

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Disse protokoller vil hjælpe brugerne med at undersøge mitokondrie energimetabolisme i 3D-kræftcellelinjeafledte sfæroider ved hjælp af Seahorse ekstracellulær fluxanalyse.
Abstract
Tredimensionelle (3D) cellulære aggregater, kaldet sfæroider, er blevet forkant med in vitro-cellekultur i de senere år. I modsætning til dyrkning af celler som todimensionale, encellede monolag (2D-kultur) fremmer, regulerer og understøtter sfæroid cellekultur fysiologisk cellulær arkitektur og egenskaber, der findes in vivo, herunder ekspression af ekstracellulære matrixproteiner, cellesignalering, genekspression, proteinproduktion, differentiering og proliferation. Betydningen af 3D-kultur er blevet anerkendt inden for mange forskningsområder, herunder onkologi, diabetes, stamcellebiologi og vævsteknik. I løbet af det sidste årti er der udviklet forbedrede metoder til at producere sfæroider og vurdere deres metaboliske funktion og skæbne.
Ekstracellulære flux (XF) analysatorer er blevet brugt til at udforske mitokondriefunktion i 3D-mikrotissuer såsom sfæroider ved hjælp af enten en XF24-ø-indfangningsplade eller en XFe96-sfæroid mikroplade. Imidlertid er forskellige protokoller og optimering af sonderende mitokondrieenergimetabolisme i sfæroider ved hjælp af XF-teknologi ikke beskrevet detaljeret. Dette papir indeholder detaljerede protokoller til sondering af mitokondrieenergimetabolisme i enkelt 3D-sfæroider ved hjælp af sfæroidmikroplader med XFe96 XF-analysatoren. Ved hjælp af forskellige kræftcellelinjer er XF-teknologi påvist at være i stand til at skelne mellem cellulær respiration i 3D-sfæroider af ikke kun forskellige størrelser, men også forskellige volumener, cellenumre, DNA-indhold og type.
De optimale mitokondrie effektor sammensatte koncentrationer af oligomycin, BAM15, rotenon og antimycin A anvendes til at undersøge specifikke parametre for mitokondrie energimetabolisme i 3D sfæroider. Dette papir diskuterer også metoder til normalisering af data opnået fra sfæroider og behandler mange overvejelser, der bør overvejes, når man udforsker sfæroid metabolisme ved hjælp af XF-teknologi. Denne protokol vil hjælpe med at drive forskning i avancerede in vitro sfæroidmodeller.
Introduction
Fremskridt inden for in vitro-modeller inden for biologisk forskning har udviklet sig hurtigt i løbet af de sidste 20 år. Sådanne modeller omfatter nu organ-on-a-chip-modaliteter, organoider og 3D-mikrotissu-sfæroider, som alle er blevet et fælles fokus for at forbedre translationen mellem in vitro- og in vivo-undersøgelser. Anvendelsen af avancerede in vitro-modeller, navnlig sfæroider, spænder over flere forskningsområder, herunder vævsteknik, stamcelleforskning, kræft og sygdomsbiologi 1,2,3,4,5,6,7 og sikkerhedstest, herunder genetisk toksikologi 8,9,10, nanomaterialer toksikologi11 12,13,14 og lægemiddelsikkerhed og effektivitetstest 8,15,16,17,18,19.
Normal cellemorfologi er afgørende for biologisk fænotype og aktivitet. Dyrkning af celler i 3D-mikrotissue-sfæroider gør det muligt for celler at vedtage en morfologi, fænotypisk funktion og arkitektur, mere beslægtet med den, der observeres in vivo , men vanskelig at fange med klassiske monolagscellekulturteknikker. Både in vivo og in vitro påvirkes cellulær funktion direkte af det cellulære mikromiljø, som ikke er begrænset til cellulær kommunikation og programmering (f.eks. Celle-celle-krydsdannelser, muligheder for at danne cellenicher); celleeksponering for hormoner og vækstfaktorer i de umiddelbare miljøer (f.eks. cellulær cytokineksponering som en del af en inflammatorisk reaktion); sammensætning af fysiske og kemiske matricer (f.eks. om celler dyrkes i stiv vævskulturplast eller et elastisk vævsmiljø) og vigtigst af alt, hvordan cellulær metabolisme påvirkes af ernæring og adgang til ilt samt behandling af metaboliske affaldsprodukter såsom mælkesyre.
Metabolisk fluxanalyse er en kraftfuld måde at undersøge cellulær metabolisme inden for definerede in vitro-systemer . Specifikt giver XF-teknologi mulighed for analyse af levende ændringer i realtid i cellulær bioenergetik af intakte celler og væv. I betragtning af at mange intracellulære metaboliske hændelser forekommer inden for få sekunder til minutter, er funktionelle tilgange i realtid afgørende for at forstå realtidsændringer i cellulær metabolisk flux i intakte celler og væv in vitro.
Dette papir indeholder protokoller til dyrkning af kræftafledte cellelinjer A549 (lungeadenocarcinom), HepG2 / C3A (hepatocellulært carcinom), MCF-7 (brystadenocarcinom) og SK-OV-3 (ovarieadenocarcinom) som in vitro 3D-sfæroidmodeller ved hjælp af tvungen aggregeringsmetoder (figur 1). Det beskriver også (i) detaljeret, hvordan man undersøger mitokondrieenergimetabolisme af enkelt 3D-sfæroider ved hjælp af Agilent XFe96 XF-analysatoren, (ii) fremhæver måder at optimere XF-assays ved hjælp af enkelt 3D-sfæroider og (iii) diskuterer vigtige overvejelser og begrænsninger ved sondering af 3D-sfæroidmetabolisme ved hjælp af denne tilgang. Vigtigst af alt beskriver dette papir, hvordan datasæt indsamles, der gør det muligt at beregne iltforbrugshastighed (OCR) for at bestemme oxidativ phosphorylering og dermed mitokondriefunktion i cellulære sfæroider. Selvom det ikke analyseres for denne protokol, er ekstracellulær forsuringshastighed (ECAR) en anden parameter, der måles sammen med OCR-data i XF-eksperimenter. ECAR fortolkes dog ofte dårligt eller forkert fra XF-datasæt. Vi giver en kommentar til begrænsningerne ved beregning af ECAR efter grundlæggende tilgange fra teknologiproducenten.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figur 1: Grafisk arbejdsgang til generering af cellulære sfæroider, ekstracellulær fluxanalyse og downstream-assays. Fire kræftcellelinjer blev selektivt dyrket som monolag (A), løsrevet fra vævskulturkolber og podet i ultralave vedhæftede 96-brønd mikroplader til dannelse af sfæroider (B). A549 lungecarcinom, HepG2 / C3A levercarcinom, SK-OV-3 ovarieadenocarcinom og MCF-7 brystcarcinomceller blev podet ved 1 × 103-8 × 103 celler / brønd og vokset op til 7 dage for at danne enkeltsfæroider og optimere sfæroidsåningstæthed og dyrkningstid ved kontinuerlig observation og planimetriske målinger. Når de var dannet, blev enkeltsfæroider vasket i et serumfrit XF-medium og omhyggeligt podet i sfæroid assaymikroplader, forbelagt med poly-D-lysin (C). Sfæroider blev udsat for ekstracellulær fluxanalyse ved hjælp af XFe96-analysatoren ved hjælp af flere protokoller til adressering af: (1) optimal sfæroidstørrelse til basal mitokondriel respirationsrespons; (2) optimeret titrering af mitokondrie respiratoriske hæmmere; (3) optimering af sfæroid placering i mikropladebrønde. (D) Post XF-analyser, fasekontrastmikroskopi og sfæroid DNA-kvantificering blev anvendt til datanormalisering og andre downstream in vitro-assays . Klik her for at se en større version af denne figur.
1. Dyrkning af kræftcellelinjer som 3D in vitro sfæroider
| Celle linje | Beskrivelse | Kultur medium | Kilde |
| A549 | Lungekarcinom celle linje | Rpmi 1640 | Europæisk samling af autentificerede cellekulturer (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Hepatisk karcinomcellelinje, et klonalt derivat af den overordnede HepG2-cellelinje | Dmem | Amerikansk vævskultursamling (ATCC) |
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| McF7 | Bryst adenocarcinom celle linje | Rpmi 1640 | Europæisk samling af autentificerede cellekulturer (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Ovarie adenocarcinom celle linje | Rpmi 1640 | Europæisk samling af autentificerede cellekulturer (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| Komponent | RPMI-assaymedium (50 ml endeligt volumen) | ||
| Basis medium | Agilent Søhest XF RPMI, pH 7,4 | ||
| Glukose (1 M sterilt lager) | 11 mM (0,55 ml stamopløsning) | ||
| L-glutamin (200 mM sterilt lager) | 2 mM (0,5 ml stamopløsning) | ||
| Natriumpyruvat (100 mM sterilt lager) | 1 mM (0,5 ml stamopløsning) |
Tabel 1: Kræftcellelinjemedier og XF-mediesammensætninger.
- Kultur alle cellelinjer ved hjælp af standard aseptisk vævskulturteknik og bekræft, at de er fri for mycoplasma ved hjælp af et passende assay kit.
- Cellelinjerne i T75-vævskulturkolber eller tilsvarende dyrkes ved hjælp af det anbefalede medium (tabel 1). Kultur cellelinjerne til 65-80% sammenløb og passere dem regelmæssigt op til maksimalt 25 passager.
- Cellekulturkolberne skylles to gange i Dulbeccos modificerede fosfatbufferede saltvand (DBPS).
- Cellerne løsnes fra kolberne med 3 ml celledissociationsreagens (se materialetabellen) i 5 minutter ved 37 °C, og løsrivelsen bekræftes ved mikroskopi.
- Aspirer den løsrevne cellesuspension forsigtigt for at sikre en enkeltcellesuspension og deaktiver celledissociationsreagenset med 7 ml komplet vævskulturmedium.
- Saml cellerne ved centrifugering ved 300 × g i 5 minutter, kassér supernatanten, og resuspend cellerne i komplet medium.
- Tæl cellerne ved hjælp af et hæmocytometer eller en automatiseret celletæller og titrere til den ønskede celletæthed, der kræves til såning.
BEMÆRK: For at så en hel 96-brøndplade ved 100 μL / brønd ved 4 × 103 celler / brønd, skal celler titreres til 4 × 104 celler / ml i et anbefalet volumen på 12 ml. - Cellesuspensionen dekanteres i et sterilt reservoir, og 100 μL af cellesuspensionen hældes i hver brønd i en celleafvisende mikroplade ved hjælp af en multikanals pipettor.
BEMÆRK: Kun de indre 60 brønde i en mikroplade skal sås og resten fyldes med DPBS. Dette vil danne en fordampningsbarriere, sikre sfæroid homogenitet over pladen og minimere pladekanteffekter. - Centrifuger sfæroidmikroplader ved 300 × g i 15 minutter for at tvinge cellerne til løse aggregater.
- Pladerne inkuberes ved 37 °C, 5 % CO2 i mindst 3 dage for at sikre dannelse af kugler.
- Udfør fasekontrastmikroskopi ved hjælp af standardiseret laboratoriepraksis for at overvåge væksten af sfæroider. Genopfyld cellekulturmediet hver 3. dag eller to gange om ugen ved at udføre en halvvolumen mediumudveksling.
2. Sondering af mitokondrie energimetabolisme af enkeltsfæroider ved hjælp af ekstracellulær flux (XF) teknologi
- Forberedelse af analyse (en dag før)
- Kontroller sfæroid levedygtighed ved hjælp af et omvendt lysmikroskop med fasekontrast ved 4x forstørrelse for at sikre intakt sfæroid struktur, morfologi og generel ensartethed mellem prøver.
- Hydrere sensorpatronen.
- Aliquot ~ 20 ml af kalibranten i et konisk rør.
- Det koniske rør indeholdende kalibranten anbringes i en ikke-CO2 37 °C inkubator natten over.
- Fjern indholdet fra analysesættet.
- Fjern sensorpatronen fra brugspladen, og placer den på hovedet på bordpladen ved siden af brugspladen.
- Pipetter 200 μL steril ddH2O i hver brønd i sensorpatronens hjælpeplade ved hjælp af en multikanals P300-pipette.
- Placer sensorpatronen oven på brugspladen.
- Kontroller, at vandstanden i hver brønd er høj nok til at nedsænke sensorsonderne.
- Overfør den samlede sensorpatron til en ikke-CO2 37 °C inkubator, og lad den stå natten over.
BEMÆRK: Dette trin kan udføres 12-72 timer før analysen påbegyndes.
- Frakke sfæroid assay mikroplade
- Ved hjælp af aseptiske teknikker tilsættes 30 μL/brønd steril poly-D-lysin (0,1 mg/ml) opløsning til den sfæroide mikroplade og inkuberes i 30 minutter ved stuetemperatur.
- Opsuge opløsningen fra hver brønd i den sfæroide mikroplade, vend pladen, og bank den fast på silkepapir for at fjerne enhver resterende opløsning.
- Pladen vaskes to gange med 200 μL/brønd steril ddH2O.
- Efter den sidste vask skal du vende mikropladen og banke den fast på silkepapir for at fjerne eventuelt resterende vand.
- Lad pladen lufttørre i 30 minutter, før den bruges eller opbevares ved 4 °C til fremtidig brug.
BEMÆRK: Den sfæroide assaymikroplade skal være belagt med et molekylært klæbemiddel for at sikre, at sfæroiderne er fastgjort i bunden af mikropladen. Uden et molekylært klæbemiddel kan sfæroider løsnes og forstyrre analyseresultater. Andre molekylære klæbemidler kan også bruges som et alternativ til Poly-D-Lysin til forbelægning plader. Forlakerede plader kan opbevares ved 4 °C, men bør overlades til ligevægt til stuetemperatur, inden analysen påbegyndes.
- Forbered XF-analysemedium
- XF RPMI-medium, som beskrevet i tabel 1, og sterilfilter med et sprøjtefilter på 0,22 μm
- Forberedelse af assay (1 time før assay)
- Forvarm det supplerede XF RPMI-assaymedium til 37 °C.
- Forvarm den overtrukne sfæroidassaymikroplade i en ikke-CO2 37 °C inkubator eller tørbad.
- Forbered sensorpatronen.
- Tag det koniske rør, der indeholder kalibranten og sensorpatronen, ud af luftinkubatoren.
- Fjern sensorpatronen fra brugspladen, og placer den på hovedet på arbejdsfladen.
- Brug en P300 multikanalpipette til at suge vandet fra brugspladen og kassere det.
- Kalibrantopløsningen hældes i et sterilt reagensbeholder, og der tilsættes 200 μL/brønd af den forvarmte kalibrant til brugspladen ved hjælp af en P300 multikanalpipette.
- Tag sensorpatronen op, og læg den tilbage oven på brugspladen, så sensorerne er godt nedsænket i kalibranten.
- Overfør den samlede sensorpatron tilbage i den ikke-CO2 37 °C inkubator, indtil den er klar til at indlæse portindsprøjtningsopløsningerne.
- Vask sfæroiderne med assaymediet.
- Sfæroide kulturplader fjernes fra 37 °C, 5 % CO2-inkubatoren , og sfæroider observeres under mikroskopet for at sikre deres integritet inden de sfæroide overførselstrin.
- Ilæg alle huller i den sfæriske plade med 180 μL/brønd af forvarmt assaymedium, inklusive eventuelle baggrundskorrektionsbrønde.
- Fyld delvist en 7 cm petriskål med 3 ml analysemedium.
- Ved hjælp af en multikanalpipette fyldt med brede åbningspipettespidser overføres sfæroiderne fra 96-brønds kulturpladen til 7 cm petriskåle ved at indstille pipetten til et aspirationsvolumen på 10-50 μL.
- Frø sfæroider i den forbelagte sfæroidassaymikroplade.
- Ved hjælp af et dissektionsmikroskop og et lyskasseapparat overføres sfæroiderne fra petriskålen til den sfæroide assaymikroplade som beskrevet nedenfor.
- Volumenet af en enkeltkanals pipettor udstyret med en bred åbningspipettespids indstilles til 20 μL, og opsuges forsigtigt en enkelt sfæroid. Placer spidsen direkte i midten af hver brønd i den sfæroide assaymikroplade, og lad tyngdekraften eluere en enkelt sfæroid ind i midten af hver brønd, dvs. udstød ikke noget medium fra pipettespidsen og lad kapillærvirkning trække sfæroidet ud af pipettespidsen. For at bekræfte eluering kan indholdet af pipetten pipetteres tilbage i 7 cm petriskålen under mikroskopet.
BEMÆRK: Tyngdekraftselvitation af en enkelt sfæroid tager typisk 15-30 s afhængigt af sfæroid størrelse / densitet. I løbet af denne tid bør pipetten ikke fjernes. Eventuelle baggrundskorrektionsbrønde bør være fri for sfæroider og kun indeholde assaymedium. Under mikroskopet skal du bekræfte placeringen af hver sfæroid. Hver sfæroid bør ideelt set placeres i midten af hver brønd. - Når alle sfæroiderne er overført til den sfæroide assaymikroplade, overføres pladen til en ikke-CO2-inkubator ved 37 °C i mindst 1 time før analysen.
- Volumenet af en enkeltkanals pipettor udstyret med en bred åbningspipettespids indstilles til 20 μL, og opsuges forsigtigt en enkelt sfæroid. Placer spidsen direkte i midten af hver brønd i den sfæroide assaymikroplade, og lad tyngdekraften eluere en enkelt sfæroid ind i midten af hver brønd, dvs. udstød ikke noget medium fra pipettespidsen og lad kapillærvirkning trække sfæroidet ud af pipettespidsen. For at bekræfte eluering kan indholdet af pipetten pipetteres tilbage i 7 cm petriskålen under mikroskopet.
- Ved hjælp af et dissektionsmikroskop og et lyskasseapparat overføres sfæroiderne fra petriskålen til den sfæroide assaymikroplade som beskrevet nedenfor.
3. Klargøring og indlæsning af forbindelser i sensorpatronen til XF-assays
| Injektion strategi | Forbindelse (port) | XFe96 mikrobrønd startvolumen (μL) | Ønsket endelig brøndkoncentration | Portvolumen (μL) | Endelig XFe96 mikrowell volumen post injektion (μL) | Koncentration af arbejdslager |
| 1 | Oligomycin (A) | 180 | 3 ug/ml | 20 | 200 | 30 μg/ml |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycin A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycin A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabel 2: Mitokondrieforbindelseskoncentrationer til sondering af mitokondrieenergimetabolisme af enkelt 3D-sfæroider ved hjælp af XFe96 Analyzer.
- Arbejdslagerkoncentrationerne af hver forbindelse forberedes som anført i tabel 2 ved hjælp af fuldt suppleret, forvarmt XF RPMI-assaymedium.
- Orienter patronpladen (koblet til brugspladen) kolonnevis, 1-12 fra venstre mod højre.
- Hvis du bruger en læssevejledning, skal du placere den oven på patronpladen i henhold til brøndpålæsningsproceduren, f.eks. hvis port A indlæses først, skal du sikre dig, at A er synlig i øverste venstre hjørne af styrepinden.
- Arbejdsløsningen af hver forbindelse overføres til et egnet reservoir, og der doseres 20 μL i alle tilsvarende porte ved hjælp af en kalibreret P100 multikanalpipette. Gentag for hver forbindelse i de resterende porte.
BEMÆRK: Hvis der ikke bruges porte på sensorens patronplade, kan disse efterlades tomme eller fyldes med analysemedium. Hvis der kun anvendes et udvalg af et bestemt portbogstav, skal det sikres, at de andre porte, der svarer til det pågældende bogstav, er lastet med analysemedium. ellers vil luft blive injiceret i brønden, hvilket kompromitterer resultaterne i disse brønde. - Efter portindlæsning skal du fjerne pladeindlæsningsstyrene (hvis de bruges) og forberede analysatoren til ilægning af sensorpatronen.
BEMÆRK: Hvis analysen ikke køres umiddelbart efter isætning af portene, skal du sætte låget tilbage på sensorpatronen og sætte pladen tilbage i 37 °C luftinkubatoren, indtil den er klar til at indlæse i maskinen.
4. Analysedesign, injektionsstrategier og dataindsamling
- Kører analysen
- Tænd for analysatoren, og tilslut til controlleren (computeren).
BEMÆRK: Dette kan bekræftes af instrumentforbindelsesstatus i widgetpanelet i Wave Controller-softwaren. - Naviger til skabelonsiden i WAVE-softwaren, find analyseskabelonfilen til eksperimentet, og dobbeltklik for at åbne den.
BEMÆRK: Hvis analyseskabelonen ikke vises i visningen Skabeloner , skal du importere skabelonfilen til skabelonmappen fra et delt netværksdrev eller USB-flashdrev. - For at starte analysen skal du klikke på fanen Kør analyse .
BEMÆRK: Hvis gruppedefinitionerne er blevet korrekt fordelt på pladekortet, vil analysen være klar til at køre som angivet med det grønne kryds i højre side af siden. På dette tidspunkt kan eventuelle yderligere oplysninger indtastes på assayoversigtssiden eller den side, der er tom; fortsæt til næste trin. På grund af den forsinkede penetration af mitokondriemodulatorer i 3D-mikrotissu-sfæroider (figur 2) skal du bruge måleprotokoloplysningerne beskrevet i tabel 3.
- Tænd for analysatoren, og tilslut til controlleren (computeren).
| Måleperiode | Injektionsnummer og port | Oplysninger om måling | Varighed af periode (h:min:s) |
| Kalibrering | Finder ikke anvendelse | XF-analysatorer udfører altid denne kalibrering for at sikre, at målingerne er nøjagtige | 00:20:00 (dette er et gennemsnit og kan variere mellem maskiner) |
| Ligestilling | Finder ikke anvendelse | Ækvilibrering sker efter kalibrering, og det anbefales. | 00:10:00 |
| Basal | Finder ikke anvendelse | Cykler = 5 | 00:30:00 |
| Blanding = 3:00 | |||
| Vent = 0:00 | |||
| Foranstaltning = 3:00 | |||
| Oligomycin / BAM15 | Injektion 1 (Port A) | Cyklusser = 10 | 01:00:00 |
| Blanding = 3:00 | |||
| Vent = 0:00 | |||
| Foranstaltning = 3:00 | |||
| Rotenon + antimycin A | Injektion 2 (Port B) | Cyklusser = 10 | 01:00:00 |
| Blanding = 3:00 | |||
| Vent = 0:00 | |||
| Foranstaltning = 3:00 | |||
| Samlet tid: | 03:00:00 | ||
Tabel 3: Protokolopsætning til sondering af mitokondrieenergimetabolisme af enkelt 3D-sfæroider ved hjælp af XFe96 Analyzer.
- Klik på startkørsel for at åbne dialogboksen Gem placering .
- Indtast lagringsstedet for resultatfilen, og placer den samlede sensorpatron på den termiske bakke, der vises fra døren på siden af analysatoren. Vent på, at den termiske bakke åbnes automatisk, og at skærmen viser meddelelsen Load Calibrant Utility Plate . Før du følger vejledningen på skærmen, skal du sikre dig i) korrekt montering af sensorpatronen på hjælpepladen, ii) låget fjernes fra sensorpatronen og iii) korrekt orientering af sensorpatronen på brugspladen.
- Følg kommandoerne på skærmen for at starte kalibrering af sensorpatroner.
BEMÆRK: Den tid, det tager at gennemføre kalibreringen, er ca. 10-20 minutter (for assays ved 37 °C). - Efter kalibrering af sensorpatronen skal du lægge den sfæroide mikroplade i analysatoren ved at følge instruktionerne på skærmen på Wave Controller for at starte 12 minutters ækvilibreringstrinnet.
BEMÆRK: Grønne felter med hvide flueben angiver en 'god' kalibrering for den brønd. Hvis nogen brønde ikke giver en 'god' kalibrering, vil de blive angivet med en rød boks og et hvidt kryds. Sådanne brønde skal noteres og udelukkes fra enhver analyse, efter at analysen er afsluttet ved hjælp af fanen modifikationsassay . - Vent på, at analysatoren automatisk begynder at indsamle baselinemålinger, efter at maskinen har fuldført ækvilibreringstrinnet (som beskrevet i instrumentprotokollen).
- For at fuldføre eksperimentet skal du følge kommandoerne på skærmen på WAVE-controlleren.
BEMÆRK: Når den sfæroide mikroplade er fjernet fra analysatoren, skal sensorpatronen kasseres, og den sfæroidplade afsættes til yderligere analyse, hvis det er nødvendigt (f.eks. dobbeltstrenget (ds) DNA-kvantificering). Hvis mikropladen ikke er nødvendig til yderligere analyse, kan den kasseres sammen med sensorpatronen. - Vent på, at analysedialogboksen vises, og se resultaterne, eller vend tilbage til skabelonvisningen .
5. Data normalisering og analyse strategier - post assay normalisering og downstream assays (valgfri trin)
- Normalisering af data
- For at normalisere sfæroiddata henvises til rækken af protokoller, der er relevante for datanormaliseringsstrategier til beregning af sfæroidstørrelse og volumen og kvantificering af dsDNA i sfæroidassays. Disse er medtaget som supplerende filer; Se supplerende fil 1 og supplerende fil 2.
- Analyse af data
- Hvis du vil eksportere data til en af de automatiserede analysegeneratorer, skal du følge dataeksportkommandoerne på WAVE-controlleren og vælge den eksportgenerator, der svarer til analysetypen. Alternativt kan du eksportere datafilen og uploade den til Seahorse analytics.
BEMÆRK: Ulempen ved rapportgeneratorer og Seahorse-analyser er, at dataanalyse er begrænset til, hvordan XF-analysen er designet og ikke tillader, at der tages gennemsnit på tværs af målecyklusser. Manuel eksport af datasæt fra instrumentsoftwaren giver mulighed for brugerpræferencer i denne henseende. I betragtning af at injektionsstrategien til vurdering af mitokondrie respiration af 3D-sfæroider sandsynligvis vil afvige fra en typisk 'MitoStress' -test, er der udviklet en række regnearkskabeloner til at hjælpe med at analysere disse datasæt, der er specifikke for 3D-cellekulturer og vil blive leveret efter anmodning. Disse dataskabelonfiler vil give data om de vigtigste mitokondrie respiratoriske parametre, der er beskrevet og forklaret i figur 2. - For at analysere dataene skal du eksportere dataene som en regnearksrapport fra WAVE-controllersoftwaren og bruge en uafhængig regnearksskabelon til analyse.
- Hvis du vil eksportere data til en af de automatiserede analysegeneratorer, skal du følge dataeksportkommandoerne på WAVE-controlleren og vælge den eksportgenerator, der svarer til analysetypen. Alternativt kan du eksportere datafilen og uploade den til Seahorse analytics.

Figur 2: Skematiske deskriptorer for parametre afledt af ekstracellulære fluxdataanalyser. Forkortelse: OCR = iltforbrugshastighed. Klik her for at se en større version af denne figur.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
For at opnå velformede, kompakte sfæroider blev hver cellelinje optimeret individuelt til såningstæthed og dyrkningsvarighed (figur 3). A549, HepG2/C3A og SK-OV-3 cellelinjer dannede oprindeligt løse aggregater, der ikke gik videre til runde sfæroider med klart definerede perimeter før efter 7 dage i kultur. Omvendt kan MCF-7-celler danne sfæroider inden for 3 dage. Der var en klar sammenhæng mellem den indledende cellesåningstæthed og sfæroidvolumen efter dyrkningsperioden for alle sfæroidmodeller. Sfæroid størrelse og morfologi blev optimeret til såningstæthed. Morfologi og cirkularitet begyndte at falde med øget sfæroidstørrelse i alle modeller. Seeding strategier for cellelinjer blev optimeret ved 4 × 103 celler / brønd for A549 og SK-OV-3 celler; HepG2 /C3A-celler er tidligere blevet optimeret andre steder til 1 × 103 celler / brønd, og MCF-7-celler blev brugt ved 4 × 103 celler / brønd i alle assays. Ved optimerede såningsstrategier var sfæroidvolumen mellem 5,46 × 107 μm3 (SK-OV-3) og 1,45 × 108 μm3 (A549) (figur 3B). Alle sfæroidtyper havde en lineær korrelation mellem den oprindelige såtæthed og sfæroidvolumen, hvor A549 og HepG2/C3A havdeR2-værdier på henholdsvis 0,957 og 0,947. MCF-7 og SK-OV-3 sfæroidvolumener viste sig begge at have en større korrelation med den oprindelige såtæthed, R2 = 0,977 (figur 3A).
Sfæroid cirkularitet blev beregnet ved hjælp af billedplanimetri i FIJI-analysesoftware ved hjælp af de lange og korte sfæroiddiametre. Perfekt sfæroid symmetri havde cirkularitet = 1,0; afvigelse fra 1,0 indikerede et tab af cirkularitet (figur 3C). Cirkulariteten var større i MCF-7 sfæroider end andre modeller, hvor cirkulariteten blev opretholdt mellem 0,83 og 0,9 ved alle såtætheder. Til sammenligning var den ydre omkreds af SK-OV-3 sfæroider ikke så klart defineret, og sfæroidvolumen var signifikant mindre selv efter 7 dage i kultur, hvilket gav sfæroider med en maksimal cirkularitet på 0,61 ved en såtæthed på 4 × 103 / brønd. HepG2/C3A-celler viste sig også at danne tætte, velformede sfæroider med en jævn morfologi over overfladearealet af alle sfæroider, med cirkularitet opretholdt på 0,79 for celler podet ved 1 × 103 celler / brønd. A549-celler syntes at følge en tendens, hvor sfæroid cirkularitet og morfologi blev forbedret med såtæthed; cirkulariteten var dog ikke større end 0,63 ved den tæthed, der blev anvendt i disse eksperimenter.
Basal mitokondrie respiration blev beregnet som OCR målt fra sfæroider podet ved 1 × 103, 2 × 103, 4 × 103 eller 8 × 103 celler / brønd i ultralave vedhæftede sfæroidkulturmikroplader (figur 3D). For alle sfæroidtyper steg OCR med sfæroidstørrelse og var lineært korreleret med sfæroidvolumen med R2 højest i MCF-7 sfæroider ved 0,988 og lavest i SK-OV-3 sfæroider ved 0,744 (figur 3E). Den målte OCR var statistisk forskellig mellem alle eksperimentelle grupper. A549 havde den laveste OCR og opnåede kun 18 pmol/min/brønd ved den største sfæroidstørrelse (figur 3D). Omvendt gav MCF-7-sfæroider en lignende OCR ved den mindste sfæroidstørrelse efter kun 3 dage i kultur og nåede en maksimal baseline OCR på 53 pmol / min / brønd for den største sfæroidstørrelse (figur 3D). HepG2/C3A gav OCR-data, der var meget konsistente med sfæroidstørrelse og morfologi. I HepG2/C3A-sfæroider podet fra 1 × 103 celler/brønd nåede baseline OCR i gennemsnit 15 pmol/min/brønd og steg til maksimalt 52 pmol/min/brønd i de største sfæroider (figur 3D). OCR i SK-OV-3 sfæroider var kun signifikant mellem sfæroider dyrket fra 4 × 103 celler / brønd og 8 × 103 celler / brønd, med ringe forskel set i sfæroider dyrket mellem 1 × 103, 2 × 103 eller 4 × 103 celler. På trods af størrelsesforskelle var OCR-data meget ens mellem HepG2/C3A og MCF-7 sfæroider på alle størrelsespunkter. I forhold til sfæroidstørrelse (μm3) var baseline OCR af MCF-7-sfæroider sammenlignelig med hepG2/C3A-sfæroider dyrket over 7 dage fra 1.000 celler pr. Brønd.

Figur 3: Sfæroidvækstparametre bestemmer baseline mitokondrie respiration. (A) A549, HepG2 / C3A, MCF-7 og SK-OV-3 sfæroider blev optimeret til såningstæthed og deres vækst overvåget ved 1 × 103, 2 × 103, 4 × 103 og 8 × 103 celler / brønd i hver af fotomikrograferne, fra henholdsvis øverst til venstre til nederst til højre; skalastænger = 500 μm. (B) Sfæroidstørrelse blev beregnet ved hjælp af planimetriske data fra indsamlede fotomikrografer og sammenlignet ved hjælp af Pearsons korrelationsstatistik; stiplede linjer repræsenterer fordelingen af 95% konfidensinterval. (C) Sfæroid morfologi blev sammenlignet ved beregning af cirkularitet. (D) OCR blev målt 5x, hvorefter rotenon-antimycin A blev tilføjet for at tage højde for nonmitokondriel respirationsfrekvens ved hjælp af Agilent Seahorse XFe96-analysatoren. OCRbasal målt som OCR - OCRr/a blev sammenlignet mellem såtæthed (D) og sfæroidvolumen (E). Data er gennemsnit ± SEM fra 5-8 brøndreplikater pr. sfæroidtype og cellesåningstæthed. Forkortelser: OCR = iltforbrugshastighed; OCRbasal = Basal mitokondrie respiration; OCRr/a = OCR efter tilsætning af rotenon-antimycin A. Klik her for at se en større version af denne figur.
Koncentrations- og tidsforløbet for eksponering for respiratoriske modulatorer i XF-analyse er et kritisk trin i assayoptimering. Respiratoriske modulatorforbindelser-oligomycin, BAM15, en blanding af rotenon-antimycin A- eller en DMSO-køretøjskontrol blev sekventielt injiceret gennem sensorpatroninjektionsportene i mikropladebrønde indeholdende MCF-7-sfæroiderne (figur 4A). Fire målecyklusser blev gennemført for at bestemme den gennemsnitlige basale OCR på 30-40 pmol / min / brønd for alle prøvegrupper. I resten af assayets varighed blev respiratoriske modulatorer sekventielt tilsat hver 5. målecyklus for at opnå endelige brøndkoncentrationer på 0,5 μM (0,5 μg/ml oligomycin) efter injektion 1; 2,0 μM (2 μg/ml oligomycin) efter injektion 2; 5 μM (5 μg/ml oligomycin) efter injektion 3; og endelig en maksimal brøndkoncentration på 11 μM (11 μg/ml oligomycin) efter den fjerde sekventielle tilsætning. MCF-7 sfæroider reagerede ikke på køretøjets kontrol under hele eksperimentet (figur 4A). Basal OCR ændrede sig umiddelbart efter den første injektion af hver enkelt forbindelse ved den laveste koncentration på 0,5 μM eller 0,5 μg/ml oligomycin (figur 4B). OCR i MCF-7 sfæroider blev sænket med oligomycin fra 41 pmol/min/brønd til 23 pmol/min/brønd efter 5 målecyklusser efter den første injektion på 0,5 μg/ml (figur 4B).
Som reaktion på 0,5 μm BAM15 blev OCR øget fra 33 til 41 pmol/min/længe før den anden injektion (figur 4C). Til sammenligning sænkede kombinationen af rotenon plus antimycin A OCR fra 37 til 13 pmol/min/længe før den anden injektion (figur 4D). Kinetiske spor afslørede yderligere et stabilt lineært fald (oligomycin og rotenon-antimycin A) eller stigning (BAM15) i OCR. For alle sammensatte doseringsregimer blev der opnået en steady state OCR inden for 10-12 komplette målecyklusser (60-72 min) ved en samlet brøndkoncentration på 2 μM BAM15, 2 μM rotenon, 2 μM antimycin A og 2 μg / ml oligomycin (figur 4A). Iltforbruget nåede et steady state plateau ved ~ 19 pmol / min / brønd (oligomycin), 52 pmol / min / brønd (BAM15) og 10 pmol / min / brønd (rotenon-antimycin A) (figur 4A). Forøgelse af den sammensatte koncentration af oligomycin, BAM15 eller rotenon plus antimycin A havde yderligere ingen åbenbar effekt på OCR, som forblev konstant i resten af analysen. Disse data viser, at både sammensat koncentration og tidsforløb for eksponering for respiratoriske modulatorforbindelser bør overvejes til analyseoptimering ved anvendelse af 3D-sfæroider.

Figur 4: Titrering af respiratoriske modulatorforbindelser som et vigtigt trin til optimering af ekstracellulær fluxanalyse. (A) MCF-7 sfæroider blev podet ved 4 × 103 celler / brønd og dyrket over 3 dage, før de blev anbragt i brønde af en sfæroid assaymikroplade indeholdende XF RPMI og undersøgt for OCR ± mitokondriemodulatorer ved hjælp af XFe96-analysatoren. OCR blev målt 5x, hvorefter titreringer af enten køretøjskontrol, oligomycin (B), BAM15 (C) eller rotenon-antimycin A (D) blev tilsat for at hæmme mitokondrie ATP-syntase, bestemme maksimal respiratorisk kapacitet eller etablere henholdsvis nonmitokondrie respirationsfrekvens. Koncentrationen af hver mitokondriemodulator blev forøget over 4 individuelle titreringsinjektionsstrategier (0,5 μM, 1,5 μM, 3 μM og 6 μM; enheder for oligomycin er μg/ml) for at bestemme maksimal steady state OCR som reaktion på optimal sammensat koncentration. OCR blev målt i 5 målecyklusser mellem hver injektion. Data er gennemsnit ± SEM fra 5-8 individuelle brøndreplikater. Forkortelse: OCR = iltforbrugshastighed. Klik her for at se en større version af denne figur.
En af de største fordele ved XF-teknologi er evnen til at undersøge mitokondriefunktion i intakte celler og væv. For at undersøge specifikke aspekter af mitokondriefunktionen i celler og væv tilsættes mitokondriemodulatorer sekventielt til brønde i prøvemikropladen gennem de 4 tilgængelige injektionsporte på sensorpatronen. Den typiske sekvens af modulatorer, der anvendes til at undersøge mitokondrieparametre i XF-assays, er oligomycin, en protonofor (f.eks. FCCP eller BAM15) og en kombination af rotenon plus Antimycin A, som tilsættes sekventielt for at hæmme mitokondrie-ATP-syntasen, bestemme maksimal respiratorisk kapacitet og korrigere for henholdsvis nonmitochondriel respirationsfrekvens. Denne typiske sekvens af modulatortilsætninger kaldes MitoStress-testen af analyseteknologiproducenten. I betragtning af at oligomycin kan hæmme uafbrydelig stimuleret respiration i nogle cellemonolag20, undersøgte vi dette med kræftafledte 3D-sfæroider ved at måle ukoblet stimuleret OCR (OCRmax) før (enkelt) og efter (sekventiel) oligomycininjektion (figur 5A-D). OCRmax var ikke signifikant begrænset af tilsætning af oligomycin i sfæroider dannet af HEPG2/C3A eller SK-OV-3 (figur 5E og figur 5G). OCRmax blev imidlertid signifikant sænket i A549 og MCF-7 sfæroider efter en sekventiel injektion af BAM15 efter oligomycin sammenlignet med OCRmax opnået fra en enkelt injektion af BAM15 (figur 5F og figur 5H). Medmindre andet er kendt, anbefales det derfor at anvende separate brønde til behandling med oligomycin og uncoupler med en endelig tilsætning af rotenon og antimycin A, når man undersøger mitokondrieenergimetabolismen af 3D-sfæroider. Denne tilgang giver stadig mulighed for beregning af alle mitokondrieparametre som med en typisk MitoStress-test, hvor forbindelser tilsættes sekventielt.

Figur 5: Enkelt eller sekventiel injektion af mitokondrie respiratoriske forbindelser. Kræftcelleafledte sfæroider af MCF-7, HEPG2/C3A, SK-OV-3 og A549 blev anbragt i brønde af en XFe96 sfæroid mikroplade i XF RPMI og undersøgt for OCR ved hjælp af Agilent Seahorse XFe96 analysatoren. OCR blev målt 5x, hvorefter 2 μg/ml oligomycin (injektion Port A: grøn trace) eller 5 μM BAM15 (injektion Port A: blå spor eller injektionsport B: grøn spor) for at hæmme mitokondrie ATP-syntase og bestemme maksimal respirationskapacitet. Kinetiske OCR-data udtrykkes som % basal (A-D). Maksimal respiratorisk kapacitet (OCRmax) blev beregnet som en faktor for basal OCR ved ligningen: OCRmax = OCRBAM15 / OCRbasal. OCRmax blev opnået fra OCR-gennemsnit på tværs af målecyklusser 8-10 efter BAM15 injektion med (grønne søjler) og uden (blå søjler) oligomycin. Data er gennemsnit ± SEM fra 3-8 individuelle brøndreplikater på tværs af sfæroidassaymikropladen. Forkortelser: OCR = iltforbrugshastighed. Klik her for at se en større version af denne figur.
Ved hjælp af optimale cellesåningstætheder, sammensatte koncentrationer, injektionsstrategi og målecyklusperiode bestemt i disse optimeringseksperimenter (tabel 3) har vi udviklet en detaljeret protokol til nøjagtig sondering af basal mitokondrie respiration: OCRbasal (figur 6A), ADP phosphorylering respiration: OCRADP (figur 6B), lækage respiration: OCRomy (figur 6C), koblingseffektivitet (figur 6D ), maksimal respirationskapacitet: OCRmax (figur 6E) og ledig respirationskapacitet: OCRspare (figur 6F) ved hjælp af kræftafledte 3D-sfæroider.

Figur 6: Sondering af OCR med XF-teknologi for at etablere mitokondrieenergimetabolisme af kræftafledte sfæroider. Kræftcelleafledte sfæroider af MCF-7, HEPG2 / C3A, SK-OV-3 og A549 blev anbragt i brønde i en sfæroid assaymikroplade i XF RPMI og undersøgt til OCR ved hjælp af Agilent Seahorse XFe96-analysatoren. OCR blev målt 5x, hvorefter 2 μg/ml oligomycin eller 5 μM BAM15 og RA blev tilsat for at hæmme mitokondrie-ATP-syntase, bestemme maksimal respirationskapacitet og beregne henholdsvis den ikke-mitokondriale respirationsfrekvens. (A) Basal mitokondrie respiration (OCRbasal) blev beregnet som gennemsnittet af OCR fra de 3 målecyklusser før port A injektion. (B) Koblingseffektiviteten af oxidativ phosphorylering blev tilnærmet ved at udtrykke OCRADP (OCRbasal- OCRlækage) som en procentdel af OCRbasal. (C) ADP phosphoryleringsrespiration (OCRADP) blev målt som oligomycinfølsom OCR, beregnet ud fra den gennemsnitlige OCR på tværs af målecyklus 11-13 før injektion af BAM15. (D) Leakomy respiration (OCR-lækage) blev målt som OCR-ufølsom over for oligomycin, beregnet ud fra den gennemsnitlige gennemsnitlige OCR på tværs af målecyklus 11-13. (E) Maksimal respirationskapacitet (OCRmax) blev målt som den gennemsnitlige maksimale OCR målt efter INJEKTION AF BAM15. (F) Ledig respirationskapacitet blev beregnet ved at udtrykkeOCR-reserve (OCRmax - OCRbasal) som en procentdel af OCRbasal. OCR efter rotenon-antimycin A-injektion (OCRr/a) blev trukket fra alle hastigheder for at korrigere for nonmitokondriel OCR. Data er gennemsnit ± SEM fra 3-8 individuelle brøndreplikater på tværs af XFe96 sfæroidpladen. Forkortelser: OCR = iltforbrugshastighed; RA = 2 μM rotenon-2 μM antimycin A. Klik her for at se en større version af denne figur.
MCF-7 sfæroider dyrket fra 4 × 103 celler / godt over 3 dage blev brugt som model til at bestemme optimal overførsel, placering og analyse inden for sfæroid assaymikroplader. Ved hjælp af dimensioner, der blev leveret til den sfæroide mikroplade fra producenten, blev brøndoverfladen opdelt i tre zoneområder for optimal sfæroid placering (figur 7A), hvor zone 1 blev fremhævet som den optimale zone i midten af brønden. Ved omhyggelig pipettering ved hjælp af brede pipettespidser blev sfæroider overført til sfæroidpladerne og tilfældigt fordelt over brøndoverfladerne ved tyngdekraftselvuering (figur 7B). Hvor sfæroider blev omhyggeligt overført ved hjælp af tyngdekraftsevolution, kunne de fleste sfæroider typisk findes i zone 1-2 af mikropladen ved hjælp af de anbefalede overførselsteknikker fra producenten. Hvor sfæroider blev tvunget ud af pipettespidsen ved aspiration, blev sfæroider ofte placeret uden for disse zoner og kunne ikke ses ved hjælp af mikroskopi.
For at sammenligne sfæroidplaceringspositioner blev MCF-7-sfæroider overført til de sfæroide assaymikroplader i udpegede zoner 1-3 eller uden for zonen (figur 7A). Disse 4 brønde blev sporet gennem et kinetisk eksperiment OCR ved baseline og efter tilsætning af oligomycin, BAM15 eller rotenon-antimycin A (figur 7C). OCR blev beregnet ud fra gennemsnittet af tre cyklusaflæsninger før hver injektion (figur 7B). OCR blev målt kinetisk over 200 min i de 4 udvalgte brønde (figur 7C) og baseline-korrigeret (figur 7D). Hvor sfæroider blev placeret i zone 3 eller uden for zone, var baseline OCR'er signifikant lavere end sfæroider placeret i zone 1 og 2 (figur 7C). Virkningerne af respiratoriske forbindelser oligomycin, BAM15 og rotenon-antimycin A varierede også dramatisk mellem sfæroider placeret i zone 1 og 2 sammenlignet med zone 3 og regioner uden for zonen. En stigning i OCR blev set med oligomycin i sfæroider placeret i zone 3 eller uden for zone (figur 7E). Desuden oplevede sfæroider placeret i zone 3 eller uden for zone en for høj respons på BAM15 med OCR højere end baseline efter rotenon-antimycin A-injektion (figur 7E). På trods af en næsten dobbelt stigning i basal OCR (figur 7C) med sfæroider placeret i zone 2 versus zone 1, var foldændringerne som reaktion på alle respiratoriske forbindelser meget ens (figur 7E), hvilket tyder på, at forskelle i basal OCR mellem sfæroider placeret i zone 1 eller 2 sandsynligvis ikke er resultatet af placering i brønden.

Figur 7: Placering af sfæroider i den sfæroide assaymikroplade dikterer basale OCR- og mitokondriemodulatoreffekter ved hjælp af XF-teknologi. MCF-7 sfæroider blev podet ved 4 × 103 celler / brønd og dyrket over 3 dage, før de blev anbragt i brøndene i den sfæroide mikroplade indeholdende XF RPMI og undersøgt for OCR-± mitokondriemodulatorer ved hjælp af Agilent Seahorse XFe96-analysatoren. A) Fotomikrografer af sfæroide zonepositioner i sfæroid assaymikroplader efter analysens varighed. skala bar = 500 μm og OCR fanget fra tilsvarende brønde over tid udtrykt som enten pmol/min-1/well-1 (B) eller % basal (C). D) Mitokondriemodulatorvirkninger af MCF-7-sfæroider placeret i forskellige zoner inden for sfærøsotmikropladen. data udtrykt som foldændring fra basal. (E) Eksempel på kinetisk sporfremhævning, hvilke OCR-datamålinger (røde cirkler) der bruges til at beregne responsen for hver mitokondriemodulator for data præsenteret i E. De viste data er fra individuelle brøndsvar. Forkortelse: OCR = iltforbrugshastighed. Klik her for at se en større version af denne figur.
Udvælgelseskriterierne for baggrunden er af stor betydning; anvendelsen af yderbrønde til baggrundskorrektion er ikke repræsentativ for alle mikropladebrønde, hvilket kan føre til, at der drages forkerte dataantagelser, og at der drages fejlagtige datakonklusioner på grund af kanteffekter på tværs af den sfæroide mikroplade. For at vurdere denne observation blev MCF-7 sfæroider brugt til at sammenligne assaykorrektionsprocedurerne for at udlede OCR-værdier som reaktion på tilføjelsen af en køretøjskontrol, oligomycin, BAM15 eller rotenon-antimycin A (figur 8). Alle respiratoriske forbindelser gav de forventede kinetiske OCR-profiler for de udvalgte forbindelser, hvilket afslørede en gennemsnitlig stabil basal respirationshastighed på 20-30 pmol / min / brønd (figur 8A). Hvor assaydataene blev analyseret ved hjælp af de yderste brønde til baggrundstemperaturkorrektion (A1, A12, H1 og H12), var værdierne afsløret for OCR efter tilsætning af respiratoriske forbindelser imidlertid særligt lave; OCR gav negative værdier for rotenon-antimycin A. Som svar på disse observationer blev alternativ analyse udført ved hjælp af en række tomme brønde, tilfældigt fordelt over den sfæroide mikroplade, som baggrundstemperaturkorrektionsbrønde (figur 8B). Hvor der blev anvendt alternativ baggrundskorrektion, var alle relative sammensatte virkninger på OCR de samme i begge analysesæt; de absolutte OCR-værdier steg dog med ca. 10 pmol/min/brønd (figur 8). Disse data fremhæver effekten og vigtigheden af baggrundstemperaturkorrektion på sfæroid assaymikroplader og understreger vigtigheden af brugeroptimering til XF-analyse.

Figur 8: Tilfældigt valg af brønde til baggrundskorrektion for at forbedre kontrollen for temperaturgradienter på tværs af den sfæroide assaymikroplade. OCR-data ekstrapoleret fra figur 2A ved hjælp af anbefalede brønde til baggrundskorrektion (A) versus tilfældigt tildelte brønde til baggrundskorrektion (B). Forkortelse: OCR = iltforbrugshastighed. Klik her for at se en større version af denne figur.
I modsætning til cellemonolag repræsenterer sfæroider en heterogen aggregering af celler i et 3D-rum og kræver derfor grundig overvejelse med hensyn til analyse, især når disse data normaliseres. Dette papir præsenterer tre tilgange til normalisering af XF-data erhvervet fra MCF-7 sfæroider (figur 9). Når den ikke er normaliseret, korrelerer OCR positivt (R2 = 0,98) med sfæroidstørrelse (bestemt ved indledende cellesåningstæthed) signifikant sammenlignet statistisk med Pearson-korrelationskoefficient, P = 0,0057 (figur 9A). Dette lineære forhold sænkes, når OCR normaliseres til den oprindelige cellesåningstæthed (R2 = 0,78) og korrelerer ikke længere signifikant med sfæroidstørrelse (P = 0,117, figur 9B). Dette er også tilfældet, når det normaliseres til sfæroidvolumen (R2 = 0, 77; Pearson korrelationskoefficient P = 0,120, figur 9C) og nukleart dsDNA-indhold (R2 = 0,58; Pearson korrelationskoefficient P = 0,233, figur 9D). Disse data fremhæver vigtigheden af at normalisere XF-data, når man undersøger mitokondriemetabolismen af sfæroider, især hvis de er af forskellig størrelse.

Figur 9: Normalisering af ekstracellulære fluxdata erhvervet fra cellulære sfæroider. (A) Rå OCR-data blev opnået fra MCF-7 dyrket over 3 dage og plottet ved hjælp af Pearsons model for at opnå en korrelationskoefficient mellem sfæroid seeding densitet og OCR; P-værdi sat til 0,05. (B) Rå OCR-data blev normaliseret mod initial sfæroid seeding densitet; C) MCF-7 sfæroidvolumen opnået ved mikroskopiplanimetri og (D) nukleart ds DNA-indhold sammenlignet ved hjælp af Pearsons korrelationskoefficient. Forkortelser: OCR = iltforbrugshastighed; ds DNA = dobbeltstrenget DNA. Klik her for at se en større version af denne figur.
| Celle linje | Frøtæthed (brønd) | Sfæroid vækst (dage) | Endeligt sfæroidvolumen (μM3) | Basal OCR (pmolO2/min/brønd) | Følsomhed for basal OCR opfyldt (JA/NEJ) |
| SKOV | 1000 | 5 | 9,52E+06 | 28 ± 3,5 | Ja |
| SKOV | 2000 | 5 | 2.38E+07 | 26 ± 1.4 | Ja |
| SKOV | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | Ja |
| SKOV | 8000 | 5 | 1.11E+08 | 50 ± 7,9 | Ja |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0,7 | Nej |
| HepG2 | 2000 | 5 | 2,88E+07 | 23 ± 1,8 | Ja |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1,7 | Ja |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2,8 | Ja |
| A549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Ja |
| A549 | 2000 | 5 | 3,57E+07 | 41 ± 1,6 | Ja |
| A549 | 4000 | 5 | 6,93E+07 | 53 ± 7.2 | Ja |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8,4 | Ja |
| McF-7 | 1000 | 3 | 1.60E+07 | 29 ± 0,8 | Ja |
| McF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1,7 | Ja |
| McF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Ja |
| McF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2,9 | Ja |
Tabel 4: Optimerede parametre til bestemmelse af basale OCR-målinger i enkelte 3D-sfæroider. Forkortelse: OCR = iltforbrugshastighed.
Supplerende fil 1: Analyse af sfæroid størrelse og volumen. Klik her for at downloade denne fil.
Supplerende fil 2: Kvantificering af dobbeltstrenget DNA fra sfæroider i sfæroidmikropladen. Klik her for at downloade denne fil.
Supplerende fil 3: Anbefalinger til antallet af replikater, der kræves for at opnå pålidelige XF-analysedatasæt. Klik her for at downloade denne fil.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vigtigste resultater og resultater
Dette papir giver en detaljeret protokol til sonde mitokondrie energimetabolisme af enkelt 3D-sfæroider ved hjælp af en række kræftafledte cellelinjer med XFe96 XF Analyzer. En metode er udviklet og beskrevet til hurtig dyrkning af A549, HepG2 / C3A, MCF7 og SK-OV-3 cellulære sfæroider ved hjælp af celleafvisende teknologier til tvungen aggregering. Denne protokol omhandler mange overvejelser om sondering af sfæroidmetabolisme med XF-teknologi, herunder (1) optimering af sfæroidkulturprotokoller og håndtering og overførsel af sfæroider til specifikke sfæroidassaymikroplader fra teknologiproducenten fra deres originale dyrkningsbeholdere; 2) koncentrationen af respiratoriske forbindelser, der skal anvendes, og tidsafhængigheden af sammensat penetration 3) de injektionsstrategier, der skal anvendes og (4) måder at normalisere data mellem eksperimentelle grupper. Alle disse overvejelser er blevet undersøgt i det aktuelle dokument og behandles nærmere nedenfor. Disse metoder præsenteres som forenklede tilgange til generering af konsistente metaboliske iltfluxdata ved hjælp af enkelte 3D-sfæroider med XFe96 Flux-analysatoren. Denne eksperimentelle tilgang kan bruges som udgangspunkt og rubrik til brug i andre sfæroidmodeller, der let implementeres inden for en grundlæggende laboratorieindstilling.
Overvejelser
Sfæroid vækst, størrelse og følsomhed af XF-teknologi
For at etablere reproducerbare data med XF-teknologi er det vigtigt at karakterisere og optimere analysen for den specifikke model. Denne tilgang er relativt enkel i et grundlæggende monolag af celler; Dette giver dog yderligere udfordringer ved dyrkning af celler som 3D-sfæroider. Under de eksperimenter, der blev præsenteret her, blev RPMI-medium fra producenten suppleret ved brug. Mens det bemærkes, at nogle cellelinjer, nemlig HepG2 / C3A, blev dyrket i DMEM-vækstmedium, havde substitution med RPMI-DMEM-formuleringer begrænset indflydelse på XF-analyse under disse relativt korte assays (~ 3-5 timer). Formuleringen af de to medier er meget ens, og brugerne kunne 'tune' Seahorse RPMI-medier til at matche matrixen af deres cellekulturmedier gennem tilskud, f.eks. øget glukose, yderligere tilsætning af kulhydratkilder. Kritisk for den endelige formulering af alle XF-buffere og -medier er fraværet af phenolrød, som sandsynligvis vil forstyrre de fluorescerende sonder i XF-sondepatronpladen, og natriumbicarbonat, hvilket vil føre til alkalinitet på grund af manglen på CO2 -buffering, der er til stede i cellekulturinkubatorer. Andre medier og buffere kan købes og/eller laves internt. For eksempel er Krebs Ringer HEPES buffer en simpel buffer, der kan bruges til at vurdere respiration i mange forskellige celler, herunder sfæroidmodeller. Brugere af XF-assays bør dog bemærke, at en ændring i medium/buffer og dets tilskud kan ændre dets samlede bufferkapacitet. Dette er især bekymrende, når brugere kan være interesserede i at måle ECAR, hvor substratets bufferfaktor skal vurderes for at muliggøre ECAR-transformation til protonudstrømningshastighed (PER).
Da cellulær OCR målt ved XF-teknologi er proportional med celletætheden, når celletallet i brønden er inden for systemets følsomhed, var det vigtigt at undersøge dette forhold ved hjælp af enkelte 3D-sfæroider. Ved at undersøge OCR af enkelte 3D-sfæroider dyrket fra 4 forskellige kræftcellelinjer podet ved tætheder på 1.000, 2.000, 4.000 eller 8.000 celler pr. Brønd, viser vi, at XFe96-analysatoren er følsom nok til at opfange ændringer i hastigheden af mitokondrie respiration mellem 3D-sfæroider dyrket fra forskellige cellesåningstætheder (figur 3). Vi viser, at det optimale område af cellesåningstæthed og dermed sfæroidvolumen til dannelse af 3D-sfæroider til sondering af OCR varierer afhængigt af celletype. Dette vises ved det lineære forhold mellem OCR og frøtæthed eller sfæroidvolumen (figur 3). For A549- og HepG2/C3A-celler var den optimale såtæthed for OCR-følsomhed mellem 1.000 og 8.000 celler/brønd; det var 2.000-8.000 celler/brønd for MCF-7 og 4.000-8.000 celler/brønd for SK-OV-3 celler. Disse data viser, at optimering af sfæroid størrelse er af særlig betydning ved vurdering af OCR ved hjælp af XF-teknologi.
Overvejelser om minimale og maksimale sfæroidvolumener og basal OCR
Generelt vil der altid være minimums- og maksimumstærskler for målbare OCR-parametre, der anbefales af fremstillingen til disse eksperimenter. For XFe96-analysatoren er basal OCR mellem 20 pmol O2/min/brønd og 200 pmol O2/min/brønd henholdsvis nedre og øvre grænser. Dette er tilfældet med monolagsceller og sfæroider, og hvor den eksperimentelle model sidder inden for dette dynamiske OCR-område vil afhænge af mængden af tilgængeligt biologisk materiale, f.eks. antallet af celler som et monolag eller størrelsen af sfæroider. Se tabel 4 for et eksempel på, hvordan OCR-tærskler blev opnået ved hjælp af de sfæroidmodeller, der anvendes her. Det kan være klogt at kontrollere iltniveauet i brønden, for hvilken disse data også er tilgængelige fra disse målinger som niveaudata. Dette bør ses rutinemæssigt fra hvert eksperiment til kvalitetskontrolformål. Hvis der er iltsvind i brønden, vil dette fremgå af data. Hvis dette er tilfældet, kan det være nødvendigt at justere målecyklusserne i forsøget. for eksempel at øge blandingstrinnet, så iltniveauet i brønden genvindes inden den næste måleperiode inden for målecyklussen. Selvom det er muligt, har vi fundet dette meget usandsynligt for enkelt-sfæroideksperimenter ved hjælp af de beskrevne cellelinjer.
Valg af mitokondrie uncouplers til ekstracellulære flux assays
Protonionophorer, såsom carbonylcyanid 4-(trifluoromethoxy) phenylhydrazon (FCCP)21, carbonylcyanid m-chlorphenylhydrazon (CCCP)22 eller BAM1523, er potente småmolekylekemikalier, der er i stand til at forstyrre den elektrokemiske protongradient på tværs af mitokondriemembraner, hæmme produktionen af ATP og i sidste ende afkoble mitokondrie respiration24 . Der udvikles fortsat nye små molekyler til disse formål, navnlig til behandling af stofskiftesygdomme 25,26,27; se to fremragende anmeldelser28,29. Omvendt har afkobling af oxidativ respiration været forbundet med uønsket off-target toksicitet30. Inden for in vitro-cellulære assays depolariserer molekylet FCCP imidlertid mitokondriemembranpotentiale og udøver off-target effekter såsom plasmamembrandepolarisering, der forstyrrer NA + ionflux31; interferens med cellulær proteinbehandling32 og endda inducerende cellulær ældning33. BAM15 blev oprindeligt introduceret i 2013 som en mitokondrie uncoupler med minimal indflydelse på plasmamembraner23, med protonoforisk aktivitet i mikromolarområdet i hele celler og nanomolær rækkevidde i isolerede mitokondrier 23,34.
I betragtning af styrken af FCCP på plasmamembrandepolarisering er BAM15 en mere pålidelig protonofor til afkobling af respiration i intakte hele celler i ekstracellulære fluxassays. Selvom FCCP og dets modstykke, CCCP, er blevet brugt i over 50 år til at analysere maksimal respiratorisk kapacitet og fortsat anvendes bredt i XF-undersøgelser, undervurderer brugen af disse små molekyler ofte mitokondrie og cellulær metabolisk kapacitet. Dette er delvist forbundet med, hvorfor så mange publikationer, der bruger XF-teknologi, falder i fælden med at rapportere negative ekstra respiratoriske kapaciteter eller undervurdere ægte mitokondrie respiratoriske kapaciteter, når FCCP anvendes. Den ekstra styrke af FCCP i intakte celler og væv fører ofte til kompromitteret mitokondriefunktion, og celler kan kæmpe for at fungere hensigtsmæssigt for at opretholde en maksimal åndedrætskapacitet på tværs af flere målecyklusser efter deres tilsætning, selv ved meget lave koncentrationer35. Derfor kan cellernes respons på FCCP i mange undersøgelser findes at falde efter den indledende målecyklusperiode. Mens FCCP rutinemæssigt er blevet anvendt til XF-analyse, anvendes BAM15 fortrinsvis i tilfælde, der involverer hele celler eller sfæroidmodeller, da det kan opretholde en maksimal åndedrætskapacitet i fuldt depolariserede mitokondrier i koncentrationer så høje som 10 μM3. Desuden inducerer BAM15 virkninger på ekstracellulær forsuring, som falder sammen med næringsstofoxidation gennem hydrering af CO2 til dannelse af HCO3- og H + i større grad end FCCP3. Ikke desto mindre bør nogen af disse afkoblinger i tilfælde af isolerede mitokondrier og permeabiliserede celler fungere såvel som BAM15 til mitokondrieafkobling, hvis de titreres i den korrekte koncentration.
Kinetik af sammensat penetration og assay cykling
Koncentrationerne, penetrationen og kinetiske profiler af kemiske forbindelser, der bruges til at udføre en typisk MitoStress-test med 3D-sfæroider ved hjælp af XF-analysatoren, er mere komplekse at adressere. I betragtning af at sfæroider præsenterer 3D-struktur, er molekylernes indtrængning over sfæroidens diameter en uendeligt mere kompleks proces end på tværs af cellemonolag. For eksempel blev den kinetiske penetration og derfor følsomheden over for den kemoterapeutiske sorafenib bestemt af sfæroid alder og derfor størrelse i en HepG2 sfæroid model36. Småmolekylekemikaliers (f.eks. lægemidler, nanopartiklers) evne til at nå et biologisk mål afhænger af flere underliggende faktorer, herunder kompleksiteten af det system, der skal gennemtrænges dynamisk og diffunderes gennem 37,38. Dette gælder især for lægemidler rettet mod tumorvæv39. Svarende til tumormålretning i forbindelse med en 3D-sfæroid, størrelse, kompaktitet og andre fænotypiske reaktioner, såsom ekspression af lægemiddeltransportproteiner, kan styre penetrationstiden og koncentrationen af en forbindelse, der kræves for at fremkalde et biologisk respons.
I denne protokol behandlede vi problemet omkring penetrationstid og koncentration af små molekyler som reaktion på ATP-syntasehæmmeren oligomycin, protonoforen og mitokondrie-uncoupler BAM15 og kombinationen af Complex I- og Complex III-hæmmerne rotenon og antimycin A. Ved at undersøge OCR for enkelte MCF-7-sfæroider udsat for flere titreringer af disse almindelige respiratoriske forbindelser viser vi, at den optimale koncentration af hver forbindelse, der kræves for at inducere en steady-state respirationshastighed, falder inden for et lignende interval som for monolagsceller (figur 4). Det er vigtigt og forskelligt fra deres monolagsmodstykker, at det er vist, at forøgelse af antallet af målecyklusser mellem injektioner er nøglen til at opnå en steady state OCR i enkelt 3D-sfæroider. Disse data fremhæver vigtigheden af sammensat penetration og deres respektive kinetiske profiler, når man udforsker mitokondrie respiratoriske parametre for 3D-sfæroider ved hjælp af disse tilgange. Ved hjælp af sfæroidoptimeringsegenskaber, koncentrationer af forbindelser og målecyklustider informeret af dataene i figur 3, figur 4 og figur 5 blev der etableret en valideret MitoStress-test til undersøgelse af specifikke parametre for mitokondrie oxidativ metabolisme i en række kræftafledte 3D-sfæroider (figur 6). Af betydning, og ligesom nogle monolagscancercellelinjer40, blev den maksimale respiratoriske kapacitet (hastighed af ukoblet stimuleret respiration) af visse kræftafledte 3D-sfæroider hæmmet af oligomycin (figur 5). Specifikt viste 3D-sfæroider dyrket fra enten A549- eller MCF-7-celler en signifikant lavere maksimal respirationshastighed, når de blev afkoblet med BAM15 efter oligomycininjektion sammenlignet med at blive afkoblet af BAM15 uden oligomycin (figur 5F og figur 5H). I betragtning af at denne effekt kan være til stede i andre 3D-sfæroidkulturer, foreslår vi, at medmindre en tidligere valideret protokol anvendes, bør maksimal respiratorisk kapacitet i 3D-sfæroider estimeres uden oligomycin.
Samtidig indsamling af ECAR-data som et mål for glykolytisk flux i cellulære sfæroider
Som det typisk ses i litteraturen eller informationen fra teknologiproducenten, er den glykolytiske hastighed af sfæroider, målt som ECAR, en sekundær parameter, der kan fanges sammen med OCR. Beregning af ECAR alene er ikke en nyttig eller meningsfuld parameter i noget XF-eksperiment, da den ikke korrigeres for bufferkapaciteten af XF-assaybufferen eller tilsætning af mitokondrieforsuring, der stammer fra hydrering af CO2 tilHCO3- og H +. ECAR er først indsigtsfuld, når disse datakorrektioner er anvendt, hvorefter det bliver muligt at give mere nøjagtige konklusioner om glykolytisk flux. For at korrigere for bufferkapaciteten til at generere mere meningsfulde PER-data, skal man kende volumenet af mikrokammeret for den sfæroide mikroplade. Producenten har ikke været i stand til at levere et ægte volumen til dette med den sfæroide mikroplade, og derfor kan PER-data ikke bestemmes let. Selvom disse målinger kunne opnås empirisk, var dette uden for dette manuskripts anvendelsesområde. Men med de relevante korrektioner og kendskab til volumenet af mikrokammeret for en given sfæroidstørrelse til stede (f.eks. Opnåelse af et mål for sfæroidtæthed) i brønden, ville ECAR-data blive meningsfulde, og beregninger af glykolytisk PER kunne foretages. Derfor kunne XF-data derefter være mere informative til undersøgelse af glykolytisk og oxidativ metabolisme i sfæroider, men kun hvis disse parametre blev overvejet i dybden.
Sfæroid dannelse, håndtering, overførsel og bevægelse
Nogle cellelinjer er bedre egnet til dannelse af sfæroider end andre og danner muligvis slet ikke sfæroider, f.eks. danner MCF-7 ovariecancerceller41,42 meget cirkulære sfæroider sammenlignet med andre cellelinjer (figur 3). Som et andet eksempel har Capan-1 kræftceller i bugspytkirtlen vist sig at danne bedre sfæroider end Panc-1 eller BxPC343. Tilsvarende er hepatisk carcinomcellelinjer kendt for at have variable evner til at danne kompakte sfæroider 5,44 med en observeret ændring i fænotype, såsom forbedret lægemiddelmetabolisme eller produktion af albumin, som det er tilfældet for HepG2 versus HepG2 / C3A 9,45,46 eller HepaRG sfæroider 17,47,48 . Derfor bør brugerne optimere sfæroiddyrkningsteknikker i overensstemmelse hermed og udføre titreringseksperimenter for at bestemme optimal frøtæthed og dyrkningstidsforløb. Desuden har formuleringen og sammensætningen af assaymedier vist sig at påvirke sfæroidformuleringen, herunder tilsætning af methylcellulose, ofte tilsat til medier for at øge matrixviskositeten 43,49,50. Derfor bør den optimale cellemediumsammensætning bestemmes empirisk for alle anvendte cellelinjer.
Antallet af mediumudvekslinger gennem sfæroidkultur bestemmes af den anvendte cellelinje. Imidlertid er typisk halvvolumen medium udvekslinger hver 2-3 dage gældende i de fleste tilfælde for at genopbygge næringsstoffer. Vi brugte den tvungne aggregerede tilgang til at generere 3D-sfæroider ved hjælp af celleafvisende mikroplader fra kommercielt tilgængelige kilder til hurtig udvikling og implementering af sfæroidmodeller i XF-analyseundersøgelser. Alternative platforme kan dog være bedre egnet til at generere sfæroider fra andre celletyper, f.eks. hængende-drop eller matrix-indlejrede tilgange. I ressourcebegrænsede laboratorier kan brugerne ønske at se mod agarose-væskeoverlejringsteknikken til dannelse af celleafvisende mikropladeoverflader51,52 for betydeligt at reducere de økonomiske omkostninger ved indledende sfæroid metodeudviklingstrin. Bevægelsen af sfæroider mellem kulturbeholdere er nødvendig for at udføre XF-analyse og andre nedstrøms assays. Nem overførsel dikteres typisk af sfæroid størrelse og samlet tæthed. Vi anbefaler at bruge en P200- eller P1000-pipettespids med bred åbning for at opretholde sfæroid integritet; Mindre borede pipettespidser risikerer mekanisk forstyrrelse af sfæroiden, som kan købes kommercielt eller med omhu fremstilles ved blot at trimme enden af pipettespidsen for at øge åbningen. Denne fremgangsmåde kan dog være tilbøjelig til at indføre pelsning til plasten omkring enden af spidsen, hvilket kan forårsage mekanisk forstyrrelse under håndtering. Brugen af baggrundsbelysning eller lysboks er også nyttig til sfæroid håndtering og observation under et dissektionsmikroskop som et vigtigt skridt for at sikre en vellykket overførsel af sfæroider til sfæroid assaymikropladen. Desuden er den sfæroide position i brønden på en sfæroid assaymikroplade af særlig betydning og påvirker direkte OCR og sammensat effekt under en typisk MitoStress-test (figur 7), sandsynligvis på grund af forholdet mellem sfæroidpositionen og sensorsondens fluoroforer.
Baggrundskorrektion og temperaturreguleringsbrønde
Brugen af mikropladebaserede assays er en meget anvendt tilgang inden for flere forskningsområder; Deres anvendelse giver dog flere praktiske udfordringer. Som det er tilfældet i andre eksperimentelle tilgange, især dem, der bruger 96 (eller større) array-format, kan mikropladegeometri og positionering påvirke temperatur- og gasudvekslingsgradienter over pladen over tid, ofte omtalt som 'kanteffekter'53,54. Vi fandt, at det samme var tilfældet for den sfæroide assaymikroplade. I henhold til producentens retningslinjer og protokoller er de yderste hjørnebrønde: A1, A12, H1 og H12 altid udpeget som baggrundskorrektions- og temperaturreguleringsbrønde til XFe96-analysatoren. Omvendt er A1 og D6 med 24-brønds array-formatet betegnet som kontrolbrønde sammen med to andre brønde jævnt fordelt over midten af pladen ved B4 og C3. Ved udførelse af XF-sfæroidanalyse fandt vi signifikant afvigelse i data, der oprindeligt blev indsamlet ved hjælp af producentens vejledning. Dette var på trods af, at der blev taget de nødvendige skridt til at sikre, at der blev foretaget en forvikling af temperatur og CO2 -indhold, inden dataindsamlingen begyndte, hvilket ofte gav negative værdier for OCR efter injektion af visse respiratoriske hæmmere (figur 8).
Vi fandt, at disse observationer sandsynligvis skyldtes kanteffekter på tværs af den sfæroide assaymikroplade. I figur 8 fandt vi, at omfordeling af baggrundskontrolbrønde på tværs af mikropladen, XF-data blev justeret ca. 2 gange. To mest sandsynlige årsager er (1) på grund af fordampningseffekter ved kantbrøndene, hvilket resulterer i et mindre samlet volumenkammer for XFe96-sonden at prøve fra, og (2) fra utilstrækkelige temperaturækvilibrationer mellem de brønde, der er udpeget til baggrundskorrektion og prøvebrønde, hvilket resulterer i datasæt, der enten maskerer eller overpuster OCR. For at undgå sådanne resultater anbefales det derfor, især i forbindelse med sfæroidanalyse, at brugerne omfordeler brønde, der er udpeget til baggrundskorrektion, over hele sfæroide assaymikropladen og tager de nødvendige skridt til at forligebinde deres assay, inden de får XF-data.
Normalisering af data
Ud over at give en detaljeret protokol til sondering af mitokondrieenergimetabolisme af enkelt 3D-sfæroider med XF-teknologi, præsenterer dette papir også mulige måder at normalisere mitokondrie respiratoriske hastighedsdata opnået med 3D-sfæroider. Ved hjælp af respirationsfrekvensdata opnået med MCF-7 sfæroider dyrket ved forskellige cellesåningstætheder (figur 3) præsenterer vi basale mitokondrie respiratoriske hastigheder fra MCF-7 sfæroider af stigende størrelse og diameter, når de normaliseres til indledende cellesåningstæthed, sfæroidvolumen og dsDNA-indhold (figur 9). Den passende normaliseringsmetode er afgørende for den nøjagtige fortolkning af XF-datasæt, især når man sammenligner in vitro 3D-sfæroidmodeller og forskellige celletyper. Dårlig normalisering kan føre til fejlagtige resultater, der simpelthen ikke kan sammenlignes mellem datasæt. Proteinindhold foretrækkes ikke til normalisering af sfæroid XF-data, da forbehandlinger kan påvirke proteinsyntesehastigheder uden signifikant effekt på respirationsfrekvensen. Desuden kan betydelige, inkonsekvente mængder protein binde sig til sfæroide mikroplader på cellelysis, hvilket introducerer variation i proteinindholdet mellem brønde. Dette kan være yderligere kompliceret i XF-analyser ved hjælp af sfæroider eller ikke-tilstødende celler, der kræver biomolekylære lim at binde til, som kan indeholde protein.
I modsætning til intracellulært proteinindhold er kerne-DNA-indhold uafhængigt af celletype og er proportionalt med cellenummer (figur 9D) - en mere præcis og mindre tidskrævende tilgang end opdeling af sfæroider til kvantificering af cellenummer. Omvendt fandt Yepéz et al.55, der gennemførte XF-analyser i monolag af fibroblasterceller, at normalisering af XF-data til celleantal introducerede større spredning af data end før normalisering. Kerne-DNA-indhold er uafhængigt af differentieret tilstand eller fænotype og er derfor mere nøjagtigt til normalisering af sfæroiddata i XF-assays end proteinindhold. DNA-indhold har også været en gennemprøvet strategi til analyse af andre metabolisme-forbundne datasæt56. Det er dog vigtigt at bemærke, at kerne-DNA-indhold kvantificeres fra alle celler, der er til stede i sfæroidet; derfor anbefales normalisering af DNA-indhold ikke til XF-datasæt, hvor sfæroider gennemgår behandlinger, der kan resultere i betydeligt tab af cellelevedygtighed. For sådanne datasæt foretrækkes normalisering til cellelevedygtighed, hvis det er muligt, eller dataene kan baseline-korrigeres til basal respiration.
Brug af ekstra åndedrætskapacitet som eksempel på vigtigheden af datanormalisering
Ekstra respiratorisk kapacitet er et mål for hastigheden af maksimal mitokondrie respiratorisk kapacitet minus basal mitokondrie respirationsfrekvens (figur 6). Problemet med rapportering af data af denne type som en hastighed, dvs. pmolO2 / min / godt inden for visse eksperimenter, er imidlertid, at dataene er uden normalisering. Selvom man normaliserer sfæroiddata til celletæthed / DNA-indhold, udelukker dette ofte nøgleparameteren, der skal normaliseres for mitokondrietæthed i cellerne. I betragtning af at en ændring i mitokondrietæthed vil føre til en proportional ændring i basal og maksimal respiration, vil ledig kapacitet også stige. For eksempel, hvis sfæroid OCRbasal er 200 og OCRmax er 400, rapporteres ledig kapacitet som 200; hvis OCRbasal er 100 og OCRmax, så er ledig kapacitet også 100; men i procent er de begge 50% af maksimal (eller 100% af basal). Derfor ændres den uudnyttede kapacitet ikke mellem disse to eksempler på trods af forskelle i satser på 200 og 100, når de beregnes som pmols O2/min/brønd. Internt normaliserede værdier er mere pålidelige og indsigtsfulde for at gøre XF-data mere sammenlignelige på tværs af undersøgelser og projekter. For at gøre dette for ekstra åndedrætskapacitet har vi valgt at præsentere dette som en procentdel af maksimal respiration i stedet for en absolut sats. Dette kan også præsenteres som en procentdel af basal respiration. Dette ville være tilfældet, hvis man arbejder med celler eller sfæroider. I betragtning af at placeringen af sfæroidet i mikrobrøndpladen kan ændre den absolutte OCR, men ikke de relative ændringer med inhibitorer eller frakoblinger, er det vigtigere at se på internt normaliserede reaktioner i sfæroider som foldændring eller procentdele.
De sfæroidmodeller, der genereres her, præsenterer en række celletyper og arkitektur, der ikke kan fanges i klassiske 2D-modeller. Disse omfatter heterogent, rumligt arrangement af celler i tre dimensioner, forbedrede celle-cellekontakter (f.eks. Dannelse af mellemrumskryds og ekstracellulære matricer) og biokemiske gradienter over sfæroiddiameteren (f.eks. pH-gradienter, iltdiffusionsadgang til næringsstoffer). Brug af ekstracellulær flux til at studere in vitro sfæroidbiologi kan gøre det muligt at identificere optimale mål for lægemiddelterapier gennem metaboliske forstyrrelser. Disse kan ekstrapoleres fra in vitro-sfæroider til in vivo-tumorer og identificere veje, der kan målrette sfæroide tumormetabolisme, f.eks. kulhydratudnyttelse under sfæroidvækst. Terapeutiske modaliteter kan være effektive til at målrette sfæroider i tidlige vækstfaser, men viser sig mindre effektive i de senere faser af sfæroidvækst, efterhånden som metabolisk netværkskompleksitet modnes. Afslutningsvis vil udviklingen af 3D-cellekulturmodeller og sofistikerede analyseteknologier inden for biologisk forskning fortsat være et dynamisk og hurtigt skiftende felt med uovertruffen potentiale. Ekstracellulær fluxanalyse af in vitro-cellekultursfæroider kunne anvendes som en banebrydende forskningsmetode til at fremme forskningsresultater, der kunne ekstrapoleres for at forstå humanrelevant biologi bedre, reducere brugen af dyremodeller i forskning og forbedre patientcentreret forskning.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingen interessekonflikter at erklære.
Acknowledgments
N.J.C blev støttet af en BBSRC MIBTP CASE Award med Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Kræftforskning nummer 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
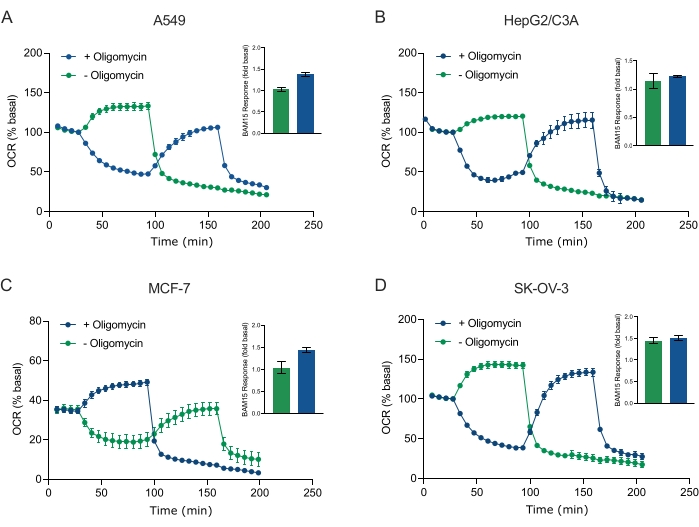
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
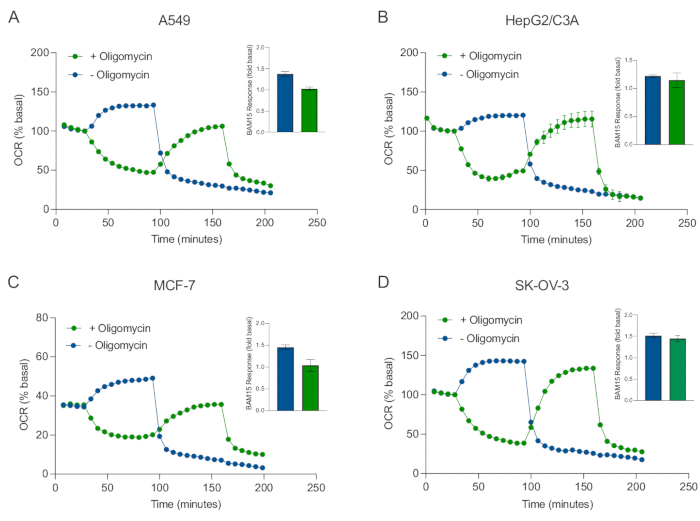
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
