתוח XF.

איור 8: בחירה אקראית של בארות לתיקון רקע כדי לשפר את הבקרה על שיפועי טמפרטורה על פני המיקרו-לוחית של הבדיקה הספרואידית. נתוני זיהוי תווים אופטי (OCR) שעברו אקסטרפולציה מאיור 2A באמצעות בארות מומלצות לתיקון רקע (A) לעומת בארות שהוקצו באופן אקראי לתיקון רקע (B). קיצור: OCR = שיעור צריכת חמצן. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
שלא כמו מונו-שכבות של תאים, כדוריות מייצגות צבירה הטרוגנית של תאים במרחב תלת-ממדי ולכן דורשות התייחסות מעמיקה ביחס לניתוח, במיוחד בעת נרמול נתונים אלה. מאמר זה מציג שלוש גישות לנרמול נתוני XF שנרכשו מספרואידים מסוג MCF-7 (איור 9). כאשר הוא אינו נורמלי, OCR מתואם באופן חיובי (R2 = 0.98) עם גודל ספרואידי (נקבע על ידי צפיפות זריעת תאים ראשונית) באופן משמעותי כאשר משווים אותו סטטיסטית עם מקדם המתאם של פירסון, P = 0.0057 (איור 9A). הקשר הליניארי הזה מורד כאשר OCR מנורמל לצפיפות הזריעה הראשונית של התאים (R2 = 0.78) וכבר אינו מתואם באופן משמעותי עם גודל הספרואידים (P = 0.117, איור 9B). זה גם המקרה כאשר מנורמל לנפח ספרואידי (R2 = 0.77; מקדם המתאם של פירסון P = 0.120, איור 9C) ותכולת dsDNA גרעינית (R2 = 0.58; מקדם המתאם של פירסון P= 0.233, איור 9D). נתונים אלה מדגישים את החשיבות של נרמול נתוני XF בעת בדיקת חילוף החומרים המיטוכונדריאלי של ספרואידים, במיוחד אם הם בגדלים שונים.

איור 9: נורמליזציה של נתוני שטף חוץ-תאיים שנרכשו מספרואידים תאיים. (A) נתוני OCR גולמיים התקבלו מ-MCF-7 בתרבית במשך 3 ימים והותוו באמצעות המודל של פירסון כדי להשיג מקדם מתאם בין צפיפות זריעה כדורית לבין זיהוי תווים אופטי (OCR) ערך P מוגדר ב- 0.05. (B) נתוני זיהוי תווים אופטיים גולמיים נורמלו כנגד צפיפות הזריעה הראשונית של הספרואידים; (C) נפח ספרואיד MCF-7 המתקבל מפלנטימטריה של מיקרוסקופיה; ו-(D) תכולת דנ"א של ds גרעינית בהשוואה באמצעות מקדם המתאם של פירסון. קיצורים: OCR = שיעור צריכת חמצן; ds DNA = דנ"א דו-גדילי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
| קו סלולרי | צפיפות זריעה (טוב) | גדילה כדורית (ימים) | נפח ספרואידי סופי (μM3) | זיהוי תווים אופטי (pmolO2 לדקה/באר) | רגישות למפגש עם זיהוי תווים אופטי (כן/לא) |
| סקוב | 1000 | 5 | 9.52E+06 | 28 ± 3.5 | כן |
| סקוב | 2000 | 5 | 2.38E+07 | 26 ± 1.4 | כן |
| סקוב | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | כן |
| סקוב | 8000 | 5 | 1.11E+08 | 50 ± 7.9 | כן |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0.7 | לא |
| HepG2 | 2000 | 5 | 2.88E+07 | 23 ± 1.8 | כן |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1.7 | כן |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2.8 | כן |
| A549 | 1000 | 5 | 2.11E+07 | 30 ± 2.5 | כן |
| A549 | 2000 | 5 | 3.57E+07 | 41 ± 1.6 | כן |
| A549 | 4000 | 5 | 6.93E+07 | 53 ± 7.2 | כן |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8.4 | כן |
| MCF-7 | 1000 | 3 | 1.60E+07 | 29 ± 0.8 | כן |
| MCF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1.7 | כן |
| MCF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1.7 | כן |
| MCF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2.9 | כן |
טבלה 4: פרמטרים ממוטבים לקביעת מדידות זיהוי תווים אופטי (OCR) בסיסי בספרואידים תלת-ממדיים בודדים. קיצור: OCR = שיעור צריכת חמצן.
קובץ משלים 1: ניתוח של גודל ונפח ספרואידים. אנא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 2: כימות של דנ"א דו-גדילי מספרואידים במיקרו-לוחית הספרואידית. אנא לחץ כאן כדי להוריד קובץ זה.
קובץ משלים 3: המלצות למספר המשכפלים הנדרשים כדי להשיג מערכי נתונים אמינים של בדיקת XF. אנא לחץ כאן כדי להוריד קובץ זה.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
ממצאים ותפוקות עיקריים
מאמר זה מספק פרוטוקול מפורט לבדיקת חילוף החומרים של אנרגיה מיטוכונדריאלית של ספרואידים תלת-ממדיים בודדים באמצעות סדרה של קווי תאים שמקורם בסרטן עם מנתח XFe96 XF. שיטה מפותחת ומתוארת לטיפוח מהיר של A549, HepG2/C3A, MCF7 ו-SK-OV-3 כדוריים תאיים באמצעות טכנולוגיות דוחות תאים לצבירה כפויה. פרוטוקול זה נותן מענה לשיקולים רבים של בדיקת חילוף החומרים של הספרואידים באמצעות טכנולוגיית XF, כולל (1) אופטימיזציה של פרוטוקולי תרביות ספרואידים וטיפול והעברה של ספרואידים למיקרו-לוחות בדיקה ספרואידיים ספציפיים מיצרן הטכנולוגיה מכלי התרבות המקוריים שלהם; (2) ריכוז התרכובות הנשימתיות שיש להשתמש בהן והתלות בזמן של חדירת התרכובות; (3) אסטרטגיות הזרקה לשימוש; ו-(4) דרכים לנרמל נתונים בין קבוצות ניסוי. כל השיקולים הללו נבחנו במאמר הנוכחי ונדונים ביתר פירוט בהמשך. שיטות אלה מוצגות כגישות פשוטות ליצירת נתוני שטף חמצן מטבוליים עקביים באמצעות ספרואידים תלת-ממדיים בודדים עם מנתח השטף XFe96. גישה ניסיונית זו יכולה לשמש כנקודת מוצא ורובריקה לשימוש במודלים ספרואידיים אחרים המיושמים בקלות במסגרת מעבדה בסיסית.
שיקולים
צמיחה, גודל ורגישות כדורית של טכנולוגיית XF
כדי ליצור נתונים הניתנים לשחזור עם טכנולוגיית XF, חיוני לאפיין ולמטב את הבדיקה עבור המודל הספציפי. גישה זו היא פשוטה יחסית במונולייר בסיסי של תאים; עם זאת, זה מציב אתגרים נוספים בעת טיפוח תאים כספרואידים תלת-ממדיים. במהלך הניסויים שהוצגו כאן, מדיום RPMI של היצרן הושלם עם השימוש. בעוד שצוין כי כמה קווי תאים, כלומר HepG2/C3A, היו תרבית במדיום גדילה DMEM, במהלך בדיקות קצרות יחסית אלה (~ 3-5 שעות), החלפה עם נוסחאות RPMI-DMEM הייתה השפעה מוגבלת על ניתוח XF. הניסוח של שתי המדיות דומה מאוד, ומשתמשים יכולים 'לכוונן' את מדיית ה-RPMI של סוסון הים כך שתתאים למטריצה של מדיומים של תרביות התאים שלהם באמצעות תוסף תזונה, למשל, גלוקוז מוגבר, תוספת נוספת של מקורות פחמימות. קריטי לניסוח הסופי של כל מאגרי ה-XF והמדומים הוא היעדר פנול-אדום, מה שעלול להפריע לבדיקות הפלואורסצנטיות בתוך צלחת מחסנית הגשושית XF, ונתרן ביקרבונט, שיוביל לאלקליניות בשל היעדר אגירת CO2 הקיימת באינקובטורים של תרביות תאים. ניתן לרכוש ו/או ליצור מדיה ומאגרים אחרים בתוך החברה. לדוגמה, מאגר HEPES של Krebs Ringer הוא מאגר פשוט שניתן להשתמש בו כדי להעריך נשימה בתאים רבים ושונים, כולל מודלים ספרואידיים. עם זאת, משתמשים במבחני XF צריכים לשים לב ששינוי במדיום/מאגר וההשלמה שלו עשויים לשנות את יכולת האגירה הכוללת שלו. זה מדאיג במיוחד כאשר משתמשים עשויים להיות מעוניינים למדוד ECAR, שבו יש להעריך את גורם המאגר של המדיום כדי לאפשר טרנספורמציה של ECAR לקצב שטף הפרוטונים (PER).
מכיוון ש-OCR תאי שנמדד על ידי טכנולוגיית XF פרופורציונלי לצפיפות התא כאשר מספר התאים בבאר נמצא ברגישות של המערכת, היה חשוב לחקור את הקשר הזה באמצעות ספרואידים תלת-ממדיים בודדים. על-ידי בדיקת OCR של ספרואידים תלת-ממדיים בודדים בתרבית מ-4 קווי תאים סרטניים שונים שנזרעו בצפיפויות של 1,000, 2,000, 4,000 או 8,000 תאים לבאר, אנו מראים שמנתח XFe96 רגיש מספיק כדי לקלוט שינויים בקצב הנשימה המיטוכונדרית בין ספרואידים תלת-ממדיים הגדלים מצפיפות זריעת תאים שונות (איור 3). אנו מראים שהטווח האופטימלי של צפיפות זריעת התאים, ולכן נפח הספרואידים ליצירת ספרואידים תלת-ממדיים לבדיקת זיהוי תווים אופטי (OCR), משתנים בהתאם לסוג התא. זה מוצג על ידי הקשר הליניארי בין זיהוי תווים אופטי (OCR) לבין צפיפות הזריעה או נפח הספרואידים (איור 3). עבור תאי A549 ו-HepG2/C3A, צפיפות הזריעה האופטימלית לרגישות ל-OCR הייתה בין 1,000 ל-8,000 תאים/באר; זה היה 2,000-8,000 תאים / באר עבור MCF-7 ו 4,000-8,000 תאים / באר עבור תאי SK-OV-3. נתונים אלה מראים כי אופטימיזציה של גודל ספרואידים היא בעלת חשיבות מיוחדת בעת הערכת זיהוי תווים אופטי (OCR) באמצעות טכנולוגיית XF.
שיקולים על נפחים כדוריים מינימליים ומקסימליים ו- OCR בסיסי
באופן כללי, תמיד יהיו ספי מינימום ומקסימום עבור פרמטרים הניתנים למדידה של זיהוי תווים אופטי (OCR) המומלצים על ידי היצרן עבור ניסויים אלה. עבור מנתח XFe96, OCR בסיסי בין 20 pmol O2/min/well ו-200 pmol O2/min/well הם הגבולות התחתונים והעליונים, בהתאמה. זה המקרה עם תאים חד-שכבתיים וספרואידים, והמקום שבו המודל הניסיוני יושב בטווח ה-OCR הדינמי הזה יהיה תלוי בכמות החומר הביולוגי הזמין, למשל, מספר התאים כחד-שכבתיים או בגודל של ספרואידים. ראה טבלה 4 לקבלת דוגמה לאופן שבו הושגו ערכי סף של זיהוי תווים אופטי (OCR) על-ידי המודלים הספרואידיים שבהם נעשה שימוש כאן. ייתכן שיהיה זה נבון לבדוק את רמת החמצן בתוך הבאר שעבורה נתונים אלה זמינים גם ממדידות אלה כנתוני הרמה. יש לראות זאת באופן שגרתי מכל ניסוי למטרות בקרת איכות. אם יש דלדול חמצן בבאר, זה יתבהר בתוך הנתונים. אם זה המקרה, ייתכן שיהיה צורך בהתאמת מחזורי המדידה בתוך הניסוי; לדוגמה, הגדלת שלב הערבוב כך שרמת החמצן בבאר תתאושש לפני תקופת המדידה הבאה במחזור המדידה. למרות שהדבר אפשרי, מצאנו שזה מאוד לא סביר עבור ניסויים חד-כדוריים באמצעות קווי התאים המתוארים.
בחירה של uncouplers מיטוכונדריאליים עבור מבחני שטף חוץ-תאיים
יונופורים פרוטונים, כגון קרבוניל ציאניד 4-(טריפלואורומתוקסי) פניל-הידרזון (FCCP)21, קרבוניל ציאניד m-כלורופניל הידרזון (CCCP)22 או BAM1523, הם כימיקלים חזקים בעלי מולקולות קטנות המסוגלים לשבש את שיפוע הפרוטונים האלקטרוכימיים על פני ממברנות מיטוכונדריאליות, לעכב את ייצור ה-ATP, ובסופו של דבר לבטל את הנשימה המיטוכונדרית24 . מולקולות קטנות חדשות ממשיכות להיות מפותחות למטרות אלה, במיוחד בטיפול במחלות מטבוליות 25,26,27; עיין בשתי חוות דעת מצוינות28,29. לעומת זאת, אי-שיתוף פעולה של נשימה חמצונית נקשר לרעילות לא רצויה מחוץ למטרה30. עם זאת, בתוך מבחנים תאיים במבחנה, המולקולה FCCP עושה דה-פולריזציה של פוטנציאל הממברנה המיטוכונדרית ומפעילה אפקטים מחוץ למטרה כגון דה-פולריזציה של קרום הפלזמה, תוך הפרעה לשטף היונים של NA+ 31; הפרעה בעיבוד חלבונים תאיים32, ואף גרימת סנסנציה תאית33. BAM15 הוצג במקור בשנת 2013 כ-uncoupler מיטוכונדריאלי עם השפעה מינימלית על ממברנות פלזמה23, עם פעילות פרוטונופורית בטווח המיקרומולרי בתאים שלמים וטווח ננומולרי במיטוכונדריה מבודדת 23,34.
בהתחשב בעוצמה של FCCP בדה-פולריזציה של קרום הפלזמה, BAM15 הוא פרוטונופור אמין יותר לנטרול נשימה בתאים שלמים שלמים שלמים במבחני שטף חוץ-תאיים. למרות ש-FCCP ומקבילתו, CCCP, שימשו במשך למעלה מ-50 שנה לבדיקת יכולות נשימה מקסימליות וממשיכים להיות בשימוש נרחב במחקרי XF, השימוש במולקולות קטנות אלה ממעיט לעתים קרובות ביכולת המטבולית המיטוכונדרית והתאית. זה קשור בחלקו מדוע כל כך הרבה פרסומים המשתמשים בטכנולוגיית XF נופלים למלכודת של דיווח על יכולות נשימה רזרביות שליליות או לזלזל ביכולות הנשימה המיטוכונדריאליות האמיתיות כאשר נעשה שימוש ב- FCCP. העוצמה הנוספת של FCCP בתאים וברקמות שלמים מובילה לעתים קרובות לתפקוד מיטוכונדריאלי פגום, ותאים יכולים להתקשות לפעול כראוי כדי לשמור על קיבולת נשימה מקסימלית על פני מחזורי מדידה מרובים לאחר הוספתם, אפילו בריכוזים נמוכים מאוד35. לכן, ניתן למצוא את התגובה של תאים ל-FCCP במחקרים רבים כדי לרדת לאחר תקופת מחזור המדידה הראשונית. בעוד ש- FCCP שימש באופן שגרתי לניתוח XF, BAM15 משמש באופן מועדף במקרים המערבים תאים שלמים או מודלים ספרואידיים, בהתחשב בכך שהוא יכול לשמור על יכולת נשימה מקסימלית במיטוכונדריה דה-קוטבית מלאה בריכוזים של עד 10 μM 3. יתר על כן, BAM15 גורם להשפעות על החמצה חוץ-תאית, אשר עולה בקנה אחד עם זה של חמצון חומרים מזינים באמצעות הידרציה של CO2 כדי ליצור HCO3- ו- H+ במידה רבה יותר מאשר FCCP3. אף על פי כן, במקרה של מיטוכונדריה מבודדת ותאים מחלחלים, כל אחד מהתאים האלה צריך לתפקד כמו גם BAM15 עבור מיטוכונדריאליים ללא שיתוף פעולה אם הוא מצטמצם בריכוז הנכון.
קינטיקה של חדירת תרכובת ורכיבה על אופניים
הריכוזים, החדירה והפרופילים הקינטיים של תרכובות כימיות המשמשות לביצוע בדיקת MitoStress טיפוסית עם ספרואידים תלת-ממדיים באמצעות מנתח XF מורכבים יותר לטיפול. בהתחשב בכך שספרואידים מציגים מבנה תלת-ממדי, חדירת המולקולות לרוחב קוטר הספרואיד היא תהליך מורכב לאין שיעור יותר מאשר על פני מונו-שכבות של תאים. לדוגמה, החדירה הקינטית, ולכן, הרגישות לסוראפניב הכימותרפי נקבעה על ידי גיל הספרואידים, ולכן, הגודל במודל ספרואידי HepG236. היכולת של כימיקלים בעלי מולקולות קטנות (למשל, תרופות, ננו-חלקיקים) להגיע למטרה ביולוגית תלויה במספר גורמים בסיסיים, כולל המורכבות של המערכת שיש לחדור אליה באופן דינמי ולפזראותה דרך 37,38. זה נכון במיוחד לגבי תרופות המכוונות לרקמת הגידול39. בדומה למיקוד הגידול בהקשר של ספרואיד תלת-ממדי, גודל, קומפקטיות ותגובות פנוטיפיות אחרות כגון ביטוי של חלבוני טרנספורטר תרופתיים יכולים לשלוט בזמן החדירה ובריכוז של תרכובת הנדרשת כדי לעורר תגובה ביולוגית.
בפרוטוקול זה, התייחסנו לסוגיה סביב זמן החדירה וריכוז המולקולות הקטנות בתגובה למעכבי ה-ATP סינתאז אוליגומיצין, הפרוטונופור והמיטוכונדריאלי BAM15, והשילוב של מעכבי קומפלקס I וקומפלקס III רוטנון ואנטימיצין A. על-ידי בדיקת ה-OCR של כדוריות MCF-7 בודדות שנחשפו לטיטרציות מרובות של תרכובות הנשימה הנפוצות האלה, אנו מראים שהריכוז האופטימלי של כל תרכובת הנדרשת כדי לגרום לקצב נשימה במצב יציב נמצא בטווח דומה לזה של תאים חד-שכבתיים (איור 4). חשוב לציין, ובשונה ממקביליהם החד-שכבתיים, הוכח כי הגדלת מספר מחזורי המדידה בין הזריקות היא המפתח להשגת זיהוי תווים אופטי (OCR) במצב יציב בספרואידים תלת-ממדיים בודדים. נתונים אלה מדגישים את החשיבות של חדירת תרכובות ואת הפרופילים הקינטיים המתאימים להן כאשר בוחנים פרמטרים נשימתיים מיטוכונדריאליים של ספרואידים תלת-ממדיים באמצעות גישות אלה. באמצעות תכונות אופטימיזציה של ספרואידים, ריכוזים של תרכובות וזמני מחזור מדידה המבוססים על הנתונים שהוצגו באיור 3, איור 4 ואיור 5, נקבעה בדיקת MitoStress מאומתת לבדיקת פרמטרים ספציפיים של חילוף חומרים חמצוני מיטוכונדריאלי במגוון של ספרואידים תלת-ממדיים שמקורם בסרטן (איור 6). חשיבותה, וכמו כמה שורות של תאים סרטניים חד-שכבתיים40, היכולת הנשימתית המקסימלית (קצב הנשימה הלא מגורה-מגורה) של ספרואידים תלת-ממדיים מסוימים שמקורם בסרטן עוכבה על-ידי אוליגומיצין (איור 5). באופן ספציפי, ספרואידים תלת-ממדיים שגדלו מתאי A549 או MCF-7 הראו קצב נשימה מקסימלי נמוך משמעותית כאשר הם לא חוברו ל-BAM15 לאחר הזרקת אוליגומיצין, בהשוואה לכך שהם לא נלכדו על-ידי BAM15 ללא אוליגומיצין (איור 5F ואיור 5H). בהתחשב בכך שהשפעה זו עשויה להיות נוכחת בתרביות ספרואידיות תלת-ממדיות אחרות, אנו מציעים שאם לא נעשה שימוש בפרוטוקול שאומת בעבר, יש להעריך את יכולת הנשימה המרבית בספרואידים תלת-ממדיים ללא אוליגומיצין.
איסוף סימולטני של נתוני ECAR כמדד לשטף הגליקוליטי בספרואידים תאיים
כפי שניתן לראות בדרך כלל בספרות או במידע של יצרן הטכנולוגיה, הקצב הגליקוליטי של הספרואידים, הנמדד כ-ECAR, הוא פרמטר משני שניתן ללכוד לצד זיהוי תווים אופטי (OCR). חישוב ECAR לבדו אינו פרמטר שימושי או משמעותי בכל ניסוי XF מכיוון שהוא אינו מתוקן עבור יכולת האגירה של מאגר הבדיקה XF או תוספת של החמצה מיטוכונדריאלית, הנובעת מהידרציה של CO2 ל- HCO3 ו- H+. ECAR הוא תובנה רק לאחר שתיקוני נתונים אלה מיושמים, ולאחר מכן ניתן לספק מסקנות מדויקות יותר על השטף הגליקוליטי. כדי לתקן את יכולת האגירה כדי ליצור נתוני PER משמעותיים יותר, יש לדעת את נפח המיקרו-צ'מבר עבור המיקרו-פלטה הספרואידית. היצרן לא הצליח לספק נפח אמיתי לכך עם המיקרו-לוחית הספרואידית, ולכן לא ניתן לקבוע נתוני PER בקלות. ואכן, למרות שניתן היה להשיג מדידות אלה באופן אמפירי, זה היה מעבר להיקף של כתב יד זה. עם זאת, עם התיקונים המתאימים וידיעת נפח המיקרו-צ'מבר עבור גודל ספרואיד נתון הנמצא (למשל, קבלת מידה של צפיפות ספרואידים) בבאר, נתוני ECAR יהפכו למשמעותיים, וניתן יהיה לבצע חישובים של PER גליקוליטי. לפיכך, נתוני XF יכלו להיות אינפורמטיביים יותר לחקר חילוף החומרים הגליקוליטי והחמצוני בספרואידים, אך רק אם פרמטרים אלה נחשבו לעומק.
היווצרות, טיפול, העברה ותנועה של ספרואידים
קווי תאים מסוימים מתאימים יותר להיווצרותם של ספרואידים מאחרים, וייתכן שאינם יוצרים ספרואידים כלל, לדוגמה, תאי סרטן השחלות MCF-741,42 יוצרים ספרואידים מעגליים מאוד בהשוואה לקווי תאים אחרים (איור 3). כדוגמה נוספת, הודגם כי תאי סרטן הלבלב Capan-1 יוצרים כדוריות טובות יותר מאשר Panc-1 או BxPC343. באופן דומה, ידוע כי לקווי תאי קרצינומה בכבד יש יכולות משתנות ליצור ספרואידים קומפקטיים 5,44, עם שינוי נצפה בפנוטיפ כגון חילוף חומרים משופר של תרופות או ייצור אלבומין, כמו במקרה של HepG2 לעומת HepG2/C3A 9,45,46 או כדוריות HepaRG 17,47,48 . לכן, על המשתמשים לייעל את טכניקות התרבות הספרואידית בהתאם ולבצע ניסויי טיטרציה כדי לקבוע את צפיפות הזריעה האופטימלית ואת מהלך זמן הטיפוח. בנוסף, הפורמולציה וההרכב של מדיית הבדיקה הוכחו כמשפיעים על פורמולציה ספרואידית, כולל תוספת של מתיל צלולוז, שלעתים קרובות נוספה למדיה כדי להגדיל את צמיגות המטריצה 43,49,50. לפיכך, יש לקבוע את הרכב התווך התאי האופטימלי באופן אמפירי עבור כל קווי התאים שבהם נעשה שימוש.
מספר חילופי המדיום בתרבית הספרואידים נקבע על ידי קו התאים שבו נעשה שימוש. עם זאת, בדרך כלל, חילופים בינוניים בחצי נפח כל 2-3 ימים ישימים ברוב המקרים כדי לחדש את מלאי החומרים המזינים. השתמשנו בגישת הצבירה הכפויה כדי ליצור ספרואידים תלת-ממדיים באמצעות מיקרו-לוחות דוחי תאים ממקורות מסחריים זמינים לפיתוח ופריסה מהירים של מודלים ספרואידיים במחקרי ניתוח XF. עם זאת, פלטפורמות חלופיות עשויות להתאים יותר ליצירת ספרואידים מסוגי תאים אחרים, למשל, גישות תלויות-נפילה או מטריצה משובצות. במעבדות מוגבלות במשאבים, ייתכן שהמשתמשים ירצו להביט לעבר טכניקת שכבת-העל האגרוזית-נוזלית ליצירת משטחי מיקרו-פלטה דוחי תאים51,52 כדי להפחית באופן משמעותי את העלויות הכלכליות של שלבי הפיתוח הראשוניים של שיטת הספרואידים. התנועה של ספרואידים בין כלי תרבית נחוצה כדי לבצע ניתוח XF ומבחנים אחרים במורד הזרם. קלות ההעברה מוכתבת בדרך כלל על ידי גודל ספרואידים וצפיפות כוללת. אנו ממליצים להשתמש בקצה פיפטה רחב P200 או P1000 כדי לשמור על שלמות הספרואידים; קצוות פיפטה קטנים יותר מסתכנים בהפרעה מכנית של הספרואיד, אשר ניתן לקנות אותו באופן מסחרי או, בזהירות, נעשה על ידי חיתוך פשוט של קצה הפיפטה כדי להגדיל את הפתח. עם זאת, גישה זו עלולה להיות עלולה להחדיר פרווה לפלסטיק סביב קצהו, מה שעלול לגרום להפרעה מכנית במהלך הטיפול. השימוש בתאורה אחורית או תיבת אור שימושי גם לטיפול ותצפית של ספרואידים תחת מיקרוסקופ דיסקציה כצעד חיוני כדי להבטיח העברה מוצלחת של כדוריות לתוך מיקרו-לוחית הבדיקה של הספרואידים. יתר על כן, המיקום הספרואידי בתוך הבאר של מיקרו-לוחית בדיקה ספרואידית הוא בעל חשיבות מיוחדת ומשפיע ישירות על זיהוי תווים אופטי (OCR) ועל אפקט התרכובת במהלך בדיקת MitoStress טיפוסית (איור 7), ככל הנראה בשל הקשר בין מיקום הספרואיד לבין הבדיקה החיישן פלואורופורים.
בארות תיקון רקע ובקרת טמפרטורה
השימוש במבחנים מבוססי מיקרו-לוחות הוא גישה נפוצה במספר תחומי מחקר; עם זאת, השימוש בהם מציב מספר אתגרים מעשיים. כפי שקורה בגישות ניסיוניות אחרות, במיוחד אלה המשתמשות בתבנית מערך 96 (או יותר), הגיאומטריה והמיקום של מיקרו-לוחות יכולים להשפיע על שיפועי טמפרטורה וחילופי גזים על פני הלוח לאורך זמן, המכונים לעתים קרובות 'אפקטי קצה'53,54. מצאנו שזה נכון גם לגבי המיקרו-לוחית של הבדיקה הספרואידית. על פי ההנחיות והפרוטוקולים של היצרן, הבארות הפינתיות החיצוניות ביותר: A1, A12, H1 ו- H12 מוגדרות תמיד כבארות תיקון רקע ובקרת טמפרטורה עבור מנתח XFe96. לעומת זאת, עם פורמט מערך 24 הקידוחים, A1 ו-D6 מוגדרים כבארות בקרה, לצד שתי בארות נוספות הפרוסות באופן שווה על פני מרכז הצלחת ב-B4 ו-C3. בביצוע ניתוח XF ספרואידי, מצאנו סטייה משמעותית בנתונים שנאספו בתחילה באמצעות הנחיית היצרן. זאת למרות הכללת הצעדים הדרושים כדי להבטיח בדיקה מראש של שיווי משקל לטמפרטורה ולתכולת CO2 לפני תחילת רכישת הנתונים, ולעתים קרובות הניבו ערכים שליליים עבור זיהוי תווים אופטי (OCR) בעקבות הזרקה של מעכבי נשימה מסוימים (איור 8).
מצאנו שהתצפיות האלה ככל הנראה נובעות מהשפעות קצה על פני המיקרו-לוחית של הבדיקה הספרואידית. באיור 8 מצאנו שהפצה מחדש של בארות בקרת הרקע על פני המיקרו-לוחית, נתוני XF הותאמו בערך פי 2. שתי הסיבות הסבירות ביותר הן (1) בשל השפעות אידוי בבארות הקצה וכתוצאה מכך תא נפח כולל קטן יותר עבור הגשושית XFe96 לדגום ממנו, ו-(2) משיווי משקל טמפרטורה לא הולם בין בארות אלה המיועדות לתיקון רקע ולבארות דגימה, וכתוצאה מכך מערכי נתונים שמסווים או מנפחים יתר על המידה את זיהוי התווים האופטי .OCR. כדי להימנע מתוצאות כאלה, מומלץ, לפיכך, במיוחד בהקשר של ניתוח ספרואידים, שמשתמשים יחלקו מחדש בארות המיועדות לתיקון רקע על פני כל המיקרו-פלטה של הבדיקה הספרואידית וינקטו בצעדים הדרושים כדי לאזן מראש את הבדיקה שלהם לפני רכישת נתוני XF.
נורמליזציה של נתונים
בנוסף למתן פרוטוקול מפורט לבדיקת חילוף החומרים של אנרגיה מיטוכונדריאלית של ספרואידים תלת-ממדיים בודדים בטכנולוגיית XF, מאמר זה מציג גם דרכים אפשריות לנרמל את נתוני קצב הנשימה המיטוכונדריה המתקבלים עם ספרואידים תלת-ממדיים. באמצעות נתוני קצב הנשימה המתקבלים עם ספרואידים MCF-7 בתרבית בצפיפות זריעת תאים שונות (איור 3), אנו מציגים קצבי נשימה מיטוכונדריאליים בסיסיים מספרואידים MCF-7 בעלי גודל וקוטר הולכים וגדלים כאשר הם מנורמלים לצפיפות הזריעה הראשונית של התאים, נפח הספרואידים ותכולת ה-dsDNA (איור 9). שיטת הנורמליזציה המתאימה היא בעלת חשיבות עליונה לפרשנות מדויקת של מערכי נתונים של XF, במיוחד כאשר משווים מודלים ספרואידיים תלת-ממדיים במבחנה וסוגי תאים שונים. נורמליזציה לקויה עלולה להוביל לתוצאות שגויות שפשוט לא ניתן להשוות בין מערכי נתונים. תכולת החלבון אינה מועדפת לנורמליזציה של נתוני XF ספרואידיים, שכן טיפולים מקדימים עשויים להשפיע על שיעורי סינתזת החלבון ללא השפעה משמעותית על קצב הנשימה. יתר על כן, כמויות משמעותיות ולא עקביות של חלבון יכולות להיקשר למיקרו-לוחות כדוריים עם תזת התא, מה שמכניס שונות בתכולת החלבון בין בארות. זה עלול להיות מסובך עוד יותר בניתוחי XF באמצעות ספרואידים או תאים לא-אדומים הדורשים דבקים ביומולקולריים כדי להיקשר אליהם, שעשויים להכיל חלבון.
בניגוד לתכולת החלבון התוך-תאית, תכולת הדנ"א הגרעיני אינה תלויה בסוג התא והיא פרופורציונלית למספר התא (איור 9D) – גישה מדויקת יותר וגוזלת פחות זמן מאשר פירוק של ספרואידים לצורך כימות מספר התא. לעומת זאת, Yepéz et al.55, שערכו ניתוחי XF בחד-שכבתיים של תאי פיברובלסטים, מצאו כי נורמליזציה של נתוני XF למספר התאים הציגה פיזור גדול יותר של נתונים מאשר לפני הנורמליזציה. תכולת הדנ"א הגרעיני אינה תלויה במצב מובחן או בפנוטיפ ולכן היא מדויקת יותר לנורמליזציה של נתונים ספרואידיים במבחני XF מאשר לתכולת חלבונים. תוכן הדנ"א היה גם אסטרטגיה מוכחת לניתוח מערכי נתונים אחרים הקשורים לחילוף חומרים56. עם זאת, חשוב לציין שתכולת הדנ"א הגרעינית מכמתת מכל התאים הנמצאים בתוך הספרואיד; לכן, נורמליזציה לתכולת הדנ"א אינה מומלצת עבור מערכי נתונים של XF שבהם ספרואידים עוברים טיפולים שעלולים לגרום לאובדן משמעותי של כדאיות התאים. עבור מערכי נתונים כאלה, אם הדבר אפשרי, עדיפה נורמליזציה לכדאיות התאים, או שניתן לתקן את הנתונים באופן בסיסי לנשימה בסיסית.
שימוש ביכולת הנשימה הרזרבית כמופת לחשיבות של נורמליזציה של נתונים
קיבולת הנשימה הרזרבית היא מדד לקצב הקיבולת הנשימתית המיטוכונדרית המקסימלית פחות קצב הנשימה המיטוכונדריאלי הבסיסי (איור 6). עם זאת, הבעיה עם דיווח על נתונים מסוג זה כקצב, כלומר, pmolO2/min/well בניסויים מסוימים, היא שהנתונים ריקים מנורמליזציה. גם אם מנרמלים את נתוני הספרואידים לצפיפות התא/תכולת הדנ"א, לעתים קרובות הדבר אינו כולל את הפרמטר המרכזי שיש לנרמל עבור צפיפות המיטוכונדריה בתוך התאים. בהתחשב בכך ששינוי בצפיפות המיטוכונדרית יוביל לשינוי פרופורציונלי בנשימה הבסיסית והמקסימלית, גם הקיבולת הרזרבית תגדל. לדוגמה, אםה-OCR הספרואידי הבסיסי הוא 200 וה-OCRהמרבי הוא 400, הקיבולת הפנויה מדווחת כ-200; אםבסיס OCR הוא 100 ו OCRמקסימום, אז קיבולת רזרבית היא גם 100; עם זאת, כאחוז, שניהם 50% מהמקסימום (או 100% מהבסיסי). לכן, הקיבולת הרזרבית אינה משתנה בין שתי הדוגמאות הללו, למרות הבדלים בשיעורים של 200 ו-100 כאשר מחושבים כ-pmols O2/min/well. ערכים מנורמלים פנימיים הם אמינים יותר ובעלי תובנות רבות יותר כדי להפוך את נתוני ה- XF לדומים יותר במחקרים ובפרויקטים שונים. כדי לעשות זאת עבור קיבולת הנשימה הרזרבית, בחרנו להציג זאת כאחוז של נשימה מקסימלית במקום כשיעור מוחלט. זה יכול להיות מוצג גם כאחוז של נשימה בסיסית. זה יהיה המקרה אם עובדים עם תאים או ספרואידים. עם זאת, בהתחשב בכך שהמיקום של הספרואיד בצלחת המיקרווול עשוי לשנות את ה-OCR המוחלט אך לא את השינויים היחסיים עם מעכבים או uncouplers, חשוב יותר להסתכל על תגובות מנורמלות פנימיות בספרואידים כשינוי קיפול או אחוזים.
המודלים הספרואידיים שנוצרו כאן מציגים מגוון של סוגי תאים וארכיטקטורה שלא ניתן ללכוד אותם במודלים דו-ממדיים קלאסיים. אלה כוללים סידור מרחבי הטרוגני של תאים בשלושה ממדים, מגעים משופרים בין תאים לתאים (למשל, היווצרות צמתי רווחים ומטריצות חוץ-תאיות), ושיפועים ביוכימיים לאורך קוטר הספרואיד (למשל, גרדיאנטים של pH, גישה לפיזור חמצן לחומרים מזינים). שימוש בשטף חוץ-תאי לחקר ביולוגיה של ספרואידים במבחנה יכול לאפשר זיהוי מטרות אופטימליות לטיפולים תרופתיים באמצעות תצפיות על הפרעה מטבולית. ניתן לבצע אקסטרפולציה של אלה מספרואידים במבחנה לגידולי in vivo ולזהות מסלולים שעשויים להתמקד במטבוליזם של גידולים ספרואידים, למשל, ניצול פחמימות במהלך גדילה ספרואידית. שיטות טיפוליות עשויות להיות יעילות במיקוד ספרואידים בשלבי גדילה מוקדמים, אך להוכיח יעילות פחותה בשלבים המאוחרים יותר של צמיחת הספרואידים ככל שמורכבות הרשת המטבולית מבשילה. לסיכום, פיתוח מודלים של תרביות תאים תלת-ממדיות וטכנולוגיות ניתוח מתוחכמות במחקר ביולוגי ימשיכו להיות תחום דינמי ומשתנה במהירות עם פוטנציאל שאין שני לו. ניתוח שטף חוץ-תאי של ספרואידים מתרביות תאים במבחנה יכול לשמש כשיטת מחקר חדשנית לקידום תוצאות מחקר שניתן לבצע אקסטרפולציה כדי להבין טוב יותר את הביולוגיה הרלוונטית לאדם, להפחית את השימוש במודלים של בעלי חיים במחקר ולשפר את המחקר הממוקד בחולה.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
למחברים אין ניגודי עניינים להצהיר עליהם.
Acknowledgments
N.J.C נתמך על ידי פרס מקרה BBSRC MIBTP עם Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
חקר הסרטן גיליון 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
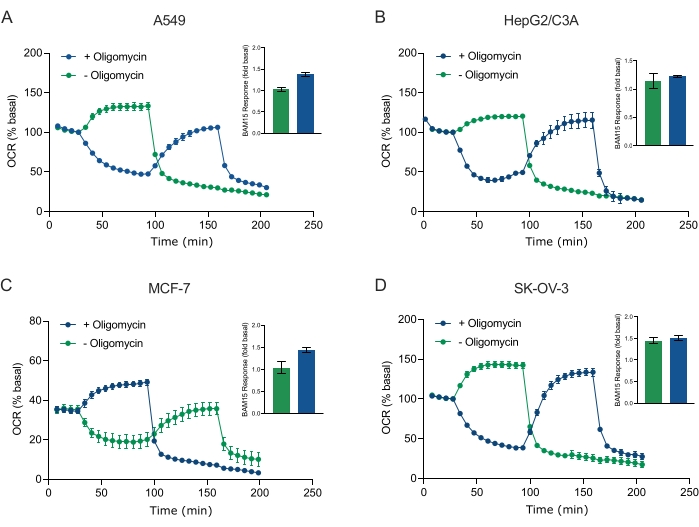
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
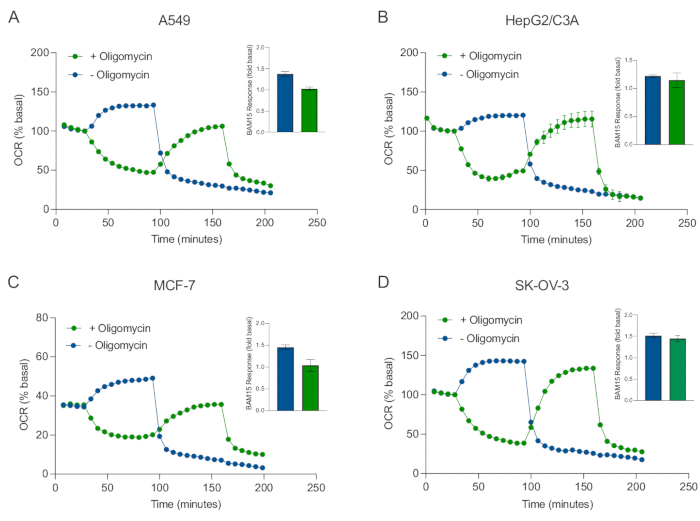
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
