

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Эти протоколы помогут пользователям исследовать энергетический метаболизм митохондрий в 3D-сфероидах, полученных из клеточных линий рака, с использованием анализа внеклеточного потока Seahorse.
Abstract
Трехмерные (3D) клеточные агрегаты, называемые сфероидами, стали авангардом клеточной культуры in vitro в последние годы. В отличие от культивирования клеток в виде двумерных одноклеточных монослоев (2D-культура), сфероидная клеточная культура способствует, регулирует и поддерживает физиологическую клеточную архитектуру и характеристики, которые существуют in vivo, включая экспрессию белков внеклеточного матрикса, клеточную сигнализацию, экспрессию генов, производство белка, дифференцировку и пролиферацию. Важность 3D-культуры была признана во многих областях исследований, включая онкологию, диабет, биологию стволовых клеток и тканевую инженерию. За последнее десятилетие были разработаны усовершенствованные методы получения сфероидов и оценки их метаболической функции и судьбы.
Анализаторы внеклеточного потока (XF) использовались для изучения митохондриальной функции в 3D-микротизюсах, таких как сфероиды, с использованием либо пластины захвата островков XF24, либо сфероидной микропластины XFe96. Однако отдельные протоколы и оптимизация зондирования митохондриального энергетического обмена у сфероидов с использованием технологии XF подробно не описаны. В данной статье представлены подробные протоколы зондирования митохондриального энергетического обмена в одиночных 3D-сфероидах с использованием сфероидных микропластин с анализатором XFe96 XF. Используя различные линии раковых клеток, технология XF способна различать клеточное дыхание в 3D-сфероидах не только разных размеров, но и разных объемов, количества клеток, содержания и типа ДНК.
Оптимальные концентрации митохондриальных эффекторных соединений олигомицина, БАМ15, ротенона и антимицина А используются для исследования специфических параметров митохондриального энергетического метаболизма в 3D-сфероидах. В этой статье также обсуждаются методы нормализации данных, полученных от сфероидов, и рассматриваются многие соображения, которые следует учитывать при изучении сфероидного метаболизма с использованием технологии XF. Этот протокол поможет стимулировать исследования в передовых сфероидных моделях in vitro .
Introduction
Достижения в области моделей in vitro в биологических исследованиях быстро прогрессировали за последние 20 лет. Такие модели теперь включают модальности «орган на чипе», органоиды и 3D-сфероиды микротизю, все из которых стали общим фокусом для улучшения трансляции между исследованиями in vitro и in vivo. Использование передовых моделей in vitro, особенно сфероидов, охватывает несколько областей исследований, включая тканевую инженерию, исследования стволовых клеток, рак и биологию заболеваний 1,2,3,4,5,6,7, и тестирование безопасности, включая генетическую токсикологию 8,9,10, токсикологию наноматериалов11, 12,13,14, и тестирование безопасности и эффективности лекарств 8,15,16,17,18,19.
Нормальная морфология клеток имеет решающее значение для биологического фенотипа и активности. Культивирование клеток в 3D-сфероиды микротизуса позволяет клеткам принимать морфологию, фенотипическую функцию и архитектуру, более похожую на ту, что наблюдается in vivo , но ее трудно захватить с помощью классических методов монослойной клеточной культуры. Как in vivo , так и in vitro клеточная функция напрямую зависит от клеточной микросреды, которая не ограничивается клеточной связью и программированием (например, образования клеточно-клеточных переходов, возможности формирования клеточных ниш); воздействие на клетки гормонов и факторов роста в непосредственной среде (например, воздействие клеточных цитокинов как часть воспалительной реакции); состав физических и химических матриц (например, выращиваются ли клетки в пластической культуре жесткой ткани или в эластичной тканевой среде); и, самое главное, как на клеточный метаболизм влияют питание и доступ к кислороду, а также переработка метаболических отходов, таких как молочная кислота.
Анализ метаболических потоков является мощным способом изучения клеточного метаболизма в определенных системах in vitro . В частности, технология XF позволяет анализировать живые изменения в клеточной биоэнергетике интактных клеток и тканей в режиме реального времени. Учитывая, что многие внутриклеточные метаболические события происходят в течение порядка секунд до минут, функциональные подходы в реальном времени имеют первостепенное значение для понимания изменений клеточного метаболического потока в реальном времени в неповрежденных клетках и тканях in vitro.
В данной статье представлены протоколы культивирования раковых клеточных линий A549 (аденокарцинома легких), HepG2/C3A (гепатоцеллюлярная карцинома), MCF-7 (аденокарцинома молочной железы) и SK-OV-3 (аденокарцинома яичников) в виде 3D-сфероидных моделей in vitro с использованием подходов принудительной агрегации (рисунок 1). В нем также (i) подробно описывается, как исследовать митохондриальный энергетический метаболизм одиночных 3D-сфероидов с помощью анализатора Agilent XFe96 XF, (ii) выделяются способы оптимизации анализов XF с использованием одиночных 3D-сфероидов и (iii) обсуждаются важные соображения и ограничения зондирования метаболизма 3D-сфероидов с использованием этого подхода. Самое главное, в этой статье описывается, как собираются наборы данных, которые позволяют рассчитать скорость потребления кислорода (OCR) для определения окислительного фосфорилирования и, следовательно, митохондриальной функции в клеточных сфероидах. Хотя это не анализируется для этого протокола, скорость внеклеточного подкисления (ECAR) является еще одним параметром, который измеряется вместе с данными OCR в экспериментах XF. Однако ECAR часто плохо или неправильно интерпретируется из наборов данных XF. Мы предоставляем комментарий относительно ограничений расчета ECAR в соответствии с основными подходами производителя технологии.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Рисунок 1: Графический рабочий процесс для генерации клеточных сфероидов, анализа внеклеточного потока и последующих анализов. Четыре линии раковых клеток были селективно культивированы как монослои (А), отделены от колб тканевых культур и засеяны в сверхнизкие прикрепленные 96-луночные микропластины с образованием сфероидов (В). A549 Легочная карцинома, HepGG2/C3A печеночная карцинома, SK-OV-3 аденокарцинома яичников и MCF-7 клетки карциномы молочной железы были посеяны через 1 × 103-8 × 103 клеток / хорошо и выращены до 7 дней для формирования одиночных сфероидов и оптимизации плотности посева сфероидов и времени культивирования путем непрерывного наблюдения и планиметрических измерений. После образования одиночные сфероиды промывали в безсыворочную XF-среду и тщательно сеяли в сфероидные микропластины, предварительно покрытые поли-D-лизином (C). Сфероиды подвергали внеклеточному анализу потока с использованием анализатора XFe96 с использованием нескольких протоколов для решения: (1) оптимального размера сфероида для ответа базального митохондриального дыхания; (2) оптимизированное титрование митохондриальных респираторных ингибиторов; (3) Оптимизация размещения сфероидов в микропластинчатых скважинах. (D) Для нормализации данных и других последующих анализов in vitro использовались анализы после XF, фазоконтрастная микроскопия и количественная оценка сфероидной ДНК. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
1. Культивирование линий раковых клеток в виде 3D сфероидов in vitro
| Клеточная линия | Описание | Культуральная среда | Источник |
| А549 | Клеточная линия карциномы легких | РПИ 1640 | Европейская коллекция аутентифицированных клеточных культур (ECACC) |
| Пируват натрия (1 мМ) | |||
| Пенициллин-Стрептомицин - (100 Ед/мл – 100 мг/мл) | |||
| 10 % (об/об) FBS | |||
| HepG2/C3A | Клеточная линия печеночной карциномы, клональное производное родительской клеточной линии HepG2 | ДМЭМ | Американская коллекция культуры тканей (ATCC) |
| Пенициллин-Стрептомицин - (100 Ед/мл – 100 мг/мл) | |||
| 10 % (об/об) FBS | |||
| МКФ7 | Клеточная линия аденокарциномы молочной железы | РПИ 1640 | Европейская коллекция аутентифицированных клеточных культур (ECACC) |
| Пируват натрия (1 мМ) | |||
| Пенициллин-Стрептомицин - (100 Ед/мл – 100 мг/мл) | |||
| 10 % (об/об) FBS | |||
| СК-ОВ-3 | Клеточная линия аденокарциномы яичников | РПИ 1640 | Европейская коллекция аутентифицированных клеточных культур (ECACC) |
| Пируват натрия (1 мМ) | |||
| Пенициллин-Стрептомицин - (100 Ед/мл – 100 мг/мл) | |||
| 10 % (об/об) FBS | |||
| Компонент | Среда для анализа RPMI (конечный объем 50 мл) | ||
| Базовый средний | Agilent Seahorse XF RPMI, pH 7.4 | ||
| Глюкоза (1 М стерильного вещества) | 11 мМ (стоковый раствор 0,55 мл) | ||
| L-глютамин (стерильный материал 200 мМ) | 2 мМ (0,5 мл запасного раствора) | ||
| Пируват натрия (стерильный материал 100 мМ) | 1 мМ (0,5 мл запасного раствора) |
Таблица 1: Раковые клеточные линейные среды и композиции XF-сред.
- Культивируйте все клеточные линии с использованием стандартной асептической техники культивирования тканей и подтвердите, что они свободны от микоплазмы, используя подходящий набор для анализа.
- Культивируйте клеточные линии в колбах для культивирования тканей Т75 или эквиваленте, используя рекомендуемую среду (таблица 1). Культивируйте клеточные линии до 65-80% слияния и проходите их регулярно максимум до 25 проходов.
- Промыть колбы клеточной культуры дважды в модифицированном фосфатно-буферном физиологическом растворе Dulbecco (DBPS).
- Отделяют клетки от колб 3 мл клеточного диссоциационного реагента (см. Таблицу материалов) в течение 5 мин при 37 °C и подтверждают отслоение микроскопией.
- Аккуратно аспирировать отсоединенную клеточную суспензию, чтобы обеспечить одноклеточную суспензию и дезактивировать реагент диссоциации клеток 7 мл полной тканевой питательной среды.
- Соберите клетки путем центрифугирования при 300 × г в течение 5 мин, выбросьте супернатант и повторно сведите клетки в полную среду.
- Подсчитайте клетки с помощью гемоцитометра или автоматизированного счетчика клеток и титруйте до желаемой плотности клеток, необходимой для посева.
ПРИМЕЧАНИЕ: Для посева всей 96-луночной пластины при 100 мкл/лунке при 4 × 103 клетках/лунке клетки следует титровать до 4 × 104 клеток/мл в рекомендуемом объеме 12 мл. - Декантируйте клеточную суспензию в стерильный резервуар и дозируйте 100 мкл клеточной суспензии в каждую лунку клеточно-репеллентной микропластины с помощью многоканального пипеттера.
ПРИМЕЧАНИЕ: Только внутренние 60 лунок микропластины должны быть засеяны, а остальные заполнены DPBS. Это сформирует барьер испарения, обеспечит однородность сфероидов по всей пластине и минимизирует эффекты края пластины. - Центрифужные сфероидные микропластины при 300 × г в течение 15 мин для форсирования клеток в рыхлые агрегаты.
- Инкубируйте пластины при 37 °C, 5% CO2 в течение минимум 3 дней, чтобы обеспечить образование сфероидов.
- Выполняйте фазово-контрастную микроскопию с использованием стандартизированных лабораторных практик для мониторинга роста сфероидов. Пополняйте клеточную культуральную среду каждые 3 дня или два раза в неделю, выполняя обмен средой в половину объема.
2. Зондирование митохондриального энергетического метаболизма одиночных сфероидов с использованием технологии Extracellular Flux (XF)
- Подготовка к анализу (за день до этого)
- Проверьте жизнеспособность сфероида с помощью инвертированного светового микроскопа с фазовым контрастом при 4-кратном увеличении, чтобы обеспечить неповрежденную сфероидную структуру, морфологию и общую однородность между образцами.
- Увлажните картридж датчика.
- Аликвота ~20 мл калибранта в коническую трубку.
- Поместите коническую трубку, содержащую калибрант, в инкубатор без CO2 37 °C на ночь.
- Извлеките содержимое из набора для анализа.
- Извлеките картридж датчика из вспомогательной пластины и поместите его вверх ногами на столешницу рядом с вспомогательной пластиной.
- Пипетка 200 мкл стерильного ddH2Oв каждую скважину картриджной пластины датчика с помощью многоканальной пипетки P300.
- Поместите картридж датчика поверх вспомогательной пластины.
- Убедитесь, что уровень воды в каждой скважине достаточно высок, чтобы погрузить датчики.
- Перенесите собранный картридж датчика в инкубатор без CO2 37 °C и оставьте на ночь.
ПРИМЕЧАНИЕ: Этот этап может быть выполнен за 12-72 ч до начала анализа.
- Микропластина сфероидного анализа покрытия
- Используя асептические методы, добавляют 30 мкл/лунку стерильного раствора поли-D-лизина (0,1 мг/мл) к сфероидной микропластине и инкубируют ее в течение 30 мин при комнатной температуре.
- Аспирируйте раствор из каждой лунки сфероидной микропластины, переверните пластину и плотно постучите ее по папиросной бумаге, чтобы удалить любой остаточный раствор.
- Дважды промыть пластину 200 мкл/лунку стерильным ddH2O.
- После окончательной стирки переверните микропластинку и плотно постучите ее на папиросную бумагу, чтобы удалить остаточную воду.
- Дайте пластине высохнуть на воздухе в течение 30 минут перед использованием или хранением при температуре 4 °C для будущего использования.
ПРИМЕЧАНИЕ: Микропластина сфероидного анализа должна быть покрыта молекулярным клеем, чтобы гарантировать, что сфероиды закреплены в нижней части микропластины. Без молекулярного клея сфероиды могут смещаться и мешать результатам анализа. Другие молекулярные клеи также могут быть использованы в качестве альтернативы Poly-D-лизину для пластин предварительного покрытия. Предварительно покрытые пластины могут храниться при температуре 4 °C, но их следует оставить для выравнивания до комнатной температуры до начала анализа.
- Подготовка среды для анализа XF
- Подготовка среды XF RPMI, как описано в таблице 1, и стерильного фильтра со шприцевым фильтром 0,22 мкм
- Подготовка к анализу (за 1 ч до анализа)
- Предварительно прогрейте добавленную XF RPMI пробирную среду до 37 °C.
- Предварительно прогрейте микропластинку сфероидного анализа с покрытием в инкубаторе безСО2 37 °C или сухой ванне.
- Подготовьте картридж датчика.
- Извлеките коническую трубку, содержащую калибрант и картридж датчика, из воздушного инкубатора.
- Извлеките картридж датчика из вспомогательной пластины и поместите его вверх ногами на рабочую поверхность.
- Используя многоканальную пипетку P300, аспирируйте воду с опорной плиты и выбросьте ее.
- Залейте раствор калибранта в резервуар для стерильных реагентов и добавьте 200 мкл/лунку предварительного калибранта к вспомогательной пластине с помощью многоканальной пипетки P300.
- Возьмите картридж датчика и поместите его обратно на верхнюю часть вспомогательной пластины, чтобы датчики были хорошо погружены в калибр.
- Перенесите собранный картридж датчика обратно в инкубатор без CO2 при 37 °C до готовности к загрузке растворов для впрыска в порт.
- Промыть сфероиды пробирной средой.
- Извлеките пластину для культуры сфероидов из инкубатора 37 °C, 5% CO2 и наблюдайте за сфероидами под микроскопом, чтобы обеспечить их целостность перед этапами переноса сфероидов.
- Загрузите все скважины сфероидной пластины 180 мкл/лунку предварительной пробирной среды, включая любые скважины фоновой коррекции.
- Частично заполните чашку Петри размером 7 см 3 мл пробирной среды.
- Используя многоканальную пипетку, нагруженную широкими наконечниками отверстий пипетки, переведите сфероиды с 96-луночной культуральной пластины в чашки Петри 7 см, установив пипетку на объем аспирации 10-50 мкл.
- Семена сфероидов в предварительно покрытую сфероидную микропластину.
- Используя микроскоп для рассечения и аппарат лайтбокса, перенесите сфероиды из чашки Петри на микропластину сфероидного анализа, как описано ниже.
- Установите объем одноканального пипеттера, оснащенного широким наконечником отверстия пипетки, на 20 мкл и тщательно аспирировать один сфероид. Поместите наконечник непосредственно в центр каждой лунки микропластины сфероидного анализа и позвольте гравитации элюировать один сфероид в центр каждой лунки, т. е. не выталкивайте какую-либо среду из кончика пипетки и позвольте капиллярному действию вывести сфероид из кончика пипетки. Для подтверждения элюирования содержимое пипетатора можно пипетировать обратно в 7 см чашку Петри под микроскопом.
ПРИМЕЧАНИЕ: Гравитационное элюирование одного сфероида обычно занимает 15-30 с в зависимости от размера/плотности сфероида. В течение этого времени пипетку не следует снимать. Любые лунки фоновой коррекции должны быть свободны от сфероидов и содержать только пробирную среду. Под микроскопом подтвердите положение каждого сфероида. Каждый сфероид в идеале должен быть расположен в центре каждой скважины. - После того, как все сфероиды были перенесены на микропластину сфероидного анализа, перенесите пластину в инкубатор без CO2 при 37 °C в течение как минимум 1 ч до анализа.
- Установите объем одноканального пипеттера, оснащенного широким наконечником отверстия пипетки, на 20 мкл и тщательно аспирировать один сфероид. Поместите наконечник непосредственно в центр каждой лунки микропластины сфероидного анализа и позвольте гравитации элюировать один сфероид в центр каждой лунки, т. е. не выталкивайте какую-либо среду из кончика пипетки и позвольте капиллярному действию вывести сфероид из кончика пипетки. Для подтверждения элюирования содержимое пипетатора можно пипетировать обратно в 7 см чашку Петри под микроскопом.
- Используя микроскоп для рассечения и аппарат лайтбокса, перенесите сфероиды из чашки Петри на микропластину сфероидного анализа, как описано ниже.
3. Подготовка и загрузка соединений в картридж датчика для XF анализов
| Инъекционная стратегия | Соединение (порт) | Начальный объем микролунки XFe96 (мкл) | Желаемая конечная концентрация скважины | Объем порта (мкл) | Конечный объем микролунки XFe96 после впрыска (мкл) | Концентрация рабочего капитала |
| 1 | Олигомицин (А) | 180 | 3 мкг/мл | 20 | 200 | 30 мкг/мл |
| Ротенон (B) | 200 | 2 мкМ | 20 | 220 | 22 мкМ | |
| Антимицин А (В) | 200 | 2 мкМ | 20 | 220 | 22 мкМ | |
| 2 | БАМ15 (А) | 180 | 5 мкМ | 20 | 200 | 50 мкМ |
| Ротенон (B) | 200 | 2 мкМ | 20 | 220 | 22 мкМ | |
| Антимицин А (В) | 200 | 2 мкМ | 20 | 220 | 22 мкМ |
Таблица 2: Концентрации митохондриальных соединений для зондирования митохондриального энергетического метаболизма одиночных 3D-сфероидов с помощью анализатора XFe96.
- Подготовьте концентрации рабочего материала каждого соединения, как указано в таблице 2 , используя полностью дополненную, предварительно высушенную среду для анализа XF RPMI.
- Ориентируйте картриджную пластину (соединенную с полезной пластиной) по колоннам, 1-12 слева направо.
- Если используется направляющая заряжания, поместите ее на пластину картриджа в соответствии с процедурой хорошо нагружа, например, если порт А загружается первым, убедитесь, что А виден в левом верхнем углу направляющей.
- Перенесите рабочий раствор каждого соединения в подходящий резервуар и, используя калиброванную многоканальную пипетку P100, дозируйте 20 мкл во все соответствующие порты. Повторите для каждого соединения в остальные порты.
ПРИМЕЧАНИЕ: Если на пластине картриджа датчика не используются какие-либо порты, их можно оставить пустыми или заполнить пробирной средой. Если используется только выбор определенной буквы порта, убедитесь, что другие порты, соответствующие этой букве, загружены средой анализа; в противном случае воздух будет закачиваться в скважину, что ставит под угрозу результаты в этих скважинах. - После загрузки порта снимите направляющие для загрузки пластин (если они используются) и подготовьте анализатор к загрузке картриджа датчика.
ПРИМЕЧАНИЕ: Если анализ не выполняется сразу после загрузки портов, поместите крышку обратно на картридж датчика и поместите пластину обратно в воздушный инкубатор при температуре 37 °C до тех пор, пока она не будет готова к загрузке в машину.
4. Проектирование анализов, стратегии инъекций и сбор данных
- Проведение анализа
- Включите анализатор и подключите к контроллеру (компьютеру).
ПРИМЕЧАНИЕ: Это можно проверить по состоянию подключения прибора на панели виджетов программного обеспечения Wave Controller. - Перейдите на страницу шаблонов в программном обеспечении WAVE, найдите файл шаблона анализа для эксперимента и дважды щелкните его, чтобы открыть.
ПРИМЕЧАНИЕ: Если шаблон анализа не отображается в представлении Шаблоны , импортируйте файл шаблона в папку шаблона с общего сетевого диска или USB-устройства флэш-памяти. - Чтобы начать анализ, перейдите на вкладку Выполнить анализ .
ПРИМЕЧАНИЕ: Если определения групп были правильно распределены на карте пластин, анализ будет готов к выполнению, как указано зеленой галочкой в правой части страницы. На этом этапе любая дополнительная информация может быть введена на странице сводки анализа или на странице, оставленной пустой; перейдите к следующему шагу. В связи с задержкой проникновения митохондриальных модуляторов в 3D-сфероиды микротизов (рисунок 2) используют информацию протокола измерения, описанную в таблице 3.
- Включите анализатор и подключите к контроллеру (компьютеру).
| Период измерения | Номер впрыска и порт | Детали измерений | Продолжительность периода (ч:мин:с) |
| Калибровка | Не применимо | Анализаторы XF всегда выполняют эту калибровку, чтобы убедиться, что измерения точны | 00:20:00 (это среднее значение и может варьироваться в зависимости от машины) |
| Уравновешивание | Не применимо | Уравновешивание происходит после калибровки и рекомендуется. | 00:10:00 |
| Базальный | Не применимо | Циклы = 5 | 00:30:00 |
| Микс = 3:00 | |||
| Ожидание = 0:00 | |||
| Мера = 3:00 | |||
| Олигомицин / БАМ15 | Инъекция 1 (порт A) | Циклов = 10 | 01:00:00 |
| Микс = 3:00 | |||
| Ожидание = 0:00 | |||
| Мера = 3:00 | |||
| Ротенон + антимицин А | Инъекция 2 (порт B) | Циклов = 10 | 01:00:00 |
| Микс = 3:00 | |||
| Ожидание = 0:00 | |||
| Мера = 3:00 | |||
| Общее время: | 03:00:00 | ||
Таблица 3: Настройка протокола для зондирования митохондриального энергетического метаболизма одиночных 3D-сфероидов с помощью анализатора XFe96.
- Нажмите кнопку Начать запуск , чтобы открыть диалоговое окно расположение сохранения .
- Введите место сохранения результирующего файла и поместите собранный картридж датчика в термоотлив, который появляется из дверцы на боковой стороне анализатора. Подождите, пока тепловой лоток откроется автоматически и на экране отобразится сообщение Load Calibrant Utility Plate . Прежде чем следовать подсказкам на экране, убедитесь: i) правильное размещение картриджа датчика на пластине Утилиты, ii) снята крышка с картриджа датчика и iii) правильная ориентация картриджа датчика на вспомогательной пластине.
- Следуйте экранным командам, чтобы начать калибровку картриджа датчика.
ПРИМЕЧАНИЕ: Время, необходимое для завершения калибровки, составляет приблизительно 10-20 мин (для анализов при 37 °C). - После калибровки картриджа датчика загрузите сфероидную микропластинку в анализатор, следуя инструкциям на экране на Wave Controller, чтобы инициировать 12-минутный шаг равновесия.
ПРИМЕЧАНИЕ: Зеленые поля с белыми галочками указывают на «хорошую» калибровку для этого колодца. Если какие-либо скважины не обеспечивают «хорошую» калибровку, они будут обозначены красным прямоугольником и белым крестом. Такие скважины должны быть отмечены и исключены из любого анализа после завершения анализа с использованием вкладки модификационного анализа . - Подождите, пока анализатор автоматически начнет получать базовые измерения после того, как машина завершит этап равновесия (как указано в протоколе прибора).
- Чтобы завершить эксперимент, следуйте экранным командам на контроллере WAVE.
ПРИМЕЧАНИЕ: После того, как сфероидная микропластина была удалена из анализатора, отбросьте картридж датчика и отложите сфероидную пластину для дальнейшего анализа, если это необходимо (например, двухцепочечная (ds) количественная оценка ДНК). Если микропластинка не требуется для дальнейшего анализа, ее можно выбросить вместе с картриджем датчика. - Дождитесь появления диалогового окна анализа и просмотрите результаты или вернитесь в представление шаблонов .
5. Стратегии нормализации и анализа данных - нормализация после анализа и последующие анализы (необязательные шаги)
- Нормализация данных
- Чтобы нормализовать данные сфероидов, обратитесь к серии протоколов, относящихся к стратегиям нормализации данных для расчета размера и объема сфероидов и количественной оценки dsDNA в сфероидных анализах. Они были включены в качестве дополнительных файлов; см. Дополнительный файл 1 и Дополнительный файл 2.
- Анализ данных
- Чтобы экспортировать данные в один из генераторов автоматизированного анализа, выполните команды экспорта данных на контроллере WAVE и выберите генератор экспорта, соответствующий типу анализа. Кроме того, можно экспортировать файл данных и загрузить его в аналитику Seahorse.
ПРИМЕЧАНИЕ: Недостатком генераторов отчетов и аналитики Seahorse является то, что анализ данных ограничен тем, как разработан анализ XF, и не позволяет измерять средние значения по циклам измерения. Ручной экспорт наборов данных из программного обеспечения прибора позволяет использовать предпочтения пользователя в этом отношении. Учитывая, что стратегия инъекций для оценки митохондриального дыхания 3D-сфероидов, вероятно, будет отличаться от типичного теста «MitoStress», была разработана серия шаблонов электронных таблиц, чтобы помочь проанализировать эти наборы данных, специфичные для 3D-клеточных культур, и будет предоставлена по запросу. Эти файлы шаблонов данных предоставят данные о ключевых параметрах митохондриальных дыхательных путей, подробно описанных и объясненных на рисунке 2. - Чтобы проанализировать данные, экспортируйте данные в виде отчета электронной таблицы из программного обеспечения контроллера WAVE и используйте независимый шаблон электронной таблицы для анализа.
- Чтобы экспортировать данные в один из генераторов автоматизированного анализа, выполните команды экспорта данных на контроллере WAVE и выберите генератор экспорта, соответствующий типу анализа. Кроме того, можно экспортировать файл данных и загрузить его в аналитику Seahorse.

Рисунок 2: Схематические дескрипторы для параметров, полученных в результате анализа данных о внеклеточном потоке. Аббревиатура: OCR = норма потребления кислорода. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Для получения хорошо сформированных, компактных сфероидов каждую клеточную линию оптимизировали индивидуально по плотности посева и продолжительности культивирования (рисунок 3). Клеточные линии A549, HepG2/C3A и SK-OV-3 первоначально образовывали рыхлые агрегаты, которые не прогрессировали до круглых сфероидов с четко определенными периметрами до 7 дней в культуре. И наоборот, клетки MCF-7 могут образовывать сфероиды в течение 3 дней. Была выявлена четкая корреляция между начальной плотностью посева клеток и объемом сфероидов после периода культивирования для всех сфероидных моделей. Размер сфероида и морфология были оптимизированы для плотности посева. Морфология и циркулярность начали снижаться с увеличением размера сфероида во всех моделях. Стратегии посева для клеточных линий были оптимизированы при 4 × 103 ячейках/лунке для клеток A549 и SK-OV-3; Ячейки HepG2/C3A были ранее оптимизированы в других местах до 1 × 103 ячеек/лунка, а клетки MCF-7 использовались при 4 × 103 ячейках/лунке во всех анализах. При оптимизированных стратегиях посева объем сфероидов составлял от 5,46 × 107 мкм3 (SK-OV-3) до 1,45 × 108 мкм3 (A549) (рисунок 3B). Все типы сфероидов имели линейную корреляцию между начальной плотностью посева и объемом сфероида, где A549 и HepG2/C3A имели значенияR2 0,957 и 0,947 соответственно. Было обнаружено, что сфероидные объемы MCF-7 и SK-OV-3 имеют большую корреляцию с начальной плотностью посева,R2=0,977 (рисунок 3A).
Сфероидная циркулярность была рассчитана с использованием планиметрии изображений в аналитическом программном обеспечении FIJI с использованием длинных и коротких диаметров сфероидов. Идеальная сфероидная симметрия имела цикличность = 1,0; отклонение от 1,0 указывает на потерю циркулярности (рисунок 3C). Циркулярность была выше у сфероидов MCF-7, чем у других моделей, где циркулярность поддерживалась между 0,83 и 0,9 при всех плотностях посева. Для сравнения, внешний периметр сфероидов SK-OV-3 не был так четко определен, а объем сфероидов был значительно меньше даже после 7 дней в культуре, что давало сфероиды с максимальной циркулярностью 0,61 при плотности посева 4 × 103/лунка. Было также обнаружено, что клетки HepG2/C3A образуют плотные, хорошо сформированные сфероиды с равномерной морфологией по всей площади поверхности всех сфероидов, с циркулярностью, поддерживаемой на уровне 0,79 для клеток, засеянных при 1 × 103 клетках / хорошо. Клетки A549, по-видимому, следовали тенденции, в которой сфероидная цикличность и морфология были усилены плотностью посева; однако циркулярность не превышала 0,63 при плотности, использованной в этих экспериментах.
Базальное митохондриальное дыхание рассчитывали как OCR, измеренное из сфероидов, посеянных в 1 × 103, 2 × 103, 4 × 103 или 8 × 103 клеток / хорошо в микропластинах ультранизкого прикрепления сфероидной культуры (рисунок 3D). Для всех типов сфероидов OCR увеличивался с размером сфероида и линейно коррелировал с объемом сфероида сR2 самым высоким в сфероидах MCF-7 при 0,988 и самым низким в сфероидах SK-OV-3 при 0,744 (рисунок 3E). Измеренный OCR статистически отличался между всеми экспериментальными группами. A549 имел самый низкий OCR, достигая только 18 пмоль / мин / скважина при наибольшем размере сфероида (рисунок 3D). И наоборот, сфероиды MCF-7 дали аналогичный OCR при наименьшем размере сфероида всего через 3 дня в культуре, достигнув максимального базового OCR 53 пмоль / мин / лунка для наибольшего размера сфероида (рисунок 3D). HepG2/C3A дал данные OCR, в значительной степени согласующиеся с размером сфероида и морфологией. В сфероидах HepG2/C3A, посеянных из 1 × 103 клеток/лунка, исходное OCR достигало в среднем 15 пмоль/мин/лунка, увеличиваясь до максимума 52 пмоль/мин/лунка в крупнейших сфероидах (рисунок 3D). OCR в сфероидах SK-OV-3 был значимым только между сфероидами, выращенными из 4 × 103 клеток / лунки и 8 × 103 клеток / хорошо, с небольшой разницей, наблюдаемой в сфероидах, выращенных между 1 × 103, 2 × 103 или 4 × 103 клетками. Несмотря на различия в размерах, данные OCR были очень похожи между сфероидами HepG2 / C3A и MCF-7 во всех точках размера. Относительно размера сфероида (мкм3) исходное OCR сфероидами MCF-7 было сопоставимо с фоноидами HepG2/C3A, выращенными в течение 7 дней из 1000 клеток на лунку.

Рисунок 3: Параметры роста сфероидов определяют исходное митохондриальное дыхание. (A) A549, HepG2/C3A, MCF-7 и SK-OV-3 сфероиды были оптимизированы для плотности посева и их роста, контролируемого при 1 × 103, 2 × 103, 4 × 103 и 8 × 103 клеток /колодец в каждой из фотомикрографов, от верхнего левого до нижнего правого края соответственно; шкала стержней = 500 мкм. (B) Размер сфероида рассчитывался с использованием планиметрических данных с собранных микрофотографий и сравнивался с использованием статистики корреляции Пирсона; пунктирные линии представляют собой распределение 95% доверительного интервала. (C) Морфология сфероидов сравнивалась путем расчета циркулярности. (D) OCR измеряли в 5 раз, после чего добавляли ротенон-антимицин А для учета немитохондриальной частоты дыхания с помощью анализатора Agilent Seahorse XFe96. Базальный OCR, измеренный как OCR - OCRr/a сравнивали между плотностью посева (D) и объемом сфероида (E). Данные представляют собой средние значения, ± SEM из 5-8 реплицированных скважин на тип сфероида и плотность посева клеток. Сокращения: OCR = норма потребления кислорода; OCRбазальное = базальное митохондриальное дыхание; OCRr/a = OCR после добавления ротенона-антимицина A. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Концентрация и время воздействия респираторных модуляторов в XF-анализе является критическим шагом в оптимизации анализа. Респираторные модуляторные соединения - олигомицин, BAM15, смесь ротенона-антимицина A- или управление транспортным средством DMSO последовательно вводили через инжекционные порты картриджа датчика в микропластинчатые колодцы, содержащие сфероиды MCF-7 (фиг.4A). Были завершены четыре цикла измерения для определения среднего базального OCR 30-40 пмоль/мин/лунка для всех групп образцов. В течение оставшейся продолжительности анализа дыхательные модуляторы последовательно добавляли каждые 5 циклов измерения для достижения конечных концентраций в скважине 0,5 мкМ (0,5 мкг/мл олигомицина) после инъекции 1; 2,0 мкМ (2 мкг/мл олигомицина) после инъекции 2; 5 мкМ (5 мкг/мл олигомицина) после инъекции 3; и, наконец, максимальная концентрация в скважине 11 мкМ (11 мкг/мл олигомицина) после четвертого последовательного добавления. Сфероиды MCF-7 не реагировали на управление транспортным средством на протяжении всего эксперимента (рисунок 4А). Базальный OCR немедленно изменялся после первой инъекции каждого соответствующего соединения при самой низкой концентрации 0,5 мкМ или 0,5 мкг/мл олигомицина (рисунок 4B). OCR в сфероидах MCF-7 понижали с олигомицином с 41 пмоль/мин/лунка до 23 пмоль/мин/хорошо после 5 циклов измерения после первой инъекции 0,5 мкг/мл (рисунок 4B).
В ответ на 0,5 мкм BAM15 OCR был увеличен с 33 до 41 пмоль/мин/задолго до второй инъекции (рисунок 4C). Для сравнения, комбинация ротенона и антимицина А снижала OCR с 37 до 13 пмоль/мин/задолго до второй инъекции (рисунок 4D). Кинетические следы дополнительно выявили устойчивое линейное снижение (олигомицин и ротенон-антимицин А) или увеличение (BAM15) в OCR. Для всех режимов дозирования соединений стационарная OCR была достигнута в течение 10-12 полных циклов измерения (60-72 мин) при общей концентрации в скважине 2 мкМ BAM15, 2 мкМ ротенона, 2 мкМ антимицина А и 2 мкг/мл олигомицина (рисунок 4А). Скорость потребления кислорода достигла стабильного плато при ~19 пмоль/мин/лунка (олигомицин), 52 пмоль/мин/лунка (БАМ15) и 10 пмоль/мин/лунка (ротенон-антимицин А) (рисунок 4А). Увеличение концентрации соединения олигомицина, BAM15 или ротенона плюс антимицин А также не оказывало очевидного влияния на OCR, которое оставалось постоянным на протяжении всей оставшейся части анализа. Эти данные демонстрируют, что как концентрация соединения, так и временная продолжительность воздействия респираторных модуляторных соединений должны учитываться для оптимизации анализа при использовании 3D-сфероидов.

Рисунок 4: Титрование респираторных модуляторных соединений как важный шаг для оптимизации анализа внеклеточного потока. (A) Сфероиды MCF-7 высыпали на 4 × 103 клетки/лунку и культивировали в течение 3 дней, прежде чем поместить в скважины сфероидной микропластины, содержащей XF RPMI, и исследовали на предмет OCR ± митохондриальных модуляторов с использованием анализатора XFe96. OCR измеряли в 5 раз, после чего добавляли титрование либо контроля транспортного средства, олигомицина (B), BAM15 (C) или ротенона-антимицина A (D) для ингибирования митохондриальной АТФ-синтазы, определения максимальной дыхательной способности или установления немитохондриальной частоты дыхания, соответственно. Концентрацию каждого митохондриального модулятора увеличивали в течение 4 отдельных стратегий инъекций титрования (0,5 мкМ, 1,5 мкМ, 3 мкМ и 6 мкМ; единицы для олигомицина составляют мкг/мл) для определения максимального стационарного OCR в ответ на оптимальную концентрацию соединения. OCR измеряли в течение 5 циклов измерения между каждой инъекцией. Данные представляют собой средние значения ± SEM из 5-8 отдельных реплик скважин. Аббревиатура: OCR = норма потребления кислорода. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Одним из основных преимуществ технологии XF является способность исследовать функцию митохондрий в интактных клетках и тканях. Для изучения конкретных аспектов митохондриальной функции в клетках и тканях митохондриальные модуляторы последовательно добавляются к отверстиям микропластины образца через 4 доступных порта для инъекций на картридже датчика. Типичной последовательностью модуляторов, используемых для исследования митохондриальных параметров в анализах XF, являются олигомицин, протонофор (например, FCCP или BAM15) и комбинация ротенона плюс антимицин А, которые добавляются последовательно для ингибирования митохондриальной АТФ-синтазы, определения максимальной дыхательной способности и корректировки на немитохондриальную частоту дыхания, соответственно. Эта типичная последовательность добавлений модулятора называется тестом MitoStress производителем технологии анализа. Учитывая, что олигомицин может ингибировать дыхание, стимулируемое непарой, в некоторых клеточных монослоях20, мы исследовали это с помощью 3D-сфероидов, полученных из рака, путем измерения несвязанного стимуляционного OCR (OCRmax) до (одиночной) и после (последовательной) инъекции олигомицина (рисунок 5A-D). OCRmax не был достоверно ограничен добавлением олигомицина в сфероиды, образованные из HEPG2/C3A или SK-OV-3 (Рисунок 5E и Рисунок 5G). Тем не менее, OCRmax был значительно снижен в сфероидах A549 и MCF-7 после последовательной инъекции BAM15 после олигомицина по сравнению сOCR max, достигнутым от одной инъекции BAM15 (рисунок 5F и рисунок 5H). Если не известно иное, поэтому рекомендуется использовать отдельные лунки для лечения олигомицином и разъединителем, с окончательным добавлением ротенона и антимицина А при исследовании митохондриального энергетического метаболизма 3D-сфероидов. Этот подход по-прежнему позволяет вычислять все митохондриальные параметры, как в типичном тесте MitoStress, где соединения добавляются последовательно.

Рисунок 5: Однократная или последовательная инъекция митохондриальных респираторных соединений. Сфероиды MCF-7, HEPG2/C3A, SK-OV-3 и A549, полученные из раковых клеток, были помещены в скважины сфероидной микропластины XFe96 в XF RPMI и исследованы на ПРЕДМЕТ OCR с использованием анализатора Agilent Seahorse XFe96. OCR измеряли в 5 раз, после чего 2 мкг/мл олигомицина (инъекционный порт A: зеленый след) или 5 мкМ BAM15 (инъекционный порт A: синий след или инъекционный порт B: зеленый след) для ингибирования митохондриальной АТФ-синтазы и определения максимальной дыхательной способности соответственно. Кинетические данные OCR выражаются в % базальных (A-D). Максимальная дыхательная емкость (OCRmax) рассчитывалась как коэффициент базального OCR по уравнению: OCRmax = OCRBAM15 / OCRбазальный. OCRmax был получен из средних значений OCR по циклам измерения 8-10 после инъекции BAM15 с (зелеными полосами) и без (синие полосы) олигомицина. Данные представляют собой средние значения ± SEM из 3-8 отдельных реплик скважин по микропластине сфероидного анализа. Сокращения: OCR = норма потребления кислорода. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Используя оптимальную плотность посева клеток, концентрации соединений, стратегию инъекции и период цикла измерения, определенные в этих экспериментах по оптимизации (таблица 3), мы разработали подробный протокол для точного зондирования базального митохондриального дыхания:базальное OCR (рисунок 6A), дыхание фосфорилирования ADP: OCRADP (рисунок 6B), дыханиеутечки : OCR omy (рисунок 6C), эффективность связи (рисунок 6D) ), максимальная дыхательная способность:OCR max (рисунок 6E) и резервная дыхательная способность:OCR резервная (рисунок 6F) с использованием 3D-сфероидов, полученных из рака.

Рисунок 6: Зондирование OCR с помощью технологии XF для установления митохондриального энергетического метаболизма сфероидов, полученных из рака. Сфероиды MCF-7, HEPG2/C3A, SK-OV-3 и A549, полученные из раковых клеток, были помещены в скважины сфероидной микропластины в XF RPMI и исследованы на предмет OCR с использованием анализатора Agilent Seahorse XFe96. OCR измеряли в 5 раз, после чего 2 мкг/мл олигомицина, или 5 мкМ BAM15, и добавляли РА для ингибирования митохондриальной АТФ-синтазы, определения максимальной дыхательной способности и расчета немитохондриальной частоты дыхания, соответственно. (A) Базальное митохондриальное дыхание (базальное OCR) было рассчитано как среднее значение OCR из 3 циклов измерения до инъекции порта A. (B) Эффективность связи окислительного фосфорилирования аппроксимировалась путем экспрессии OCRADP (OCRбазально-OCR leak) в процентах отБАЗАЛЬНОГО OCR. (C) АдФ-фосфорилирование (OCRADP) измеряли как Олигомицин-чувствительное OCR, рассчитанное на основе усредненного OCR в циклах измерения 11-13 до инъекции BAM15. (D) Дыханиеутечки (утечка OCR) измерялось как OCR, нечувствительное к олигомицину, рассчитанное на основе среднего усредненного OCR в циклах измерения 11-13. (E) Максимальная дыхательная способность (OCRmax) измерялась как средний максимальный OCR, измеренный после инъекции BAM15. (F) Запасная дыхательная способность была рассчитана путем экспрессиизапасного OCR (OCRmax - OCRbasal) в процентах от OCRбазального. OCR после инъекции ротенона-антимицина А (OCRr/a) вычитали из всех показателей с поправкой на немитохондриальную OCR. Данные представляют собой средние значения ± SEM из 3-8 отдельных реплик скважин по сфероидной пластине XFe96. Сокращения: OCR = норма потребления кислорода; РА = 2 мкМ ротенона-2 мкМ антимицина А. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Сфероиды MCF-7, выращенные из 4 × 103 клеток / в течение 3 дней, использовались в качестве модели для определения оптимального переноса, размещения и анализа в микропластинах сфероидного анализа. Используя размеры, предусмотренные для сфероидной микропластины от производителя, поверхность скважины была разделена на три зоны-зоны для оптимального размещения сфероидов (рисунок 7А), где зона 1 была выделена как оптимальная зона в центре скважины. При тщательном пипетке с использованием широкодисперсных наконечников пипеток сфероиды переносились в сфероидные пластины и случайным образом распределялись по поверхностям скважин гравитационным элюированием (рисунок 7B). Там, где сфероиды были тщательно перенесены с использованием гравитационного элюирования, большинство сфероидов обычно можно было найти в зонах 1-2 микропластины, используя рекомендуемые методы переноса от производителя. Там, где сфероиды вытеснялись из кончика пипетки путем аспирации, сфероиды часто помещались за пределы этих зон и не могли быть замечены с помощью микроскопии.
Для сравнения положений расположения сфероидов сфероиды MCF-7 переносили в сфероидные пробирные микропластины в обозначенных зонах 1-3 или вне зоны (рис. 7А). Эти 4 скважины отслеживались с помощью кинетического эксперимента OCR на исходном уровне и после добавления олигомицина, BAM15 или ротенона-антимицина A (рисунок 7C). OCR был рассчитан по среднему значению трех показаний цикла перед каждой инъекцией (рисунок 7B). OCR измеряли кинетически в течение 200 мин в 4 выбранных скважинах (рисунок 7C) и с коррекцией исходного уровня (рисунок 7D). Там, где сфероиды были помещены в зону 3 или вне зоны, базовые OCR были значительно ниже, чем сфероиды, помещенные в зоны 1 и 2 (рисунок 7C). Эффекты респираторных соединений олигомицина, BAM15 и ротенона-антимицина А также резко различались между сфероидами, помещенными в зоны 1 и 2, по сравнению с зоной 3 и внезонными областями. Увеличение OCR наблюдалось при применении олигомицина в сфероидах, помещенных в зону 3 или вне зоны (рисунок 7E). Кроме того, сфероиды, помещенные в зону 3 или вне зоны, испытывали чрезмерно высокий ответ на BAM15 с OCR выше исходного уровня после инъекции ротенона-антимицина А (рисунок 7E). Несмотря на почти двукратное увеличение базального OCR (рисунок 7C) со сфероидами, помещенными в зону 2 по сравнению с зоной 1, складчатые изменения в ответ на все респираторные соединения были очень похожи (рисунок 7E), предполагая, что различия в базальном OCR между сфероидами, помещенными в зоны 1 или 2, вряд ли будут результатом размещения в скважине.

Рисунок 7: Размещение сфероидов внутри микропластины сфероидного анализа диктует базальные эффекты OCR и митохондриального модулятора с использованием технологии XF. Сфероиды MCF-7 были посеяны на 4 × 103 клетки / лунка и культивированы в течение 3 дней, прежде чем их поместили в скважины сфероидной микропластины, содержащей XF RPMI, и исследовали на предмет OCR ± митохондриальных модуляторов с использованием анализатора Agilent Seahorse XFe96. (A) Микрофотографии положений сфероидных зон в микропластинках сфероидного анализа после продолжительности анализа; шкала bar = 500 мкм и OCR, захваченная из соответствующих скважин с течением времени, выраженная либо pmol/min-1/well-1 (B), либо % базальной (C). (D) эффекты митохондриального модулятора сфероидов MCF-7, размещенных в различных зонах в микропластине сфероидного анализа; данные, выраженные в виде изменения складки от базального. (E) Пример кинетической трассировки, выделяющей, какие измерения данных OCR (красные круги) используются для расчета отклика каждого митохондриального модулятора на данные, представленные в E. Показанные данные взяты из индивидуальных реакций скважин. Аббревиатура: OCR = норма потребления кислорода. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Критерии отбора фона имеют большое значение; использование самых внешних скважин для фоновой коррекции не является репрезентативным для всех микропластинчатых скважин, что может привести к неправильным предположениям данных и ошибочным выводам данных из-за краевых эффектов по всей сфероидной микропластине. Чтобы оценить это наблюдение, сфероиды MCF-7 использовались для сравнения процедур коррекции анализа для получения значений OCR в ответ на добавление контроля транспортного средства, олигомицина, BAM15 или ротенона-антимицина A (рисунок 8). Все респираторные соединения давали ожидаемые кинетические профили OCR для выбранных соединений, выявляя среднюю устойчивую скорость базального дыхания 20-30 пмоль/мин/лунку (рисунок 8А). Однако там, где данные анализа анализировались с использованием самых внешних скважин для коррекции фоновой температуры (A1, A12, H1 и H12), значения, выявленные для OCR после добавления респираторных соединений, были особенно низкими; OCR дал отрицательные значения для ротенона-антимицина А. В ответ на эти наблюдения альтернативный анализ проводили с использованием серии пустых скважин, случайным образом распределенных по сфероидной микропластине, в качестве скважин фоновой температурной коррекции (рисунок 8B). В тех случаях, когда применялась альтернативная коррекция фона, все относительные составные эффекты на OCR были одинаковыми в обоих аналитических наборах; однако абсолютные значения OCR увеличились примерно на 10 пмоль/мин/лунка (рисунок 8). Эти данные подчеркивают силу и важность коррекции фоновой температуры на микропластинках сфероидного анализа и подчеркивают важность оптимизации пользователя для анализа XF.

Рисунок 8: Случайный выбор скважин для коррекции фона для улучшения контроля градиентов температуры на микропластине сфероидного анализа. Данные OCR, экстраполированные из рисунка 2A с использованием рекомендуемых скважин для фоновой коррекции (A) по сравнению со случайно назначенными скважинами для фоновой коррекции (B). Аббревиатура: OCR = норма потребления кислорода. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
В отличие от клеточных монослоев, сфероиды представляют собой гетерогенную агрегацию клеток в 3D-пространстве и поэтому требуют тщательного рассмотрения в отношении анализа, особенно при нормализации этих данных. В данной работе представлены три подхода к нормализации XF-данных, полученных из сфероидов MCF-7 (рисунок 9). При ненормализованном OCR положительно коррелирует (R2 = 0,98) с размером сфероида (определяемым начальной плотностью посева клеток) значительно при сравнении статистически с коэффициентом корреляции Пирсона, P = 0,0057 (рисунок 9A). Эта линейная зависимость понижается, когда OCR нормализуется до начальной плотности посева клеток (R2 = 0,78) и больше не коррелирует с размером сфероида (P = 0,117, рисунок 9B). Это также имеет место при нормализованном до сфероидного объема (R2 = 0,77; Коэффициент корреляции Пирсона P = 0,120, рисунок 9C) и содержание ядерной дцДНК (R2 = 0,58; Коэффициент корреляции Пирсона P= 0,233, рисунок 9D). Эти данные подчеркивают важность нормализации данных XF при зондировании митохондриального метаболизма сфероидов, особенно если они имеют разные размеры.

Рисунок 9: Нормализация данных о внеклеточном потоке, полученных от клеточных сфероидов. (A) Необработанные данные OCR были получены из MCF-7, культивируемого в течение 3 дней, и построены с использованием модели Пирсона для получения коэффициента корреляции между плотностью посева сфероидов и OCR; Значение P установлено равным 0,05. (B) Необработанные данные OCR были нормализованы по отношению к начальной плотности посева сфероидов; (C) объем сфероида MCF-7, полученный с помощью микроскопической планиметрии; и (D) содержание ядерной DS ДНК, сравниваемое с использованием коэффициента корреляции Пирсона. Сокращения: OCR = норма потребления кислорода; ds ДНК = двухцепочечная ДНК. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
| Клеточная линия | Плотность посева (скважина) | Рост сфероидов (дней) | Конечный объем сфероида (мкМ3) | Базальный OCR (pmolO2/мин/лунка) | Чувствительность для базального OCR достигнута (ДА/НЕТ) |
| СКОВ | 1000 | 5 | 9.52Е+06 | 28 ± 3,5 | Да |
| СКОВ | 2000 | 5 | 2.38Е+07 | 26 ± 1.4 | Да |
| СКОВ | 4000 | 5 | 4.92Е+07 | 36 ± 3,1 | Да |
| СКОВ | 8000 | 5 | 1.11Е+08 | 50 ± 7.9 | Да |
| ГепГ2 | 1000 | 5 | 1.11Е+07 | 15 ± 0.7 | Нет |
| ГепГ2 | 2000 | 5 | 2.88E+07 | 23 ± 1,8 | Да |
| ГепГ2 | 4000 | 5 | 5.46Е+07 | 31 ± 1.7 | Да |
| ГепГ2 | 8000 | 5 | 1.21E+08 | 52 ± 2,8 | Да |
| А549 | 1000 | 5 | 2.11Е+07 | 30 ± 2,5 | Да |
| А549 | 2000 | 5 | 3.57Е+07 | 41 ± 1.6 | Да |
| А549 | 4000 | 5 | 6.93E+07 | 53 ± 7,2 | Да |
| А549 | 8000 | 5 | 1.44E+08 | 65 ± 8,4 | Да |
| МКФ-7 | 1000 | 3 | 1.60E+07 | 29 ± 0.8 | Да |
| МКФ-7 | 2000 | 3 | 2.52Е+07 | 37 ± 1.7 | Да |
| МКФ-7 | 4000 | 3 | 6.00Е+07 | 46 ± 1.7 | Да |
| МКФ-7 | 8000 | 3 | 1.06Е+08 | 66 ± 2.9 | Да |
Таблица 4: Оптимизированы параметры для определения базальных измерений OCR в одиночных 3D сфероидах. Аббревиатура: OCR = норма потребления кислорода.
Дополнительный файл 1: Анализ размера и объема сфероида. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный файл 2: Количественная оценка двухцепочечной ДНК из сфероидов в сфероидной микропластине. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный файл 3: Рекомендации по количеству реплик, необходимых для получения надежных наборов данных XF-анализа. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Основные выводы и результаты
В этой статье представлен подробный протокол для исследования митохондриального энергетического метаболизма отдельных 3D-сфероидов с использованием серии клеточных линий, полученных из рака, с помощью анализатора XFe96 XF. Разработан и описан способ быстрого культивирования клеточных сфероидов A549, HepG2/C3A, MCF7 и SK-OV-3 с использованием клеточно-репеллентных технологий принудительной агрегации. Этот протокол учитывает многие соображения зондирования метаболизма сфероидов с помощью технологии XF, включая (1) оптимизацию протоколов сфероидных культур и обработку и передачу сфероидов в конкретные микропластины сфероидного анализа от производителя технологии из их оригинальных сосудов культивирования; 2) концентрация используемых респираторных соединений и временная зависимость проникновения соединения; 3) стратегии инъекций, которые будут использоваться; и 4) способы нормализации данных между экспериментальными группами. Все эти соображения были рассмотрены в настоящем документе и более подробно обсуждаются ниже. Эти методы представлены в виде упрощенных подходов к генерации данных о последовательном метаболическом потоке кислорода с использованием одиночных 3D-сфероидов с анализатором XFe96 Flux. Этот экспериментальный подход может быть использован в качестве отправной точки и рубрики для использования в других сфероидных моделях, которые легко реализуются в базовых лабораторных условиях.
Соображения
Рост сфероидов, размер и чувствительность технологии XF
Чтобы установить воспроизводимые данные с помощью технологии XF, важно охарактеризовать и оптимизировать анализ для конкретной модели. Этот подход относительно прост в базовом монослое клеток; однако это создает дополнительные проблемы при культивировании клеток в качестве 3D-сфероидов. В ходе экспериментов, представленных здесь, среда RPMI от производителя была дополнена при использовании. Хотя отмечается, что некоторые клеточные линии, а именно HepG2/C3A, культивировались в среде роста DMEM, во время этих относительно коротких анализов (~3-5 ч) замена препаратами RPMI-DMEM оказала ограниченное влияние на анализ XF. Формулировки двух сред очень похожи, и пользователи могут «настроить» среду Seahorse RPMI в соответствии с матрицей своих сред клеточной культуры путем добавления, например, увеличения глюкозы, дальнейшего добавления источников углеводов. Критически важным для окончательной формулировки всех буферов и сред XF является отсутствие фенол-красного цвета, который, вероятно, будет мешать флуоресцентным зондам внутри картриджной пластины зонда XF, и бикарбоната натрия, что приведет к щелочности из-за отсутствия буферизации CO2 , присутствующей в инкубаторах клеточных культур. Другие носители и буферы могут быть приобретены и/или изготовлены собственными силами. Например, буфер Krebs Ringer HEPES представляет собой простой буфер, который можно использовать для оценки дыхания во многих различных клетках, включая сфероидные модели. Тем не менее, пользователи XF-анализов должны отметить, что изменение среды/буфера и его добавление могут изменить его общую буферную способность. Это вызывает особую озабоченность, когда пользователи могут быть заинтересованы в измерении ECAR, в котором необходимо оценить буферный коэффициент среды, чтобы позволить преобразование ECAR в скорость выброса протонов (PER).
Поскольку клеточный OCR, измеренный технологией XF, пропорционален плотности клеток, когда число клеток в скважине находится в пределах чувствительности системы, было важно исследовать эту связь с использованием одиночных 3D-сфероидов. Исследуя OCR одиночных 3D-сфероидов, культивируемых из 4 различных линий раковых клеток, засеянных с плотностью 1000, 2000, 4000 или 8000 клеток на лунку, мы показываем, что анализатор XFe96 достаточно чувствителен, чтобы улавливать изменения скорости митохондриального дыхания между 3D-сфероидами, выращенными из разных плотностей посева клеток (рисунок 3). Показано, что оптимальный диапазон плотности посева клеток, а значит и сфероидного объема для формирования 3D сфероидов для зондирования OCR, различаются в зависимости от типа клетки. Об этом свидетельствует линейная зависимость между OCR и плотностью посева или сфероидным объемом (рисунок 3). Для клеток A549 и HepG2/C3A оптимальная плотность посева для чувствительности к OCR составляла от 1000 до 8000 клеток на лунку; это было 2000-8000 клеток / лунка для MCF-7 и 4000-8000 клеток / лунка для клеток SK-OV-3. Эти данные демонстрируют, что оптимизация размера сфероида имеет особое значение при оценке OCR с использованием технологии XF.
Соображения по минимальным и максимальным объемам сфероидов и базальному OCR
В общем, всегда будут минимальные и максимальные пороговые значения для измеримых параметров OCR, рекомендованные производителем для этих экспериментов. Для анализатора XFe96 базальный OCR между 20 пмоль O2/мин/лунка и 200 пмоль O2/мин/лунка являются нижним и верхним пределами соответственно. Это относится к однослойным клеткам и сфероидам, и то, где экспериментальная модель находится в пределах этого динамического диапазона OCR, будет зависеть от количества доступного биологического материала, например, количества клеток в качестве монослоя или размера сфероидов. См. таблицу 4 для примера того, как пороговые значения OCR были достигнуты сфероидными моделями, используемыми здесь. Возможно, было бы разумно проверить уровень кислорода в скважине, для которой эти данные также доступны из этих измерений в качестве данных об уровне. Это следует регулярно рассматривать в каждом эксперименте в целях контроля качества. Если в скважине произойдет истощение кислорода, это станет очевидным в данных. В этом случае может потребоваться корректировка циклов измерения в рамках эксперимента; например, увеличение стадии перемешивания таким образом, чтобы уровень кислорода в скважине восстанавливался до следующего периода измерения в рамках цикла измерения. Хотя это возможно, мы обнаружили, что это очень маловероятно для экспериментов с одним сфероидом с использованием описанных клеточных линий.
Выбор митохондриальных разъединителей для анализа внеклеточного потока
Протонные ионофоры, такие как карбонилцианид 4-(трифторметокси) фенилгидразон (FCCP)21, карбонилцианид м-хлорфенилгидразон (CCCP)22 или BAM1523, являются мощными низкомолекулярными химическими веществами, способными нарушать электрохимический протонный градиент через митохондриальные мембраны, ингибировать выработку АТФ и, в конечном счете, разъединять митохондриальное дыхание24 . Для этих целей продолжают разрабатываться новые малые молекулы, особенно при лечении метаболических заболеваний 25,26,27; обратитесь к двум отличным отзывам28,29. И наоборот, разъединение окислительного дыхания было связано с нежелательной нецелевой токсичностью30. Однако в клеточных анализах in vitro молекула FCCP деполяризует митохондриальный мембранный потенциал и оказывает нецелевые эффекты, такие как деполяризация плазматической мембраны, нарушая поток ионов NA+ 31; вмешательство в обработку клеточного белка32 и даже индуцирование клеточного старения33. BAM15 был первоначально введен в 2013 году как митохондриальный разъединитель с минимальным влиянием на плазматические мембраны23, с протонофорной активностью в микромолярном диапазоне в целых клетках и наномолярном диапазоне в изолированных митохондриях 23,34.
Учитывая эффективность FCCP по деполяризации плазматической мембраны, BAM15 является более надежным протонофором для разъединения дыхания в интактных целых клетках во внеклеточных флюс-анализах. Хотя FCCP и его аналог, CCCP, использовались более 50 лет для анализа максимальной дыхательной способности и продолжают широко использоваться в исследованиях XF, использование этих малых молекул часто недооценивает митохондриальную и клеточную метаболическую способность. Это частично связано с тем, почему так много публикаций, использующих технологию XF, попадают в ловушку, сообщая об отрицательных резервных дыхательных возможностях или недооценивая истинные митохондриальные дыхательные возможности при использовании FCCP. Дополнительная эффективность FCCP в интактных клетках и тканях часто приводит к нарушению митохондриальной функции, и клетки могут изо всех сил пытаться работать надлежащим образом, чтобы поддерживать максимальную дыхательную способность в течение нескольких циклов измерения после их добавления, даже при очень низких концентрациях35. Таким образом, реакция клеток на FCCP может быть найдена во многих исследованиях, чтобы снизиться после начального периода цикла измерения. В то время как FCCP обычно используется для анализа XF, BAM15 используется предпочтительно в случаях, связанных с целыми клетками или сфероидными моделями, учитывая, что он может поддерживать максимальную дыхательную способность в полностью деполяризованных митохондриях при концентрациях до 10 мкМ 3. Кроме того, BAM15 индуцирует воздействие на внеклеточное подкисление, которое совпадает с окислением питательных веществ через гидратацию CO2 с образованием HCO3- и H+ в большей степени, чем FCCP3. Тем не менее, в случае изолированных митохондрий и пермеабилизированных клеток, любой из этих разъединителей должен работать так же, как BAM15 для разъединения митохондрий, если титровать в правильной концентрации.
Кинетика проникновения соединений и цикличность анализа
Концентрации, проникновение и кинетические профили химических соединений, используемые для проведения типичного теста MitoStress с 3D-сфероидами с использованием анализатора XF, более сложны для решения. Учитывая, что сфероиды имеют 3D-структуру, проникновение молекул по всему диаметру сфероида является бесконечно более сложным процессом, чем через монослои клеток. Например, кинетическое проникновение и, следовательно, чувствительность к химиотерапевтическому сорафенибу определяли по сфероидному возрасту и, следовательно, размеру в сфероиде HepG2 модели36. Способность низкомолекулярных химических веществ (например, лекарств, наночастиц) достигать биологической мишени зависит от нескольких основополагающих факторов, включая сложность системы, которая динамически проникает и диффундирует через37,38. Это особенно верно для препаратов, нацеленных на опухолевуюткань 39. Подобно нацеливанию на опухоль в контексте 3D-сфероида, размер, компактность и другие фенотипические реакции, такие как экспрессия белков-транспортеров лекарств, могут управлять временем проникновения и концентрацией соединения, необходимого для получения биологического ответа.
В этом протоколе мы рассмотрели вопрос о времени проникновения и концентрации малых молекул в ответ на ингибитор АТФ-синтазы олигомицин, протонофор и митохондриальный разъединитель BAM15, а также комбинацию ингибиторов комплекса I и комплекса III ротенона и антимицина А. Исследуя OCR отдельных сфероидов MCF-7, подвергшихся многократному титрованию этих распространенных респираторных соединений, мы демонстрируем, что оптимальная концентрация каждого соединения, необходимая для индуцирования стационарной скорости дыхания, находится в пределах того же диапазона, что и монослойные клетки (рисунок 4). Важно отметить, что, отличаясь от своих однослойных аналогов, показано, что увеличение числа циклов измерения между инъекциями является ключом к достижению стационарного OCR в одиночных 3D-сфероидах. Эти данные подчеркивают важность проникновения соединений и их соответствующих кинетических профилей при изучении параметров митохондриальных дыхательных путей 3D-сфероидов с использованием этих подходов. Используя свойства оптимизации сфероидов, концентрации соединений и время цикла измерения, основанные на данных, представленных на рисунке 3, рисунке 4 и рисунке 5, был установлен валидированный тест MitoStress для зондирования конкретных параметров митохондриального окислительного метаболизма в диапазоне 3D-сфероидов, полученных из рака (рисунок 6). Важно, как и в некоторых монослойных линиях раковых клеток40, максимальная дыхательная способность (скорость несвязанного-стимулированного дыхания) некоторых 3D-сфероидов, полученных из рака, ингибировалась олигомицином (рисунок 5). В частности, 3D-сфероиды, выращенные из клеток A549 или MCF-7, показали значительно более низкую максимальную скорость дыхания при отделении от BAM15 после инъекции олигомицина по сравнению с разъединением BAM15 без олигомицина (рисунок 5F и рисунок 5H). Учитывая, что этот эффект может присутствовать в других 3D-сфероидных культурах, мы предполагаем, что, если не используется ранее проверенный протокол, максимальная дыхательная способность в 3D-сфероидах должна быть оценена без олигомицина.
Одновременный сбор данных ECAR как мера гликолитического потока в клеточных сфероидах
Как обычно видно из литературы или информации от производителя технологии, гликолитическая скорость сфероидов, измеряемая как ECAR, является вторичным параметром, который может быть захвачен вместе с OCR. Расчет ECAR сам по себе не является полезным или значимым параметром в любом эксперименте XF, поскольку он не корректируется на буферную способность буфера анализа XF или добавление митохондриального подкисления, которое возникает в результате гидратации CO2 до HCO3- и H+. ECAR становится проницательным только после того, как эти поправки данных применяются, после чего становится возможным предоставить более точные выводы о гликолитическом потоке. Чтобы скорректировать буферную способность генерировать более значимые данные PER, необходимо знать объем микрокамеры для сфероидной микропластины. Производитель не смог предоставить истинный объем для этого с помощью сфероидной микропластины, и поэтому данные PER не могут быть легко определены. Действительно, хотя эти измерения могли быть достигнуты эмпирически, это выходило за рамки данной рукописи. Однако при соответствующих поправках и знании объема микрокамеры для данного размера сфероида, присутствующего (например, получение меры плотности сфероида) в скважине, данные ECAR станут значимыми, и можно будет произвести расчеты гликолитического PER. Следовательно, данные XF могут быть более информативными для исследования гликолитического и окислительного метаболизма в сфероидах, но только в том случае, если эти параметры будут рассмотрены углубленно.
Формирование, обработка, перенос и перемещение сфероидов
Некоторые клеточные линии лучше подходят для образования сфероидов, чем другие, и могут вообще не образовывать сфероидов, например, MCF-7 клетки рака яичников41,42 образуют высококруглые сфероиды по сравнению с другими клеточными линиями (рисунок 3). В качестве другого примера, было показано, что раковые клетки поджелудочной железы Capan-1 образуют лучшие сфероиды, чем Panc-1 или BxPC343. Аналогичным образом, клеточные линии печеночной карциномы, как известно, обладают переменной способностью образовывать компактные сфероиды 5,44 с наблюдаемым изменением фенотипа, таким как усиленный метаболизм лекарств или производство альбумина, как в случае HepG2 против HepG2 / C3A 9,45,46 или hepaRG сфероидов 17,47,48 . Поэтому пользователи должны соответствующим образом оптимизировать методы культивирования сфероидов и проводить эксперименты по титрованию для определения оптимальной плотности посева и времени выращивания. Кроме того, было показано, что состав и состав пробирных сред влияют на сфероидную композицию, включая добавление метилцеллюлозы, часто добавляемой в среды для увеличения вязкости матрицы 43,49,50. Следовательно, оптимальный состав клеточной среды должен быть определен эмпирически для всех используемых клеточных линий.
Количество обменов среды по всей сфероидной культуре определяется используемой клеточной линией. Однако, как правило, полуобъемные средние обмены каждые 2-3 дня применимы в большинстве случаев для восполнения питательных веществ. Мы использовали принудительно-агрегатный подход для генерации 3D-сфероидов с использованием клеточно-репеллентных микропластин из коммерчески доступных источников для быстрой разработки и развертывания сфероидных моделей в исследованиях XF-анализов. Однако альтернативные платформы могут быть лучше приспособлены для генерации сфероидов из других типов клеток, например, подходов, висячих или матричных. В лабораториях с ограниченными ресурсами пользователи, возможно, пожелают обратить внимание на метод наложения агарозы-жидкости для формирования поверхностей микропластин с отталкивающими клетками51,52, чтобы значительно снизить экономические затраты на начальные этапы разработки сфероидного метода. Движение сфероидов между сосудами культуры необходимо для выполнения XF-анализа и других последующих анализов. Легкость передачи обычно диктуется размером сфероида и общей плотностью. Мы рекомендуем использовать широкофюзеляжный наконечник пипетки P200 или P1000 для поддержания целостности сфероидов; Наконечники пипеток с меньшим отверстием могут привести к механическому разрушению сфероида, который можно купить в коммерческих целях или, с осторожностью, сделать, просто обрезав конец наконечника пипетки, чтобы увеличить отверстие. Тем не менее, этот подход может привести к загрязнению пластика вокруг конца наконечника, что может привести к механическим нарушениям во время обработки. Использование подсветки или лайтбокса также полезно для обработки сфероидов и наблюдения под рассеченным микроскопом в качестве важного шага для обеспечения успешного переноса сфероидов в микропластину сфероидного анализа. Кроме того, положение сфероида в лунке микропластины сфероидного анализа имеет особое значение и непосредственно влияет на OCR и составной эффект во время типичного теста MitoStress (рисунок 7), скорее всего, из-за взаимосвязи положения сфероида и флуорофоров датчика зонда.
Фоновая коррекция и контроль температуры скважин
Использование анализов на основе микропластин является широко используемым подходом в нескольких областях исследований; однако их использование сопряжено с рядом практических проблем. Как и в других экспериментальных подходах, особенно в тех, которые используют формат массива 96 (или выше), геометрия микропластины и позиционирование могут влиять на температурные и газообменные градиенты по всей пластине с течением времени, часто называемые «краевыми эффектами» 53,54. Мы обнаружили, что то же самое верно и для микропластины сфероидного анализа. В соответствии с руководящими принципами и протоколами производителя, самые внешние угловые скважины: A1, A12, H1 и H12 всегда обозначаются как скважины коррекции фона и контроля температуры для анализатора XFe96. И наоборот, в формате массива из 24 скважин A1 и D6 обозначены как контрольные скважины, наряду с двумя другими скважинами, равномерно распределенными по середине плиты при B4 и C3. При выполнении XF сфероидного анализа мы обнаружили значительные отклонения в данных, первоначально собранных с использованием рекомендаций производителя. И это несмотря на включение необходимых шагов для обеспечения предварительного выравнивания анализа температуры и содержания CO2 до начала сбора данных, что часто приводило к отрицательным значениям OCR после инъекции некоторых респираторных ингибиторов (рисунок 8).
Мы обнаружили, что эти наблюдения, вероятно, связаны с краевыми эффектами через микропластину сфероидного анализа. На рисунке 8 мы обнаружили, что при перераспределении фоновых управляющих скважин по микропластинкам данные XF корректировались примерно в 2 раза. Двумя наиболее вероятными причинами являются (1) эффекты испарения в краевых скважинах, приводящие к уменьшению общей объемной камеры для зонда XFe96 для отбора проб, и (2) недостаточное температурное равновесие между этими скважинами, предназначенными для фоновой коррекции, и пробными скважинами, что приводит к наборам данных, которые либо маскируют, либо чрезмерно раздувают OCR. Поэтому, чтобы избежать таких результатов, рекомендуется, особенно в контексте сфероидного анализа, чтобы пользователи перераспределяли скважины, предназначенные для фоновой коррекции, по всей микропластинке сфероидного анализа и предприняли необходимые шаги для предварительного уравновешивания их анализа до получения данных XF.
Нормализация данных
В дополнение к предоставлению подробного протокола для зондирования митохондриального энергетического метаболизма одиночных 3D-сфероидов с помощью технологии XF, в этой статье также представлены возможные способы нормализации данных о скорости дыхания митохондрий, полученных с помощью 3D-сфероидов. Используя данные о частоте дыхания, полученные с помощью сфероидов MCF-7, культивируемых при различной плотности посева клеток (рисунок 3), мы представляем базальные митохондриальные частоты дыхания из сфероидов MCF-7 увеличивающегося размера и диаметра при нормализации до начальной плотности посева клеток, объема сфероидов и содержания dsDNA (рисунок 9). Соответствующий метод нормализации имеет первостепенное значение для точной интерпретации наборов данных XF, особенно при сравнении 3D-сфероидных моделей in vitro и различных типов клеток. Плохая нормализация может привести к ошибочным результатам, которые просто невозможно сравнить между наборами данных. Содержание белка не является предпочтительным для нормализации данных сфероидного XF, так как предварительная обработка может влиять на скорость синтеза белка без существенного влияния на частоту дыхания. Кроме того, значительные, непоследовательные количества белка могут связываться со сфероидными микропластинами при лизисе клеток, внося изменения в содержание белка между лунками. Это может быть дополнительно осложнено в анализах XF с использованием сфероидов или непригарантных клеток, которые требуют биомолекулярных клеев для связывания, которые могут содержать белок.
В отличие от содержания внутриклеточного белка, содержание ядерной ДНК не зависит от типа клетки и пропорционально числу клеток (рисунок 9D) - более точный и менее трудоемкий подход, чем дезагрегация сфероидов для количественного определения числа клеток. И наоборот, Yepéz et al.55, проведя анализ XF в монослоях клеток фибробластов, обнаружили, что нормализация данных XF к числу клеток приводит к большей дисперсии данных, чем до нормализации. Содержание ядерной ДНК не зависит от дифференцированного состояния или фенотипа и, следовательно, более точно для нормализации сфероидных данных в анализах XF, чем содержание белка. Содержание ДНК также было проверенной стратегией для анализа других наборов данных, связанных с метаболизмом56. Однако важно отметить, что содержание ядерной ДНК количественно определяется всеми клетками, присутствующими в сфероиде; поэтому нормализация содержания ДНК не рекомендуется для наборов данных XF, в которых сфероиды проходят лечение, которое может привести к значительной потере жизнеспособности клеток. Для таких наборов данных, если это возможно, нормализация жизнеспособности клеток является предпочтительной, или данные могут быть скорректированы исходным уровнем до базального дыхания.
Использование запасной дыхательной способности в качестве примера важности нормализации данных
Запасная дыхательная способность является мерой скорости максимальной митохондриальной дыхательной способности минус базальная митохондриальная частота дыхания (рисунок 6). Однако проблема с представлением данных этого типа в виде скорости, т.е. pmolO2/мин/скважина в определенных экспериментах, заключается в том, что данные лишены нормализации. Даже если нормализовать данные сфероидов до плотности клеток / содержания ДНК, это часто исключает ключевой параметр, который необходимо нормализовать для митохондриальной плотности внутри клеток. Учитывая, что изменение плотности митохондрий приведет к пропорциональному изменению базального и максимального дыхания, резервные мощности также увеличатся. Например, еслибазальный OCR сфероида равен 200, а OCRmax равен 400, резервная емкость указывается как 200; еслиOCR базальный равен 100 и OCRmax, то резервная емкость также равна 100; однако в процентном соотношении они оба составляют 50% от максимальных (или 100% от базальных). Таким образом, резервная мощность не изменяется между этими двумя примерами, несмотря на различия в скоростях 200 и 100 при расчете как pmols O2/мин/скважина. Внутренне нормализованные значения являются более надежными и проницательными, чтобы сделать данные XF более сопоставимыми между исследованиями и проектами. Чтобы сделать это для запасной дыхательной способности, мы решили представить это как процент от максимального дыхания, а не абсолютного показателя. Это также может быть представлено в процентах от базального дыхания. Это будет иметь место при работе с клетками или сфероидами. Однако, учитывая, что расположение сфероида в пластине микролунки может изменить абсолютный OCR, но не относительные изменения с ингибиторами или разъединителями, более важно рассматривать внутренне нормализованные ответы в сфероидах как изменение складки или процентное соотношение.
Сфероидные модели, сгенерированные здесь, представляют собой ряд типов клеток и архитектуры, которые не могут быть отражены в классических 2D-моделях. К ним относятся гетерогенное пространственное расположение клеток в трех измерениях, усиленные контакты между клетками (например, образование щелевых соединений и внеклеточных матриц) и биохимические градиенты по всему диаметру сфероида (например, градиенты рН, доступ к питательным веществам для диффузии кислорода). Использование внеклеточного потока для изучения биологии сфероидов in vitro может позволить идентифицировать оптимальные мишени для лекарственной терапии с помощью наблюдений метаболических возмущений. Они могут быть экстраполированы из сфероидов in vitro в опухоли in vivo и идентифицировать пути, которые могут быть нацелены на метаболизм сфероидов и опухолей, например, использование углеводов во время роста сфероидов. Терапевтические методы могут быть эффективными в нацеливании на сфероиды в ранних фазах роста, но оказываются менее эффективными на более поздних фазах роста сфероидов по мере созревания сложности метаболической сети. В заключение, разработка 3D-моделей клеточных культур и сложных технологий анализа в биологических исследованиях будет оставаться динамичной и быстро меняющейся областью с непревзойденным потенциалом. Внеклеточный анализ потоков сфероидов клеточной культуры in vitro может быть использован в качестве передового метода исследования для продвижения результатов исследований, которые могут быть экстраполированы, чтобы лучше понять биологию, связанную с человеком, уменьшить использование животных моделей в исследованиях и улучшить исследования, ориентированные на пациента.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
У авторов нет конфликта интересов, о которых можно было бы заявить.
Acknowledgments
N.J.C был поддержан премией BBSRC MIBTP CASE Award с Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Исследование рака выпуск 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
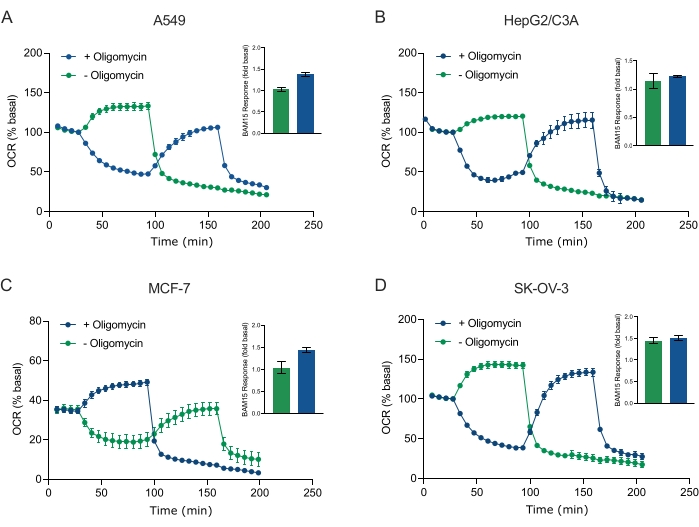
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
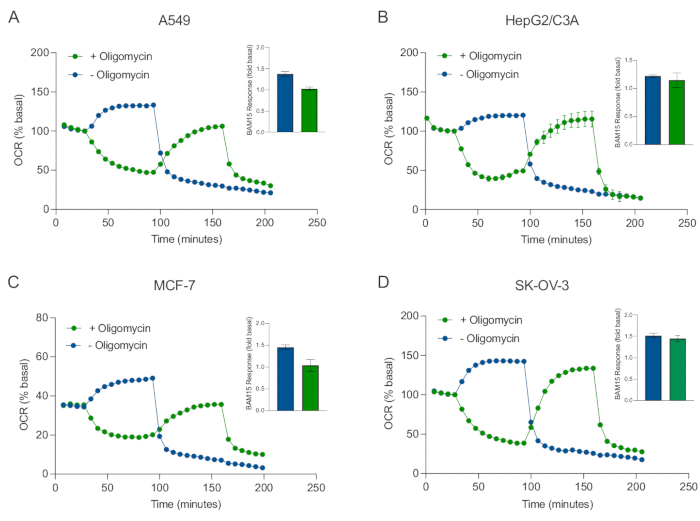
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
