

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Questi protocolli aiuteranno gli utenti a sondare il metabolismo energetico mitocondriale negli sferoidi derivati dalla linea cellulare del cancro 3D utilizzando l'analisi del flusso extracellulare di Cavalluccio marino.
Abstract
Gli aggregati cellulari tridimensionali (3D), chiamati sferoidi, sono diventati l'avanguardia della coltura cellulare in vitro negli ultimi anni. A differenza della coltura delle cellule come monostrati bidimensionali a singola cellula (coltura 2D), la coltura cellulare sferoide promuove, regola e supporta l'architettura cellulare fisiologica e le caratteristiche che esistono in vivo, compresa l'espressione di proteine della matrice extracellulare, la segnalazione cellulare, l'espressione genica, la produzione di proteine, la differenziazione e la proliferazione. L'importanza della cultura 3D è stata riconosciuta in molti campi di ricerca, tra cui oncologia, diabete, biologia delle cellule staminali e ingegneria tissutale. Nell'ultimo decennio, sono stati sviluppati metodi migliorati per produrre sferoidi e valutare la loro funzione metabolica e il loro destino.
Gli analizzatori di flusso extracellulare (XF) sono stati utilizzati per esplorare la funzione mitocondriale in microtessuti 3D come gli sferoidi utilizzando una piastra di cattura delle isole XF24 o una micropiastra sferoide XFe96. Tuttavia, protocolli distinti e l'ottimizzazione del metabolismo energetico mitocondriale di sonda negli sferoidi utilizzando la tecnologia XF non sono stati descritti in dettaglio. Questo documento fornisce protocolli dettagliati per sondare il metabolismo energetico mitocondriale in singoli sferoidi 3D utilizzando micropiastre sferoidi con l'analizzatore XF XFe96. Utilizzando diverse linee cellulari tumorali, la tecnologia XF ha dimostrato di essere in grado di distinguere tra respirazione cellulare in sferoidi 3D non solo di diverse dimensioni, ma anche di diversi volumi, numeri di cellule, contenuto e tipo di DNA.
Le concentrazioni ottimali di composti effettori mitocondriali di oligomicina, BAM15, rotenone e antimicina A vengono utilizzate per sondare parametri specifici del metabolismo energetico mitocondriale negli sferoidi 3D. Questo documento discute anche i metodi per normalizzare i dati ottenuti dagli sferoidi e affronta molte considerazioni che dovrebbero essere prese in considerazione quando si esplora il metabolismo sferoide utilizzando la tecnologia XF. Questo protocollo aiuterà a guidare la ricerca in modelli sferoidi avanzati in vitro .
Introduction
I progressi nei modelli in vitro nella ricerca biologica sono progrediti rapidamente negli ultimi 20 anni. Tali modelli ora includono modalità organ-on-a-chip, organoidi e sferoidi microtesssuei 3D, che sono diventati tutti un obiettivo comune per migliorare la traduzione tra studi in vitro e in vivo. L'uso di modelli avanzati in vitro, in particolare sferoidi, abbraccia diversi campi di ricerca, tra cui l'ingegneria tissutale, la ricerca sulle cellule staminali, il cancro e la biologia delle malattie 1,2,3,4,5,6,7 e i test di sicurezza, tra cui la tossicologia genetica 8,9,10, la tossicologia dei nanomateriali11, 12,13,14 e test di sicurezza ed efficacia dei farmaci 8,15,16,17,18,19.
La normale morfologia cellulare è fondamentale per il fenotipo e l'attività biologica. La coltivazione di cellule in sferoidi microtissuei 3D consente alle cellule di adottare una morfologia, una funzione fenotipica e un'architettura, più simile a quella osservata in vivo ma difficile da catturare con le classiche tecniche di coltura cellulare monostrato. Sia in vivo che in vitro, la funzione cellulare è direttamente influenzata dal microambiente cellulare, che non si limita alla comunicazione e alla programmazione cellulare (ad esempio, formazioni di giunzione cellula-cellula, opportunità di formare nicchie cellulari); esposizione cellulare agli ormoni e ai fattori di crescita negli ambienti immediati (ad esempio, esposizione cellulare alle citochine come parte di una risposta infiammatoria); composizione di matrici fisiche e chimiche (ad esempio, se le cellule vengono coltivate in plastica di coltura tissutale rigida o in un ambiente di tessuto elastico); e, soprattutto, come il metabolismo cellulare è influenzato dalla nutrizione e dall'accesso all'ossigeno, nonché dalla lavorazione di prodotti di scarto metabolici come l'acido lattico.
L'analisi del flusso metabolico è un modo potente per esaminare il metabolismo cellulare all'interno di sistemi in vitro definiti. In particolare, la tecnologia XF consente l'analisi di cambiamenti vivi e in tempo reale nella bioenergetica cellulare di cellule e tessuti intatti. Dato che molti eventi metabolici intracellulari si verificano nell'ordine di secondi o minuti, gli approcci funzionali in tempo reale sono fondamentali per comprendere i cambiamenti in tempo reale nel flusso metabolico cellulare in cellule e tessuti intatti in vitro.
Questo documento fornisce protocolli per la coltivazione di linee cellulari derivate dal cancro A549 (adenocarcinoma polmonare), HepG2 / C3A (carcinoma epatocellulare), MCF-7 (adenocarcinoma mammario) e SK-OV-3 (adenocarcinoma ovarico) come modelli sferoidi 3D in vitro utilizzando approcci ad aggregazione forzata (Figura 1). Inoltre (i) descrive in dettaglio come sondare il metabolismo energetico mitocondriale di singoli sferoidi 3D utilizzando l'analizzatore Agilent XFe96 XF, (ii) evidenzia i modi per ottimizzare i saggi XF utilizzando singoli sferoidi 3D e (iii) discute importanti considerazioni e limitazioni del sondaggio del metabolismo sferoide 3D utilizzando questo approccio. Ancora più importante, questo documento descrive come vengono raccolti i set di dati che consentono il calcolo del tasso di consumo di ossigeno (OCR) per determinare la fosforilazione ossidativa e quindi la funzione mitocondriale negli sferoidi cellulari. Sebbene non sia stato analizzato per questo protocollo, il tasso di acidificazione extracellulare (ECAR) è un altro parametro che viene misurato insieme ai dati OCR negli esperimenti XF. Tuttavia, ECAR è spesso interpretato in modo errato o errato dai set di dati XF. Forniamo un commento sui limiti del calcolo dell'ECAR seguendo gli approcci di base del produttore della tecnologia.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figura 1: Flusso di lavoro grafico per la generazione di sferoidi cellulari, analisi del flusso extracellulare e saggi a valle. Quattro linee cellulari tumorali sono state coltivate selettivamente come monostrati (A), staccate dai palloni di coltura tissutale e seminate in micropiastre a 96 pozzetti di attacco ultrabasso per formare sferoidi (B). Il carcinoma polmonare A549, il carcinoma epatico HepG2 / C3A, l'adenocarcinoma ovarico SK-OV-3 e le cellule di carcinoma mammario MCF-7 sono state seminate a 1 × 103-8 × 103 cellule / pozzetto e cresciute fino a 7 giorni per formare singoli sferoidi e ottimizzare la densità di semina degli sferoidi e il tempo di coltivazione mediante osservazione continua e misurazioni planimetriche. Una volta formati, i singoli sferoidi sono stati lavati in un mezzo XF privo di siero e accuratamente seminati in micropiastre per il dosaggio sferoidale, precolate con poli-D-lisina (C). Gli sferoidi sono stati sottoposti ad analisi del flusso extracellulare utilizzando l'analizzatore XFe96 utilizzando diversi protocolli per affrontare: (1) dimensioni sferoidi ottimali per la risposta respiratoria mitocondriale basale; (2) titolazione ottimizzata degli inibitori respiratori mitocondriali; (3) ottimizzazione del posizionamento sferoidale all'interno di pozzetti di micropiastre. (D) Le analisi post XF, la microscopia a contrasto di fase e la quantificazione del DNA sferoide sono state utilizzate per la normalizzazione dei dati e altri saggi in vitro a valle. Fare clic qui per visualizzare una versione più grande di questa figura.
1. Coltivazione di linee cellulari tumorali come sferoidi 3D in vitro
| Linea cellulare | Descrizione | Terreno di coltura | Fonte |
| A549 · | Linea cellulare di carcinoma polmonare | RPMI 1640 · | Collezione europea di colture cellulari autenticate (ECACC) |
| Piruvato di sodio (1 mM) | |||
| Penicillina- Streptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Linea cellulare del carcinoma epatico, un derivato clonale della linea cellulare HepG2 madre | DMEM · | American Tissue Culture Collection (ATCC) |
| Penicillina- Streptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| MCF7 · | Linea cellulare di adenocarcinoma mammario | RPMI 1640 · | Collezione europea di colture cellulari autenticate (ECACC) |
| Piruvato di sodio (1 mM) | |||
| Penicillina- Streptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 · | Linea cellulare di adenocarcinoma ovarico | RPMI 1640 · | Collezione europea di colture cellulari autenticate (ECACC) |
| Piruvato di sodio (1 mM) | |||
| Penicillina- Streptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| Componente | Mezzo di saggio RPMI (volume finale di 50 ml) | ||
| Base Media | Agilent Cavalluccio marino XF RPMI, pH 7,4 | ||
| Glucosio (1 M di brodo sterile) | 11 mM (0,55 mL di soluzione madre) | ||
| L-glutammina (200 mM di stock sterile) | 2 mM (0,5 mL di soluzione madre) | ||
| Piruvato di sodio (100 mM di stock sterile) | 1 mM (0,5 mL di soluzione madre) |
Tabella 1: Supporti della linea cellulare tumorale e composizioni dei media XF.
- Coltivare tutte le linee cellulari utilizzando la tecnica standard di coltura tissutale asettica e confermare che sono prive di micoplasma utilizzando un kit di analisi adatto.
- Coltivare le linee cellulari in palloni di coltura tissutale T75 o equivalenti, utilizzando il mezzo raccomandato (Tabella 1). Coltivare le linee cellulari al 65-80% di confluenza e passarle regolarmente fino ad un massimo di 25 passaggi.
- Risciacquare due volte i palloni di coltura cellulare nella soluzione salina tamponata con fosfato modificato (DBPS) di Dulbecco.
- Staccare le cellule dai palloni con 3 mL del reagente di dissociazione cellulare (vedere la Tabella dei materiali) per 5 minuti a 37 °C e confermare il distacco al microscopio.
- Aspirare delicatamente la sospensione cellulare staccata per garantire una sospensione unicellulare e disattivare il reagente di dissociazione cellulare con 7 mL di terreno di coltura tissutale completo.
- Raccogliere le cellule per centrifugazione a 300 × g per 5 minuti, scartare il surnatante e risospescere le cellule in mezzo completo.
- Contare le cellule utilizzando un emocitometro o un contatore cellulare automatizzato e titolare la densità cellulare desiderata necessaria per la semina.
NOTA: Per seminare un'intera piastra a 96 pozzetti a 100 μL/pozzetto a 4 × 103 cellule/pozzetto, le cellule devono essere titolate a 4 × 104 cellule/mL in un volume raccomandato di 12 mL. - Decantare la sospensione cellulare in un serbatoio sterile ed erogare 100 μL della sospensione cellulare in ciascun pozzetto di una micropiastra repellente per cellule utilizzando un pipettor multicanale.
NOTA: solo i 60 pozzetti interni di una micropiastra devono essere seminati e il resto riempito con DPBS. Ciò formerà una barriera di evaporazione, garantirà l'omogeneità sferoide attraverso la piastra e ridurrà al minimo gli effetti del bordo della piastra. - Centrifugare micropiastre sferoidi a 300 × g per 15 minuti per forzare le cellule in aggregati sciolti.
- Incubare le piastre a 37 °C, 5% CO2 per un minimo di 3 giorni per garantire la formazione di sferoidi.
- Eseguire la microscopia a contrasto di fase utilizzando pratiche di laboratorio standardizzate per monitorare la crescita degli sferoidi. Ricostituire il terreno di coltura cellulare ogni 3 giorni o due volte alla settimana eseguendo uno scambio di mezzo mezzo volume.
2. Sondare il metabolismo energetico mitocondriale di singoli sferoidi utilizzando la tecnologia Extracellular Flux (XF)
- Preparazione del test (un giorno prima)
- Controllare la vitalità sferoidale utilizzando un microscopio a luce invertita con contrasto di fase con ingrandimento 4x per garantire la struttura sferoide intatta, la morfologia e l'uniformità generale tra i campioni.
- Idratare la cartuccia del sensore.
- Aliquota ~20 mL del calibrante in un tubo conico.
- Posizionare il tubo conico contenente il calibrante in un incubatore non CO2 a 37 °C durante la notte.
- Rimuovere il contenuto dal kit di analisi.
- Rimuovere la cartuccia del sensore dalla piastra di utilità e posizionarla a testa in giù sul piano di lavoro accanto alla piastra di utilità.
- Pipettare 200 μL di ddH2O sterile in ciascun pozzetto della piastra di utilità della cartuccia del sensore utilizzando una pipetta P300 multicanale.
- Posizionare la cartuccia del sensore sulla parte superiore della piastra di utilità.
- Verificare che il livello dell'acqua in ciascun pozzo sia sufficientemente alto da immergere le sonde del sensore.
- Trasferire la cartuccia del sensore assemblata in un incubatore non CO2 a 37 °C e lasciarla per una notte.
NOTA: questo passaggio può essere eseguito 12-72 ore prima dell'inizio del test.
- Micropiastra per analisi sferoide del mantello
- Utilizzando tecniche asettiche, aggiungere 30 μL/pozzetto di soluzione sterile di Poli-D-Lisina (0,1 mg/mL) alla micropiastra sferoide e incubarla per 30 minuti a temperatura ambiente.
- Aspirare la soluzione da ciascun pozzetto della micropiastra sferoidale, invertire la piastra e picchiettarla saldamente sulla carta velina per rimuovere qualsiasi soluzione residua.
- Lavare la piastra due volte con 200 μL/pozzetto di ddH2O sterile.
- Dopo il lavaggio finale, capovolgere la micropiastra e picchiettarla saldamente sulla carta velina per rimuovere l'acqua residua.
- Lasciare asciugare la piastra all'aria per 30 minuti prima di utilizzarla o conservarla a 4 °C per un uso futuro.
NOTA: La micropiastra per il dosaggio dello sferoide deve essere rivestita con un adesivo molecolare per garantire che gli sferoidi siano fissati sul fondo della micropiastra. Senza un adesivo molecolare, gli sferoidi possono essere rimossi e interferire con i risultati del test. Altri adesivi molecolari possono anche essere utilizzati come alternativa alla poli-D-lisina per le piastre di precoating. Le piastre precoate possono essere conservate a 4 °C, ma devono essere lasciate in equilibrio a temperatura ambiente prima dell'inizio del test.
- Preparare il mezzo di analisi XF
- Preparare il mezzo RPMI XF, come descritto nella Tabella 1, e il filtro sterile con un filtro a siringa da 0,22 μm
- Preparazione del test (1 ora prima del test)
- Preriscaldare il mezzo di saggio XF RPMI integrato a 37 °C.
- Preriscaldare la micropiastra di analisi sferoidale rivestita in un incubatore non CO2 a 37 °C o in un bagno secco.
- Preparare la cartuccia del sensore.
- Estrarre il tubo conico contenente il calibrante e la cartuccia del sensore dall'incubatore ad aria.
- Rimuovere la cartuccia del sensore dalla piastra di utilità e posizionarla a testa in giù sul piano di lavoro.
- Utilizzando una pipetta multicanale P300, aspirare l'acqua dalla piastra di servizio e scartarla.
- Versare la soluzione di calibrante in un serbatoio di reagente sterile e aggiungere 200 μL/pozzetto del calibrante preriscaldato alla piastra di servizio utilizzando una pipetta multicanale P300.
- Prelevare la cartuccia del sensore e riposizionarla sopra la piastra di utilità, assicurandosi che i sensori siano ben immersi nel calibrante.
- Trasferire nuovamente la cartuccia del sensore assemblata nell'incubatore non CO2 a 37 °C fino a quando non sarà pronta per caricare le soluzioni di iniezione della porta.
- Lavare gli sferoidi con il mezzo di analisi.
- Rimuovere la piastra di coltura sferoidale dall'incubatore a 37 °C, 5% CO2 e osservare gli sferoidi al microscopio per garantirne l'integrità prima delle fasi di trasferimento dello sferoide.
- Caricare tutti i pozzetti della piastra sferoidale con 180 μL/pozzetto di terreno di saggio preriscaldato, compresi eventuali pozzetti di correzione dello sfondo.
- Riempire parzialmente una capsula di Petri di 7 cm con 3 ml del mezzo di saggio.
- Utilizzando una pipetta multicanale caricata con ampie punte di pipetta orifizio, trasferire gli sferoidi dalla piastra di coltura a 96 pozzetti in piastre di Petri da 7 cm impostando il pipettor a un volume di aspirazione di 10-50 μL.
- Sferoidi di semi nella micropiastra di analisi sferoidale pre-rivestita.
- Utilizzando un microscopio di dissezione e un apparecchio lightbox, trasferire gli sferoidi dalla capsula di Petri alla micropiastra di saggio sferoidale come descritto di seguito.
- Impostare il volume di un pipettor a canale singolo dotato di un'ampia punta della pipetta dell'orifizio a 20 μL e aspirare accuratamente un singolo sferoide. Posizionare la punta direttamente al centro di ciascun pozzetto della micropiastra di saggio sferoidale e consentire alla gravità di eluire un singolo sferoide nel centro di ciascun pozzetto, ovvero non espellere alcun mezzo dalla punta della pipetta e consentire l'azione capillare per prelevare lo sferoide dalla punta della pipetta. Per confermare l'eluizione, il contenuto del pipettor può essere reintubato nella capsula di Petri da 7 cm al microscopio.
NOTA: l'eluizione gravitazionale di un singolo sferoide richiede in genere 15-30 s a seconda delle dimensioni / densità dello sferoide. Durante questo periodo, il pipettor non deve essere rimosso. Qualsiasi pozzetto di correzione dello sfondo deve essere privo di sferoidi e contenere solo un mezzo di analisi. Al microscopio, confermare la posizione di ciascun sferoide. Ogni sferoide dovrebbe idealmente essere posizionato all'interno del centro di ciascun pozzetto. - Una volta che tutti gli sferoidi sono stati trasferiti nella micropiastra di analisi sferoidale, trasferire la piastra in un incubatore non CO2 a 37 °C per un minimo di 1 ora prima del test.
- Impostare il volume di un pipettor a canale singolo dotato di un'ampia punta della pipetta dell'orifizio a 20 μL e aspirare accuratamente un singolo sferoide. Posizionare la punta direttamente al centro di ciascun pozzetto della micropiastra di saggio sferoidale e consentire alla gravità di eluire un singolo sferoide nel centro di ciascun pozzetto, ovvero non espellere alcun mezzo dalla punta della pipetta e consentire l'azione capillare per prelevare lo sferoide dalla punta della pipetta. Per confermare l'eluizione, il contenuto del pipettor può essere reintubato nella capsula di Petri da 7 cm al microscopio.
- Utilizzando un microscopio di dissezione e un apparecchio lightbox, trasferire gli sferoidi dalla capsula di Petri alla micropiastra di saggio sferoidale come descritto di seguito.
3. Preparazione e caricamento dei composti nella cartuccia del sensore per i saggi XF
| Strategia di iniezione | Compound (Porto) | Volume iniziale del microwell XFe96 (μL) | Concentrazione finale del pozzo desiderata | Volume della porta (μL) | Volume finale del microwell XFe96 post iniezione (μL) | Concentrazione delle scorte di lavoro |
| 1 | Oligomicina (A) | 180 | 3 ug/mL | 20 | 200 | 30 μg/mL |
| Rotenone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabella 2: Concentrazioni di composti mitocondriali per sondare il metabolismo energetico mitocondriale di singoli sferoidi 3D utilizzando l'analizzatore XFe96.
- Preparare le concentrazioni di magazzino di ciascun composto come indicato nella Tabella 2 utilizzando un mezzo di saggio RPMI XF completamente integrato e preriscaldato.
- Orientare la piastra della cartuccia (accoppiata alla piastra di utilità) in base alla colonna, 1-12 da sinistra a destra.
- Se si utilizza una guida di carico, posizionarla sopra la piastra della cartuccia in base alla procedura di caricamento del pozzo, ad esempio, se la porta A viene caricata per prima, assicurarsi che A sia visibile nell'angolo in alto a sinistra della guida.
- Trasferire la soluzione di lavoro di ciascun composto in un serbatoio adatto e, utilizzando una pipetta multicanale P100 calibrata, erogare 20 μL in tutte le porte corrispondenti. Ripetere l'operazione per ogni composto nelle porte rimanenti.
NOTA: se non vengono utilizzate porte sulla piastra della cartuccia del sensore, queste possono essere lasciate vuote o riempite con il supporto di analisi. Se viene utilizzata solo una selezione di una specifica lettera di porta, assicurarsi che le altre porte corrispondenti a tale lettera siano caricate con un supporto di analisi; in caso contrario, l'aria verrà iniettata nel pozzo, compromettendo i risultati in quei pozzi. - Dopo il caricamento della porta, rimuovere le guide di caricamento delle piastre (se utilizzate) e preparare l'analizzatore per il caricamento della cartuccia del sensore.
NOTA: se il test non viene eseguito immediatamente dopo aver caricato le porte, riposizionare il coperchio sulla cartuccia del sensore e rimettere la piastra nell'incubatore ad aria a 37 °C fino a quando non sarà pronta per essere caricata nella macchina.
4. Progettazione del test, strategie di iniezione e acquisizione dati
- Esecuzione del test
- Accendere l'analizzatore e collegarlo al controller (computer).
NOTA: questo può essere verificato dallo stato della connessione dello strumento nel pannello widget del software Wave Controller. - Passare alla pagina dei modelli nel software WAVE, trovare il file del modello di analisi per l'esperimento e fare doppio clic per aprirlo.
NOTA: se il modello di analisi non viene visualizzato nella vista Modelli , importare il file modello nella cartella del modello da un'unità di rete condivisa o da un'unità flash USB. - Per avviare il test, fare clic sulla scheda Esegui test .
NOTA: se le definizioni di gruppo sono state allocate correttamente all'interno della mappa della piastra, il test sarà pronto per essere eseguito come indicato dal segno di spunta verde sul lato destro della pagina. In questa fase, qualsiasi informazione aggiuntiva può essere inserita nella pagina di riepilogo del test o nella pagina lasciata vuota; procedere al passaggio successivo. A causa della penetrazione ritardata dei modulatori mitocondriali negli sferoidi microtissue 3D (Figura 2), utilizzare le informazioni sul protocollo di misurazione descritte nella Tabella 3.
- Accendere l'analizzatore e collegarlo al controller (computer).
| Periodo di misurazione | Numero di iniezione e porta | Dettagli di misurazione | Durata del periodo (h:min:s) |
| Taratura | Non applicabile | Gli analizzatori XF eseguono sempre questa calibrazione per assicurarsi che le misurazioni siano accurate | 00:20:00 (questa è una media e può variare tra le macchine) |
| Equillibrazione | Non applicabile | L'equilibrio avviene dopo la calibrazione ed è raccomandato. | 00:10:00 |
| Basale | Non applicabile | Cicli = 5 | 00:30:00 |
| Mix = 3:00 | |||
| Attendi = 0:00 | |||
| Misura = 3:00 | |||
| Oligomicina / BAM15 | Iniezione 1 (Porta A) | Cicli = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Attendi = 0:00 | |||
| Misura = 3:00 | |||
| Rotenone + antimicina A | Iniezione 2 (Porta B) | Cicli = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Attendi = 0:00 | |||
| Misura = 3:00 | |||
| Tempo totale: | 03:00:00 | ||
Tabella 3: Configurazione del protocollo per sondare il metabolismo energetico mitocondriale di singoli sferoidi 3D utilizzando l'analizzatore XFe96.
- Fare clic su Avvia esecuzione per visualizzare la finestra di dialogo Salva percorso .
- Immettere la posizione di salvataggio per il file dei risultati e posizionare la cartuccia del sensore assemblata sul vassoio termico che appare dalla porta sul lato dell'analizzatore. Attendere che il vassoio termico si apra automaticamente e che sullo schermo venga visualizzato il messaggio Load Calibrant Utility Plate . Prima di seguire le istruzioni visualizzate sullo schermo, assicurarsi i) il corretto adattamento della cartuccia del sensore sulla piastra dell'utilità, ii) che il coperchio venga rimosso dalla cartuccia del sensore e iii) il corretto orientamento della cartuccia del sensore sulla piastra dell'utilità.
- Seguire i comandi sullo schermo per avviare la calibrazione della cartuccia del sensore.
NOTA: Il tempo necessario per completare la calibrazione è di circa 10-20 minuti (per saggi a 37 °C). - Dopo la calibrazione della cartuccia del sensore, caricare la micropiastra sferoide nell'analizzatore seguendo le istruzioni visualizzate sullo schermo sul Wave Controller per avviare la fase di bilanciamento di 12 minuti.
NOTA: le caselle verdi con segni di spunta bianchi indicano una calibrazione "buona" per quel pozzo. Se alcuni pozzi non riescono a fornire una calibrazione "buona", saranno indicati con una scatola rossa e una croce bianca. Tali pozzi devono essere annotati ed esclusi da qualsiasi analisi dopo che il test è stato completato utilizzando la scheda del test di modifica . - Attendere che l'analizzatore inizi automaticamente ad acquisire le misurazioni di base dopo che la macchina ha completato la fase di equilibratura (come indicato nel protocollo dello strumento).
- Per completare l'esperimento, seguire i comandi sullo schermo sul controller WAVE.
NOTA: una volta che la micropiastra sferoidale è stata rimossa dall'analizzatore, scartare la cartuccia del sensore e mettere da parte la piastra sferoide per ulteriori analisi, se necessario (ad esempio, quantificazione del DNA a doppio filamento (ds).) Se la micropiastra non è necessaria per ulteriori analisi, può essere scartata insieme alla cartuccia del sensore. - Attendere la visualizzazione della finestra di dialogo del test e visualizzare i risultati o tornare alla visualizzazione dei modelli .
5. Strategie di normalizzazione e analisi dei dati - normalizzazione post-test e saggi a valle (passaggi facoltativi)
- Normalizzazione dei dati
- Per normalizzare i dati sferoidi, fare riferimento alla serie di protocolli pertinenti alle strategie di normalizzazione dei dati per il calcolo delle dimensioni e del volume degli sferoidi e la quantificazione del dsDNA nei saggi sferoidi. Questi sono stati inclusi come file supplementari; vedere Il file supplementare 1 e il file supplementare 2.
- Analisi dei dati
- Per esportare i dati in uno dei generatori di analisi automatizzati, seguire i comandi di esportazione dei dati sul controller WAVE e selezionare il generatore di esportazione che corrisponde al tipo di test. In alternativa, esporta il file di dati e caricalo in Seahorse Analytics.
NOTA: lo svantaggio dei generatori di report e dell'analisi Seahorse è che l'analisi dei dati è limitata al modo in cui è progettato il test XF e non consente di prendere le medie tra i cicli di misurazione. L'esportazione manuale dei set di dati dal software dello strumento consente la preferenza dell'utente a questo proposito. Dato che la strategia di iniezione per valutare la respirazione mitocondriale degli sferoidi 3D sarà probabilmente diversa da quella di un tipico test "MitoStress", sono stati sviluppati una serie di modelli di fogli di calcolo per aiutare ad analizzare questi set di dati, specifici per le colture cellulari 3D e saranno forniti su richiesta. Questi file modello di dati forniranno dati sui principali parametri respiratori mitocondriali dettagliati e spiegati nella Figura 2. - Per analizzare i dati, esportare i dati come report del foglio di calcolo dal software del controller WAVE e utilizzare un modello di foglio di calcolo indipendente per l'analisi.
- Per esportare i dati in uno dei generatori di analisi automatizzati, seguire i comandi di esportazione dei dati sul controller WAVE e selezionare il generatore di esportazione che corrisponde al tipo di test. In alternativa, esporta il file di dati e caricalo in Seahorse Analytics.

Figura 2: Descrittori schematici per parametri derivati da analisi di dati di flusso extracellulare. Abbreviazione: OCR = tasso di consumo di ossigeno. Fare clic qui per visualizzare una versione più grande di questa figura.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Per ottenere sferoidi compatti e ben formati, ogni linea cellulare è stata ottimizzata individualmente per la densità di semina e la durata della coltivazione (Figura 3). Le linee cellulari A549, HepG2/C3A e SK-OV-3 inizialmente formavano aggregati sciolti che non progredivano verso sferoidi rotondi con perimetri chiaramente definiti fino a dopo 7 giorni in coltura. Al contrario, le cellule MCF-7 potrebbero formare sferoidi entro 3 giorni. C'era una chiara correlazione tra la densità iniziale di semina cellulare e il volume sferoide dopo il periodo di coltura per tutti i modelli sferoidi. Le dimensioni e la morfologia degli sferoidi sono state ottimizzate per la densità di semina. La morfologia e la circolarità iniziarono a diminuire con l'aumento delle dimensioni sferoidi in tutti i modelli. Le strategie di semina per le linee cellulari sono state ottimizzate a 4 × 103 celle/pozzetto per le cellule A549 e SK-OV-3; Le cellule HepG2 / C3A sono state precedentemente ottimizzate altrove a 1 × 103 cellule / pozzetto, e le cellule MCF-7 sono state utilizzate a 4 × 103 cellule / pozzetto in tutti i test. A strategie di semina ottimizzate, il volume sferoide era compreso tra 5,46 × 107 μm3 (SK-OV-3) e 1,45 × 108 μm3 (A549) (Figura 3B). Tutti i tipi di sferoidi avevano una correlazione lineare tra la densità di semina iniziale e il volume sferoide dove A549 e HepG2 / C3A avevano valori R2 di 0,957 e 0,947, rispettivamente. I volumi sferoidi MCF-7 e SK-OV-3 sono risultati entrambi avere una maggiore correlazione con la densità di semina iniziale, R2 = 0,977 (Figura 3A).
La circolarità sferoidale è stata calcolata utilizzando la planimetria dell'immagine all'interno del software di analisi FIJI utilizzando i diametri sferoidi lunghi e corti. La simmetria sferoide perfetta aveva circolarità = 1,0; la deviazione da 1,0 indicava una perdita di circolarità (Figura 3C). La circolarità era maggiore negli sferoidi MCF-7 rispetto ad altri modelli in cui la circolarità era mantenuta tra 0,83 e 0,9 a tutte le densità di semina. In confronto, il perimetro esterno degli sferoidi SK-OV-3 non era così chiaramente definito e il volume sferoide era significativamente più piccolo anche dopo 7 giorni in coltura, producendo sferoidi con una circolarità massima di 0,61 a una densità di semina di 4 × 103 / pozzetto. Si è anche scoperto che le cellule HepG2 / C3A formano sferoidi stretti e ben formati con una morfologia uniforme attraverso la superficie di tutti gli sferoidi, con circolarità mantenuta a 0,79 per le cellule seminate a 1 × 103 cellule / pozzetto. Le cellule A549 sembravano seguire una tendenza in cui la circolarità e la morfologia sferoide erano migliorate con la densità di semina; tuttavia, la circolarità non era superiore a 0,63 alla densità utilizzata in questi esperimenti.
La respirazione mitocondriale basale è stata calcolata come OCR misurata da sferoidi seminati a 1 × 103, 2 × 103, 4 × 103, o 8 × 103 cellule / pozzetto in micropiastre di coltura sferoide ad attacco ultrabasso (Figura 3D). Per tutti i tipi di sferoidi, l'OCR è aumentato con la dimensione dello sferoide ed è stato linearmente correlato al volume sferoide con R2 più alto negli sferoidi MCF-7 a 0,988 e più basso negli sferoidi SK-OV-3 a 0,744 (Figura 3E). L'OCR misurato era statisticamente diverso tra tutti i gruppi sperimentali. A549 aveva l'OCR più basso, raggiungendo solo 18 pmol / min / well alla dimensione sferoide più grande (Figura 3D). Al contrario, gli sferoidi MCF-7 hanno prodotto un OCR simile alla dimensione sferoide più piccola dopo soli 3 giorni in coltura, raggiungendo un OCR basale massimo di 53 pmol / min / pozzetto per la più grande dimensione sferoide (Figura 3D). HepG2/C3A ha prodotto dati OCR altamente coerenti con le dimensioni e la morfologia degli sferoidi. Negli sferoidi HepG2/C3A seminati da 1 × 103 cellule/pozzetto, l'OCR basale ha raggiunto una media di 15 pmol/min/well, aumentando fino ad un massimo di 52 pmol/min/well negli sferoidi più grandi (Figura 3D). L'OCR negli sferoidi SK-OV-3 era significativo solo tra gli sferoidi cresciuti da 4 × 103 cellule / pozzetto e 8 × 103 cellule / pozzetto, con poca differenza osservata negli sferoidi cresciuti tra 1 × 103, 2 × 103 o 4 × 103 cellule. Nonostante le differenze di dimensioni, i dati OCR erano molto simili tra gli sferoidi HepG2 / C3A e MCF-7 in tutti i punti di dimensione. Rispetto alla dimensione degli sferoidi (μm3), l'OCR basale degli sferoidi MCF-7 era paragonabile a quello degli sferoidi HepG2/ C3A cresciuti in 7 giorni da 1.000 cellule per pozzetto.

Figura 3: I parametri di crescita sferoidi determinano la respirazione mitocondriale al basale. (A) Gli sferoidi A549, HepG2/C3A, MCF-7 e SK-OV-3 sono stati ottimizzati per la densità di semina e la loro crescita monitorata a 1 × 103, 2 × 103, 4 × 103 e 8 × 103 cellule/pozzetto in ciascuna delle fotomicrografie, rispettivamente dall'alto a sinistra in basso a destra; barre della scala = 500 μm. (B) La dimensione dello sferoide è stata calcolata utilizzando i dati planimetrici delle fotomicrografie raccolte e confrontata utilizzando la statistica di correlazione di Pearson; le linee tratteggiate rappresentano la distribuzione dell'intervallo di confidenza del 95%. (C) La morfologia sferoide è stata confrontata mediante il calcolo della circolarità. (D) L'OCR è stato misurato 5 volte, dopo di che il rotenone-antimicina A è stato aggiunto per tenere conto della frequenza respiratoria nonmitocondriale utilizzando l'analizzatore Agilent Seahorse XFe96. OCRbasale misurato come OCR - OCRr/a è stato confrontato tra densità di semina (D) e volume sferoide (E). I dati sono medie ± SEM da 5-8 ben replica per tipo di sferoide e densità di semina cellulare. Abbreviazioni: OCR = tasso di consumo di ossigeno; OCRbasale = Respirazione mitocondriale basale; OCRr/a = OCR dopo l'aggiunta di rotenone-antimicina A. Fare clic qui per visualizzare una versione più grande di questa figura.
La concentrazione e il corso temporale per l'esposizione ai modulatori respiratori nell'analisi XF sono un passo fondamentale nell'ottimizzazione del saggio. I composti modulatori respiratori -oligomicina, BAM15, una miscela di rotenone-antimicina A- o un controllo del veicolo DMSO sono stati iniettati in sequenza attraverso le porte di iniezione della cartuccia del sensore in pozzetti micropiastre contenenti gli sferoidi MCF-7 (Figura 4A). Sono stati completati quattro cicli di misurazione per determinare l'OCR basale medio di 30-40 pmol/min/pozzetto per tutti i gruppi di campioni. Per il resto della durata del test, i modulatori respiratori sono stati aggiunti sequenzialmente ogni 5 cicli di misurazione per raggiungere concentrazioni finali di pozzo di 0,5 μM (0,5 μg/mL di oligomicina) dopo l'iniezione 1; 2,0 μM (2 μg/mL di oligomicina) dopo l'iniezione 2; 5 μM (5 μg/mL di oligomicina) dopo l'iniezione 3; infine, una concentrazione massima di pozzo di 11 μM (11 μg/mL di oligomicina) dopo la quarta addizione sequenziale. Gli sferoidi MCF-7 non hanno risposto al controllo del veicolo durante l'esperimento (Figura 4A). L'OCR basale è cambiato immediatamente dopo la prima iniezione di ciascun rispettivo composto alla concentrazione più bassa di 0,5 μM o 0,5 μg/mL di oligomicina (Figura 4B). L'OCR negli sferoidi MCF-7 è stato abbassato con oligomicina da 41 pmol/min/pozzetto a 23 pmol/min/well dopo 5 cicli di misurazione successivi alla prima iniezione di 0,5 μg/mL (Figura 4B).
In risposta a 0,5 μm BAM15, l'OCR è stato aumentato da 33 a 41 pmol/min/ben prima della seconda iniezione (Figura 4C). Comparativamente, la combinazione di rotenone più antimicina A ha abbassato l'OCR da 37 a 13 pmol/min/ben prima della seconda iniezione (Figura 4D). Le tracce cinetiche hanno inoltre rivelato una costante diminuzione lineare (oligomicina e rotenone-antimicina A) o aumento (BAM15) nell'OCR. Per tutti i regimi di dosaggio composti, è stato ottenuto un OCR allo stato stazionario entro 10-12 cicli di misurazione completi (60-72 min) ad una concentrazione totale di pozzo di 2 μM BAM15, 2 μM rotenone, 2 μM antimicina A e 2 μg/mL di oligomicina (Figura 4A). Il tasso di consumo di ossigeno ha raggiunto un plateau allo stato stazionario a ~ 19 pmol / min / well (oligomicina), 52 pmol / min / well (BAM15) e 10 pmol / min / well (rotenone-antimicina A) (Figura 4A). L'aumento della concentrazione composta di oligomicina, BAM15 o rotenone più antimicina A non ha avuto alcun effetto evidente sull'OCR, che è rimasto costante per tutto il resto del test. Questi dati dimostrano che sia la concentrazione di composti che il corso temporale di esposizione ai composti modulatori respiratori devono essere considerati per l'ottimizzazione del saggio quando si utilizzano sferoidi 3D.

Figura 4: Titolazione dei composti modulatori respiratori come passo importante per l'ottimizzazione dell'analisi del flusso extracellulare. (A) Gli sferoidi MCF-7 sono stati seminati a 4 × 103 cellule / pozzetto e coltivati per 3 giorni prima di essere collocati in pozzetti di una micropiastra di saggio sferoide contenente XF RPMI e sondati per modulatori mitocondriali OCR ± utilizzando l'analizzatore XFe96. L'OCR è stato misurato 5 volte, dopo di che sono state aggiunte titolazioni di controllo del veicolo, oligomicina (B), BAM15 (C) o rotenone-antimicina A (D) per inibire l'ATP sintasi mitocondriale, determinare la capacità respiratoria massima o stabilire la frequenza respiratoria nonmitocondriale, rispettivamente. La concentrazione di ciascun modulatore mitocondriale è stata aumentata in 4 strategie individuali di iniezione di titolazione (0,5 μM, 1,5 μM, 3 μM e 6 μM; le unità per l'oligomicina sono μg / mL) per determinare l'OCR allo stato stazionario massimo in risposta alla concentrazione ottimale del composto. L'OCR è stato misurato per 5 cicli di misurazione tra ogni iniezione. I dati sono medie ± SEM da 5-8 singole repliche di pozzi. Abbreviazione: OCR = tasso di consumo di ossigeno. Fare clic qui per visualizzare una versione più grande di questa figura.
Uno dei principali vantaggi della tecnologia XF è la capacità di sondare la funzione mitocondriale in cellule e tessuti intatti. Per esaminare aspetti specifici della funzione mitocondriale nelle cellule e nei tessuti, i modulatori mitocondriali vengono aggiunti sequenzialmente ai pozzetti della micropiastra campione attraverso le 4 porte di iniezione disponibili sulla cartuccia del sensore. La sequenza tipica di modulatori utilizzati per sondare i parametri mitocondriali nei saggi XF sono l'oligomicina, un protonoforo (ad esempio, FCCP o BAM15) e una combinazione di rotenone più antimicina A, che vengono aggiunti sequenzialmente per inibire l'ATP sintasi mitocondriale, determinare la capacità respiratoria massima e correggere la frequenza respiratoria non quadricondriale, rispettivamente. Questa tipica sequenza di aggiunte di modulatori è definita test MitoStress dal produttore della tecnologia di analisi. Dato che l'oligomicina può inibire la respirazione stimolata da un disaccoppiatori in alcuni monostrati cellulari20, abbiamo esaminato questo con sferoidi 3D derivati dal cancro misurando l'OCR non disaccoppiato-stimolato (OCRmax) prima (singolo) e dopo l'iniezione di oligomicina (sequenziale) (Figura 5A-D). L'OCRmax non è stato significativamente limitato dall'aggiunta di oligomicina negli sferoidi formati da HEPG2/C3A o SK-OV-3 (Figura 5E e Figura 5G). Tuttavia, l'OCRmax è stato significativamente abbassato negli sferoidi A549 e MCF-7 a seguito di un'iniezione sequenziale di BAM15 dopo oligomicina rispetto all'OCRmax ottenuto da una singola iniezione di BAM15 (Figura 5F e Figura 5H). Se non diversamente noto, si raccomanda quindi di utilizzare pozzetti separati per trattare con oligomicina e uncoccoppiatore, con un'aggiunta finale di rotenone e antimicina A quando si esplora il metabolismo energetico mitocondriale degli sferoidi 3D. Questo approccio consente ancora il calcolo di tutti i parametri mitocondriali come con un tipico test MitoStress in cui i composti vengono aggiunti in sequenza.

Figura 5: Iniezione singola o sequenziale di composti respiratori mitocondriali. Gli sferoidi derivati dalle cellule tumorali di MCF-7, HEPG2/C3A, SK-OV-3 e A549 sono stati collocati in pozzetti di una micropiastra sferoide XFe96 in XF RPMI e sondati per OCR utilizzando l'analizzatore Agilent Seahorse XFe96. L'OCR è stato misurato 5x, dopo di che 2 μg/mL di oligomicina (porta di iniezione A: traccia verde) o 5 μM BAM15 (porta di iniezione A: traccia blu o porta di iniezione B: traccia verde) per inibire rispettivamente l'ATP sintasi mitocondriale e determinare la capacità respiratoria massima. I dati OCR cinetici sono espressi in % basale (A-D). La capacità respiratoria massima (OCRmax) è stata calcolata come fattore di OCR basale dall'equazione: OCRmax = OCRBAM15 / OCRbasale. OCRmax è stato ottenuto da medie OCR attraverso cicli di misurazione 8-10 post iniezione BAM15 con (barre verdi) e senza (barre blu) oligomicina. I dati sono medie ± SEM da 3-8 singoli pozzi replica attraverso la micropiastra di saggio sferoidale. Abbreviazioni: OCR = tasso di consumo di ossigeno. Fare clic qui per visualizzare una versione più grande di questa figura.
Utilizzando densità di semina cellulare ottimali, concentrazioni composte, strategia di iniezione e periodo del ciclo di misurazione determinato in questi esperimenti di ottimizzazione (Tabella 3), abbiamo sviluppato un protocollo dettagliato per sondare con precisione la respirazione mitocondriale basale: OCRbasale (Figura 6A), respirazione di fosforilazione ADP: OCRADP (Figura 6B), respirazione delle perdite: OCRomy (Figura 6C), efficienza di accoppiamento (Figura 6D ), capacità respiratoria massima: OCRmax (Figura 6E) e capacità respiratoria di riserva: OCRdi riserva (Figura 6F) utilizzando sferoidi 3D derivati dal cancro.

Figura 6: Sondare l'OCR con la tecnologia XF per stabilire il metabolismo energetico mitocondriale degli sferoidi derivati dal cancro. Gli sferoidi derivati dalle cellule tumorali di MCF-7, HEPG2/C3A, SK-OV-3 e A549 sono stati collocati in pozzetti di una micropiastra di saggio sferoidale in XF RPMI e sondati per OCR utilizzando l'analizzatore Agilent Seahorse XFe96. L'OCR è stato misurato 5x, dopo di che 2 μg/mL di oligomicina, o 5 μM BAM15, e l'AR è stato aggiunto per inibire l'ATP sintasi mitocondriale, determinare la capacità respiratoria massima e calcolare la frequenza respiratoria nonmitocondriale, rispettivamente. (A) La respirazione mitocondriale basale (OCRbasale) è stata calcolata come media dell'OCR dai 3 cicli di misurazione prima dell'iniezione della porta A. (B) L'efficienza di accoppiamento della fosforilazione ossidativa è stata approssimata esprimendo OCRADP (OCRbasal-OCR leak) come percentuale di OCRbasale. (C) La respirazione di fosforilazione ADP (OCRADP) è stata misurata come OCR oligomicina-sensibile, calcolata dall'OCR medio attraverso cicli di misurazione 11-13 prima dell'iniezione di BAM15. (D) La perditadi omy respirazione (perdita OCR) è stata misurata come OCR insensibile all'oligomicina, calcolata dall'OCR medio medio attraverso i cicli di misurazione 11-13. (E) La capacità respiratoria massima (OCRmax) è stata misurata come l'OCR massimo medio misurato dopo l'iniezione di BAM15. (F) La capacità respiratoria di riserva è stata calcolata esprimendo OCRdi riserva (OCRmax - OCRbasale) come percentuale di OCR basale. L'OCR dopo iniezione di rotenone-antimicina A (OCRr/a) è stata sottratta da tutte le velocità per correggere l'OCR nonmitocondriale. I dati sono medie ± SEM da 3-8 singoli pozzi replicati attraverso la piastra sferoidale XFe96. Abbreviazioni: OCR = tasso di consumo di ossigeno; RA = 2 μM rotenone-2 μM antimicina A. Fare clic qui per visualizzare una versione più grande di questa figura.
Gli sferoidi MCF-7 cresciuti da 4 × 103 cellule / ben oltre 3 giorni sono stati utilizzati come modello per determinare il trasferimento, il posizionamento e l'analisi ottimali all'interno di micropiastre di saggio sferoidale. Utilizzando le dimensioni fornite per la micropiastra sferoidale dal produttore, la superficie del pozzo è stata divisa in tre aree di zona per un posizionamento sferoide ottimale (Figura 7A), dove la zona 1 è stata evidenziata come la zona ottimale al centro del pozzo. Con un'attenta pipettaggio utilizzando punte di pipette a orifizio largo, gli sferoidi sono stati trasferiti nelle piastre sferoidi e distribuiti casualmente sulle superfici del pozzo mediante eluizione gravitazionale (Figura 7B). Dove gli sferoidi sono stati accuratamente trasferiti utilizzando l'eluizione gravitazionale, la maggior parte degli sferoidi potrebbe in genere essere trovata nelle zone 1-2 della micropiastra, utilizzando le tecniche di trasferimento raccomandate dal produttore. Dove gli sferoidi venivano spinti fuori dalla punta della pipetta per aspirazione, gli sferoidi venivano spesso posizionati oltre queste zone e non potevano essere visti al microscopio.
Per confrontare le posizioni di posizionamento degli sferoidi, gli sferoidi MCF-7 sono stati trasferiti nelle micropiastre del saggio sferoide nelle zone designate 1-3 o fuori zona (Figura 7A). Questi 4 pozzetti sono stati tracciati attraverso un esperimento cinetico OCR al basale e dopo l'aggiunta di oligomicina, BAM15 o rotenone-antimicina A (Figura 7C). L'OCR è stato calcolato sulla base della media delle letture di tre cicli prima di ogni iniezione (Figura 7B). L'OCR è stato misurato cineticamente su 200 minuti nei 4 pozzi selezionati (Figura 7C) e corretto al basale (Figura 7D). Dove gli sferoidi sono stati collocati nella zona 3 o fuori zona, gli OCR al basale erano significativamente inferiori agli sferoidi collocati nelle zone 1 e 2 (Figura 7C). Anche gli effetti dei composti respiratori oligomicina, BAM15 e rotenone-antimicina A differivano notevolmente tra gli sferoidi collocati nelle zone 1 e 2 rispetto alla zona 3 e alle regioni fuori zona. Un aumento dell'OCR è stato osservato con oligomicina in sferoidi collocati nella zona 3 o fuori zona (Figura 7E). Inoltre, gli sferoidi collocati nella zona 3 o fuori zona hanno sperimentato una risposta eccessivamente elevata a BAM15 con OCR superiore al basale dopo l'iniezione di rotenone-antimicina A (Figura 7E). Nonostante un aumento quasi doppio dell'OCR basale (Figura 7C) con sferoidi collocati nella zona 2 rispetto alla zona 1, i cambiamenti di piega in risposta a tutti i composti respiratori erano molto simili (Figura 7E), suggerendo che le differenze nell'OCR basale tra gli sferoidi collocati nelle zone 1 o 2 sono improbabili che siano il risultato del posizionamento all'interno del pozzo.

Figura 7: Il posizionamento degli sferoidi all'interno della micropiastra di saggio sferoidale determina l'OCR basale e gli effetti del modulatore mitocondriale utilizzando la tecnologia XF. Gli sferoidi MCF-7 sono stati seminati a 4 × 103 cellule / pozzetto e coltivati per oltre 3 giorni prima di essere collocati nei pozzetti della microplacca sferoide contenente XF RPMI e sondati per i modulatori mitocondriali OCR ± utilizzando l'analizzatore Agilent Seahorse XFe96. (A) Fotomicrografie delle posizioni delle zone sferoidi nelle micropiastre del saggio sferoidale dopo la durata del saggio; barra di scala = 500 μm e OCR catturati dai pozzi corrispondenti nel tempo espressi come pmol/min-1/well-1 (B) o % basale (C). (D) Effetti modulatori mitocondriali degli sferoidi MCF-7 collocati in zone diverse all'interno della micropiastra di saggio sferoidale; dati espressi come variazione di piega da basale. (E) Esempio di traccia cinetica che evidenzia quali misurazioni dei dati OCR (cerchi rossi) sono utilizzate per calcolare la risposta di ciascun modulatore mitocondriale per i dati presentati in E. I dati mostrati provengono da singole risposte ai pozzi. Abbreviazione: OCR = tasso di consumo di ossigeno. Fare clic qui per visualizzare una versione più grande di questa figura.
I criteri di selezione per lo sfondo sono di grande importanza; l'uso di pozzi più esterni per la correzione di fondo non è rappresentativo di tutti i pozzetti micropiastre, il che può portare a ipotesi di dati errate e conclusioni errate sui dati a causa di effetti di bordo attraverso la microplacca sferoide. Per valutare questa osservazione, gli sferoidi MCF-7 sono stati utilizzati per confrontare le procedure di correzione del saggio per derivare i valori OCR in risposta all'aggiunta di un controllo del veicolo, oligomicina, BAM15 o rotenone-antimicina A (Figura 8). Tutti i composti respiratori hanno prodotto i profili OCR cinetici attesi per i composti selezionati, rivelando un tasso medio di respirazione basale costante di 20-30 pmol/min/well (Figura 8A). Tuttavia, laddove i dati del saggio sono stati analizzati utilizzando i pozzetti più esterni per la correzione della temperatura di fondo (A1, A12, H1 e H12), i valori rivelati per l'OCR dopo l'aggiunta di composti respiratori erano particolarmente bassi; L'OCR ha prodotto valori negativi per il rotenone-antimicina A. In risposta a queste osservazioni, l'analisi alternativa è stata eseguita utilizzando una serie di pozzi vuoti, distribuiti casualmente attraverso la micropiastra sferoide, come pozzi di correzione della temperatura di fondo (Figura 8B). Laddove è stata applicata una correzione alternativa dello sfondo, tutti gli effetti composti relativi sull'OCR erano gli stessi in entrambi i set di analisi; tuttavia, i valori assoluti di OCR sono aumentati di circa 10 pmol/min/well (Figura 8). Questi dati evidenziano la potenza e l'importanza della correzione della temperatura di fondo sulle micropiastre per saggi sferoidi e sottolineano l'importanza dell'ottimizzazione dell'utente per l'analisi XF.

Figura 8: Selezione casuale dei pozzetti per la correzione dello sfondo per migliorare il controllo dei gradienti di temperatura attraverso la micropiastra di saggio sferoidale. Dati OCR estrapolati dalla Figura 2A utilizzando pozzi consigliati per la correzione dello sfondo (A) rispetto a pozzi assegnati casualmente per la correzione dello sfondo (B). Abbreviazione: OCR = tasso di consumo di ossigeno. Fare clic qui per visualizzare una versione più grande di questa figura.
A differenza dei monostrati cellulari, gli sferoidi rappresentano un'aggregazione eterogenea di cellule in uno spazio 3D e quindi richiedono un'attenta considerazione rispetto all'analisi, in particolare quando si normalizzano questi dati. Questo documento presenta tre approcci per normalizzare i dati XF acquisiti dagli sferoidi MCF-7 (Figura 9). Quando non normalizzato, l'OCR correla positivamente (R2 = 0,98) con la dimensione dello sferoide (determinata dalla densità iniziale di semina delle cellule) in modo significativo se confrontato statisticamente con il coefficiente di correlazione di Pearson, P = 0,0057 (Figura 9A). Questa relazione lineare si abbassa quando l'OCR viene normalizzato alla densità iniziale di semina cellulare (R2 = 0,78) e non è più significativamente correlata alla dimensione sferoide (P = 0,117, Figura 9B). Questo è anche il caso quando normalizzato al volume sferoide (R2 = 0,77; Coefficiente di correlazione di Pearson P = 0,120, Figura 9C) e contenuto di dsDNA nucleare (R2 = 0,58; Coefficiente di correlazione di Pearson P= 0,233, Figura 9D). Questi dati evidenziano l'importanza di normalizzare i dati XF quando si sonda il metabolismo mitocondriale degli sferoidi, specialmente se sono di dimensioni diverse.

Figura 9: Normalizzazione dei dati di flusso extracellulare acquisiti dagli sferoidi cellulari. (A) I dati OCR grezzi sono stati ottenuti da MCF-7 coltivato in 3 giorni e tracciato utilizzando il modello di Pearson per ottenere un coefficiente di correlazione tra densità di semina sferoide e OCR; Valore P impostato su 0,05. (B) I dati OCR grezzi sono stati normalizzati rispetto alla densità iniziale di semina sferoidale; (C) volume sferoide MCF-7 ottenuto dalla planimetria al microscopio; e (D) contenuto di DNA ds nucleare confrontato utilizzando il coefficiente di correlazione di Pearson. Abbreviazioni: OCR = tasso di consumo di ossigeno; ds DNA = DNA a doppio filamento. Fare clic qui per visualizzare una versione più grande di questa figura.
| Linea cellulare | Densità di semina (bene) | Crescita sferoide (giorni) | Volume sferoide finale (μM3) | OCR basale (pmolO2/min/pozzetto) | Sensibilità per OCR basale soddisfatta (SÌ/NO) |
| SKOV · | 1000 | 5 | 9,52E+06 | 28 ± 3,5 | Sì |
| SKOV · | 2000 | 5 | 2,38E+07 | 26 ± 1,4 | Sì |
| SKOV · | 4000 | 5 | 4,92E+07 | 36 ± 3.1 | Sì |
| SKOV · | 8000 | 5 | 1,11E+08 | 50 ± 7,9 | Sì |
| HepG2 · | 1000 | 5 | 1,11E+07 | 15 ± 0,7 | No |
| HepG2 · | 2000 | 5 | 2,88E+07 | 23 ± 1,8 | Sì |
| HepG2 · | 4000 | 5 | 5,46E+07 | 31 ± 1,7 | Sì |
| HepG2 · | 8000 | 5 | 1,21E+08 | 52 ± 2,8 | Sì |
| A549 · | 1000 | 5 | 2,11E+07 | 30 ± 2.5 | Sì |
| A549 · | 2000 | 5 | 3,57E+07 | 41 ± 1,6 | Sì |
| A549 · | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Sì |
| A549 · | 8000 | 5 | 1,44E+08 | 65 ± 8,4 | Sì |
| MCF-7 · | 1000 | 3 | 1,60E+07 | 29 ± 0,8 | Sì |
| MCF-7 · | 2000 | 3 | 2,52E+07 | 37 ± 1,7 | Sì |
| MCF-7 · | 4000 | 3 | 6,00E+07 | 46 ± 1,7 | Sì |
| MCF-7 · | 8000 | 3 | 1,06E+08 | 66 ± 2,9 | Sì |
Tabella 4: Parametri ottimizzati per la determinazione delle misure OCR basali in singoli sferoidi 3D. Abbreviazione: OCR = tasso di consumo di ossigeno.
File supplementare 1: Analisi delle dimensioni e del volume degli sferoidi. Fare clic qui per scaricare questo file.
File supplementare 2: Quantificazione del DNA a doppio filamento dagli sferoidi nella microplacca sferoide. Fare clic qui per scaricare questo file.
File supplementare 3: Raccomandazioni per il numero di repliche necessarie per ottenere set di dati di analisi XF affidabili. Fare clic qui per scaricare questo file.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Principali risultati e realizzazioni
Questo documento fornisce un protocollo dettagliato per sondare il metabolismo energetico mitocondriale di singoli sferoidi 3D utilizzando una serie di linee cellulari derivate dal cancro con l'analizzatore XF XFe96. Viene sviluppato e descritto un metodo per la rapida coltivazione di sferoidi cellulari A549, HepG2/C3A, MCF7 e SK-OV-3 utilizzando tecnologie cellorepellenti per l'aggregazione forzata. Questo protocollo affronta molte considerazioni sulla sonda del metabolismo sferoide con la tecnologia XF, tra cui (1) l'ottimizzazione dei protocolli di coltura sferoide e la manipolazione e il trasferimento di sferoidi in specifiche micropiastre per saggi sferoidi dal produttore della tecnologia dai loro vasi di coltivazione originali; (2) la concentrazione di composti respiratori da utilizzare e la dipendenza dal tempo della penetrazione di composti; (3) strategie di iniezione da utilizzare; e (4) modi per normalizzare i dati tra gruppi sperimentali. Tutte queste considerazioni sono state esaminate nel presente documento e sono discusse in modo più dettagliato di seguito. Questi metodi sono presentati come approcci semplificati per generare dati coerenti sul flusso metabolico di ossigeno utilizzando singoli sferoidi 3D con l'analizzatore di flusso XFe96. Questo approccio sperimentale può essere utilizzato come punto di partenza e rubrica per l'uso in altri modelli sferoidi che sono facilmente implementabili all'interno di un ambiente di laboratorio di base.
Considerazioni
Crescita, dimensioni e sensibilità degli sferoidi della tecnologia XF
Per stabilire dati riproducibili con la tecnologia XF, è essenziale caratterizzare e ottimizzare il test per il modello specifico. Questo approccio è relativamente semplice in un monostrato di base di cellule; tuttavia, questo presenta ulteriori sfide quando si coltivano cellule come sferoidi 3D. Durante gli esperimenti qui presentati, il supporto RPMI del produttore è stato integrato al momento dell'uso. Mentre si nota che alcune linee cellulari, vale a dire HepG2 / C3A, sono state coltivate nel mezzo di crescita DMEM, durante questi saggi relativamente brevi (~ 3-5 h), la sostituzione con formulazioni RPMI-DMEM ha avuto un impatto limitato sull'analisi XF. La formulazione dei due mezzi è molto simile e gli utenti potrebbero "sintonizzare" i media RPMI di Seahorse per abbinare la matrice dei loro terreni di coltura cellulare attraverso l'integrazione, ad esempio, aumento del glucosio, ulteriore aggiunta di fonti di carboidrati. Fondamentale per la formulazione finale di tutti i tamponi e mezzi XF è l'assenza di rosso fenolo, che rischia di interferire con le sonde fluorescenti all'interno della piastra della cartuccia della sonda XF, e bicarbonato di sodio, che porterà all'alcalinità a causa della mancanza di buffering di CO 2 presente negli incubatori dicolture cellulari. Altri supporti e buffer possono essere acquistati e/o realizzati internamente. Ad esempio, krebs Ringer HEPES buffer è un semplice buffer che può essere utilizzato per valutare la respirazione in molte cellule diverse, compresi i modelli sferoidi. Tuttavia, gli utenti di saggi XF dovrebbero notare che un cambiamento nel mezzo / buffer e la sua integrazione possono modificare la sua capacità di buffering complessiva. Ciò è particolarmente preoccupante quando gli utenti possono essere interessati a misurare l'ECAR, in cui il fattore tampone del mezzo deve essere valutato per consentire la trasformazione dell'ECAR in tasso di efflusso di protoni (PER).
Poiché l'OCR cellulare misurato dalla tecnologia XF è proporzionale alla densità cellulare quando il numero di cellule nel pozzo rientra nella sensibilità del sistema, era importante studiare questa relazione utilizzando singoli sferoidi 3D. Sondando l'OCR di singoli sferoidi 3D coltivati da 4 diverse linee cellulari tumorali seminate a densità di 1.000, 2.000, 4.000 o 8.000 cellule per pozzetto, dimostriamo che l'analizzatore XFe96 è abbastanza sensibile da rilevare i cambiamenti nella velocità di respirazione mitocondriale tra sferoidi 3D cresciuti da diverse densità di semina cellulare (Figura 3). Mostriamo che l'intervallo ottimale della densità di semina cellulare, e quindi il volume sferoide per la formazione di sferoidi 3D per sondare l'OCR, differiscono a seconda del tipo di cellula. Ciò è dimostrato dalla relazione lineare tra OCR e densità di seeding o volume sferoide (Figura 3). Per le cellule A549 e HepG2/C3A, la densità di semina ottimale per la sensibilità OCR era compresa tra 1.000 e 8.000 cellule/pozzetto; era di 2.000-8.000 celle / pozzo per MCF-7 e 4.000-8.000 celle / pozzo per celle SK-OV-3. Questi dati dimostrano che l'ottimizzazione delle dimensioni degli sferoidi è di particolare importanza quando si valuta l'OCR utilizzando la tecnologia XF.
Considerazioni sui volumi sferoidi minimi e massimi e sull'OCR basale
In generale, ci saranno sempre soglie minime e massime per i parametri OCR misurabili raccomandati dal produttore per questi esperimenti. Per l'analizzatore XFe96, l'OCR basale tra 20 pmol O2/min/well e 200 pmol O2/min/well sono rispettivamente i limiti inferiore e superiore. Questo è il caso delle cellule monostrato e degli sferoidi, e dove il modello sperimentale si trova all'interno di questo intervallo OCR dinamico dipenderà dalla quantità di materiale biologico disponibile, ad esempio, il numero di cellule come monostrato o la dimensione degli sferoidi. Vedere la Tabella 4 per un esempio di come le soglie OCR sono state raggiunte dai modelli sferoidi utilizzati qui. Può essere prudente controllare il livello di ossigeno all'interno del pozzo per il quale questi dati sono disponibili anche da queste misurazioni come dati di livello. Questo dovrebbe essere visto di routine da ogni esperimento per scopi di controllo della qualità. Se c'è esaurimento di ossigeno nel pozzo, questo sarà reso evidente all'interno dei dati. In tal caso, potrebbe essere necessario regolare i cicli di misurazione all'interno dell'esperimento; ad esempio, aumentando la fase di miscelazione in modo tale che il livello di ossigeno nel pozzo venga recuperato prima del successivo periodo di misurazione all'interno del ciclo di misurazione. Sebbene possibile, abbiamo scoperto che questo è molto improbabile per gli esperimenti a singolo sferoide che utilizzano le linee cellulari descritte.
Scelta di disacperatori mitocondriali per saggi di flusso extracellulare
Gli ionofori protonici, come il cianuro di carbonile 4-(trifluoromeossi) fenilidrazone (FCCP)21, il cianuro di carbonile m-clorofenil hydrazone (CCCP)22 o BAM1523, sono potenti sostanze chimiche a piccole molecole in grado di interrompere il gradiente elettrochimico protonico attraverso le membrane mitocondriali, inibendo la produzione di ATP e, infine, disaccoppiando la respirazione mitocondriale24 . Nuove piccole molecole continuano ad essere sviluppate per questi scopi, in particolare nel trattamento della malattia metabolica 25,26,27; fare riferimento a due ottime recensioni28,29. Al contrario, il disaccoppiamento della respirazione ossidativa è stato collegato a tossicità indesiderata fuori bersaglio30. Tuttavia, all'interno di saggi cellulari in vitro, la molecola FCCP depolarizza il potenziale di membrana mitocondriale ed esercita effetti off-target come la depolarizzazione della membrana plasmatica, interrompendo il flusso di ioni NA+ 31; interferenza con l'elaborazione delle proteine cellulari32 e persino inducendo la senescenza cellulare33. BAM15 è stato originariamente introdotto nel 2013 come uncoupler mitocondriale con un'influenza minima sulle membrane plasmatiche23, con attività protonoforica nella gamma micromolare in cellule intere e gamma nanomolare nei mitocondri isolati23,34.
Data la potenza della FCCP sulla depolarizzazione della membrana plasmatica, BAM15 è un protonoforo più affidabile per disaccoppiare la respirazione in cellule intere intatte in saggi di flusso extracellulare. Sebbene FCCP e la sua controparte, CCCP, siano stati utilizzati per oltre 50 anni per analizzare le capacità respiratorie massime e continuino ad essere ampiamente utilizzati negli studi XF, l'uso di queste piccole molecole spesso sottovaluta la capacità metabolica mitocondriale e cellulare. Ciò è in parte legato al motivo per cui così tante pubblicazioni che utilizzano la tecnologia XF cadono nella trappola di riportare capacità respiratorie di riserva negative o sottovalutano le vere capacità respiratorie mitocondriali quando viene utilizzato FCCP. La potenza aggiunta di FCCP in cellule e tessuti intatti spesso porta a compromettere la funzione mitocondriale e le cellule possono avere difficoltà a operare in modo appropriato per sostenere la massima capacità respiratoria attraverso più cicli di misurazione dopo la loro aggiunta, anche a concentrazioni molto basse35. Pertanto, la risposta delle cellule alla FCCP può essere trovata in molti studi per diminuire dopo il periodo del ciclo di misurazione iniziale. Mentre FCCP è stato utilizzato di routine per l'analisi XF, BAM15 è usato preferenzialmente nei casi che coinvolgono cellule intere o modelli sferoidi, dato che può mantenere una capacità respiratoria massima nei mitocondri completamente depolarizzati a concentrazioni fino a 10 μM 3. Inoltre, BAM15 induce effetti sull'acidificazione extracellulare, che coincide con quella dell'ossidazione dei nutrienti attraverso l'idratazione di CO2 per formare HCO3- e H+ in misura maggiore rispetto a FCCP3. Tuttavia, nel caso di mitocondri isolati e cellule permeabilizzate, uno qualsiasi di questi disaccoppiatori dovrebbe funzionare così come BAM15 per il disaccoppiamento mitocondriale se titolato alla concentrazione corretta.
Cinetica della penetrazione del composto e del ciclo di analisi
Le concentrazioni, la penetrazione e i profili cinetici dei composti chimici utilizzati per condurre un tipico test MitoStress con sferoidi 3D utilizzando l'analizzatore XF sono più complessi da affrontare. Dato che gli sferoidi presentano una struttura 3D, la penetrazione di molecole attraverso il diametro dello sferoide è un processo infinitamente più complesso rispetto ai monostrati cellulari. Ad esempio, la penetrazione cinetica e, quindi, la sensibilità al sorafenib chemioterapico è stata determinata dall'età sferoide e, quindi, dalle dimensioni in un modello sferoide HepG236. La capacità delle sostanze chimiche a piccole molecole (ad esempio, farmaci, nanoparticelle) di raggiungere un bersaglio biologico dipende da diversi fattori sottostanti, tra cui la complessità del sistema di essere penetrato dinamicamente e diffuso attraverso37,38. Ciò è particolarmente vero per i farmaci che prendono di mira il tessuto tumorale39. Simile al targeting tumorale nel contesto di uno sferoide 3D, dimensioni, compattezza e altre risposte fenotipiche come l'espressione di proteine trasportatrici di farmaci possono governare il tempo di penetrazione e la concentrazione di un composto necessario per suscitare una risposta biologica.
In questo protocollo, abbiamo affrontato il problema del tempo di penetrazione e della concentrazione di piccole molecole in risposta all'oligomicina inibitore dell'ATP sintasi, al protonoforo e allo sganciatore mitocondriale BAM15 e alla combinazione degli inibitori del Complesso I e del Complesso III rotenone e antimicina A. Sondando l'OCR di singoli sferoidi MCF-7 esposti a titolazioni multiple di questi comuni composti respiratori, dimostriamo che la concentrazione ottimale di ciascun composto necessaria per indurre una frequenza respiratoria allo stato stazionario rientra in un intervallo simile a quello delle cellule monostrato (Figura 4). È importante sottolineare che, e a differenza delle loro controparti monostrato, è dimostrato che aumentare il numero di cicli di misurazione tra le iniezioni è la chiave per ottenere un OCR a stato stazionario in singoli sferoidi 3D. Questi dati evidenziano l'importanza della penetrazione dei composti e dei rispettivi profili cinetici quando si esplorano i parametri respiratori mitocondriali degli sferoidi 3D utilizzando questi approcci. Utilizzando le proprietà di ottimizzazione degli sferoidi, le concentrazioni di composti e i tempi del ciclo di misurazione informati dai dati presentati in Figura 3, Figura 4 e Figura 5, è stato stabilito un test MitoStress convalidato per sondare parametri specifici del metabolismo ossidativo mitocondriale in una gamma di sferoidi 3D derivati dal cancro (Figura 6). Di importanza, e come alcune linee cellulari tumorali monostrato40, la capacità respiratoria massima (tasso di respirazione non disaccoppiata-stimolata) di alcuni sferoidi 3D derivati dal cancro è stata inibita dall'oligomicina (Figura 5). In particolare, gli sferoidi 3D cresciuti da cellule A549 o MCF-7 hanno mostrato un tasso massimo significativamente più basso di respirazione quando disaccoppiati con BAM15 dopo l'iniezione di oligomicina rispetto all'essere disaccoppiati da BAM15 senza oligomicina (Figura 5F e Figura 5H). Dato che questo effetto può essere presente all'interno di altre colture sferoidi 3D, suggeriamo che, a meno che non venga utilizzato un protocollo precedentemente convalidato, la capacità respiratoria massima negli sferoidi 3D dovrebbe essere stimata senza oligomicina.
Raccolta simultanea di dati ECAR come misura del flusso glicolitico negli sferoidi cellulari
Come tipicamente visto in letteratura o nelle informazioni del produttore della tecnologia, il tasso glicolitico degli sferoidi, misurato come ECAR, è un parametro secondario che può essere catturato insieme all'OCR. Il calcolo dell'ECAR da solo non è un parametro utile o significativo in nessun esperimento XF in quanto non viene corretto per la capacità tampone del tampone del saggio XF o l'aggiunta di acidificazione mitocondriale, che deriva dall'idratazione di CO2 a HCO3- e H +. ECAR è perspicace solo una volta applicate queste correzioni dei dati, dopo di che diventa possibile fornire conclusioni più accurate sul flusso glicolitico. Per correggere la capacità di buffering per generare dati PER più significativi, è necessario conoscere il volume della microcamera per la micropiastra sferoide. Il produttore non è stato in grado di fornire un volume reale per questo con la micropiastra sferoidale e, pertanto, i dati PER non possono essere determinati facilmente. In effetti, sebbene queste misurazioni potessero essere raggiunte empiricamente, questo andava oltre lo scopo di questo manoscritto. Tuttavia, con le correzioni appropriate e conoscendo il volume della microcamera per una data dimensione sferoidale presente (ad esempio, ottenendo una misura della densità sferoide) nel pozzo, i dati ECAR diventerebbero significativi e potrebbero essere effettuati calcoli di PER glicolitico. Quindi, i dati XF potrebbero quindi essere più informativi per studiare il metabolismo glicolitico e ossidativo negli sferoidi, ma solo se questi parametri fossero considerati in profondità.
Formazione, manipolazione, trasferimento e movimento sferoidi
Alcune linee cellulari sono più adatte alla formazione di sferoidi rispetto ad altre e potrebbero non formare affatto sferoidi, ad esempio, le cellule tumorali ovariche MCF-741,42 formano sferoidi altamente circolari rispetto ad altre linee cellulari (Figura 3). Come altro esempio, le cellule tumorali pancreatiche Capan-1 hanno dimostrato di formare sferoidi migliori rispetto a Panc-1 o BxPC343. Allo stesso modo, le linee cellulari di carcinoma epatico sono note per avere capacità variabili di formare sferoidi compatti 5,44, con un cambiamento osservato nel fenotipo come un aumento del metabolismo dei farmaci o la produzione di albumina, come nel caso di HepG2 contro HepG2 / C3A 9,45,46 o sferoidi HepaRG 17,47,48 . Pertanto, gli utenti dovrebbero ottimizzare le tecniche di coltivazione degli sferoidi di conseguenza ed eseguire esperimenti di titolazione per determinare la densità di semina ottimale e il corso del tempo di coltivazione. Inoltre, la formulazione e la composizione dei mezzi di saggio hanno dimostrato di avere un impatto sulla formulazione sferoide, compresa l'aggiunta di metilcellulosa, spesso aggiunta ai mezzi per aumentare la viscosità della matrice 43,49,50. Quindi, la composizione ottimale del mezzo cellulare dovrebbe essere determinata empiricamente per tutte le linee cellulari utilizzate.
Il numero di scambi di mezzi durante la coltura sferoidale è determinato dalla linea cellulare utilizzata. Tuttavia, in genere, gli scambi medi a mezzo volume ogni 2-3 giorni sono applicabili nella maggior parte dei casi per reintegrare i nutrienti. Abbiamo utilizzato l'approccio dell'aggregazione forzata per generare sferoidi 3D utilizzando micropiastre cellulore-repellenti da fonti disponibili in commercio per un rapido sviluppo e distribuzione di modelli sferoidi negli studi di analisi XF. Tuttavia, piattaforme alternative potrebbero essere più adatte a generare sferoidi da altri tipi di cellule, ad esempio approcci a goccia sospesa o incorporati in matrice. Nei laboratori con risorse limitate, gli utenti potrebbero voler guardare verso la tecnica di sovrapposizione di agarosio-liquido per la formazione di superfici di micropiastre repellenti per cellule51,52 per ridurre significativamente i costi economici delle fasi iniziali di sviluppo del metodo sferoide. Il movimento degli sferoidi tra i vasi di coltura è necessario per eseguire l'analisi XF e altri saggi a valle. La facilità di trasferimento è in genere dettata dalla dimensione sferoide e dalla densità complessiva. Si consiglia di utilizzare una punta della pipetta a orifizio largo P200 o P1000 per mantenere l'integrità dello sferoide; le punte delle pipette di foro più piccolo rischiano l'interruzione meccanica dello sferoide, che può essere acquistato commercialmente o, con cura, realizzato semplicemente tagliando l'estremità della punta della pipetta per aumentare l'orifizio. Tuttavia, questo approccio può essere suscettibile di introdurre un pelo nella plastica intorno all'estremità della punta, che potrebbe causare interruzioni meccaniche durante la manipolazione. L'uso di una retroilluminazione o di una lightbox è utile anche per la manipolazione e l'osservazione degli sferoidi al microscopio di dissezione come passo essenziale per garantire il successo del trasferimento degli sferoidi nella micropiastra del saggio sferoidale. Inoltre, la posizione sferoide all'interno del pozzetto di una micropiastra di saggio sferoidale è di particolare importanza e influisce direttamente sull'OCR e sull'effetto composto durante un tipico test MitoStress (Figura 7), molto probabilmente a causa della relazione tra la posizione sferoide e i fluorofori della sonda del sensore.
Correzione dello sfondo e pozzetti di controllo della temperatura
L'uso di saggi basati su micropiastre è un approccio ampiamente utilizzato in diverse aree di ricerca; tuttavia, il loro uso presenta diverse sfide pratiche. Come è vero in altri approcci sperimentali, in particolare quelli che utilizzano il formato array 96 (o superiore), la geometria e il posizionamento delle micropiastre possono influenzare la temperatura e i gradienti di scambio di gas attraverso la piastra nel tempo, spesso indicati come "effetti bordo"53,54. Abbiamo scoperto che lo stesso vale per la micropiastra di analisi sferoidale. Secondo le linee guida e i protocolli del produttore, i pozzetti angolari più esterni: A1, A12, H1 e H12 sono sempre designati come pozzi di correzione di fondo e controllo della temperatura per l'analizzatore XFe96. Al contrario, con il formato array a 24 pozzetti, A1 e D6 sono designati come pozzi di controllo, insieme ad altri due pozzetti distribuiti uniformemente al centro della piastra a B4 e C3. Eseguendo l'analisi sferoidale XF, abbiamo riscontrato una deviazione significativa nei dati inizialmente raccolti utilizzando le indicazioni del produttore. Ciò nonostante l'inclusione delle misure necessarie per garantire la preequilibrazione del test alla temperatura e al contenuto di CO2 prima di iniziare l'acquisizione dei dati, spesso producendo valori negativi per l'OCR dopo l'iniezione di alcuni inibitori respiratori (Figura 8).
Abbiamo scoperto che queste osservazioni sono probabilmente dovute agli effetti dei bordi attraverso la micropiastra del saggio sferoidale. Nella Figura 8, abbiamo scoperto che ridistribuendo i pozzetti di controllo dello sfondo attraverso la micropiastra, i dati XF sono stati regolati di circa 2 volte. Due ragioni più probabili sono (1) dovute agli effetti di evaporazione nei pozzi di bordo che si traducono in una camera di volume totale più piccola per la sonda XFe96 da cui campionare e (2) da equilibri di temperatura inadeguati tra quei pozzetti designati per la correzione di fondo e pozzi campione, con conseguente set di dati che mascherano o gonfiano eccessivamente l'OCR. Per evitare tali risultati, si raccomanda pertanto, soprattutto nel contesto dell'analisi sferoide, che gli utenti ridistribuiscano i pozzetti designati per la correzione di fondo su tutta la micropiastra del saggio sferoide e adottino le misure necessarie per pre-equilibrare il loro test prima di acquisire i dati XF.
Normalizzazione dei dati
Oltre a fornire un protocollo dettagliato per sondare il metabolismo energetico mitocondriale di singoli sferoidi 3D con tecnologia XF, questo documento presenta anche possibili modi per normalizzare i dati della frequenza respiratoria mitocondriale ottenuti con sferoidi 3D. Utilizzando i dati sulla frequenza respiratoria ottenuti con sferoidi MCF-7 coltivati a diverse densità di semina cellulare (Figura 3), presentiamo frequenze respiratorie mitocondriali basali da sferoidi MCF-7 di dimensioni e diametro crescenti quando normalizzati alla densità iniziale di semina cellulare, volume sferoide e contenuto di dsDNA (Figura 9). Il metodo di normalizzazione appropriato è fondamentale per l'interpretazione accurata dei set di dati XF, in particolare quando si confrontano modelli sferoidi 3D in vitro e diversi tipi di cellule. Una scarsa normalizzazione può portare a risultati errati che semplicemente non possono essere confrontati tra i set di dati. Il contenuto proteico non è preferito per la normalizzazione dei dati XF sferoidi, poiché i pretrattamenti possono influire sui tassi di sintesi proteica senza effetti significativi sulla frequenza respiratoria. Inoltre, quantità significative e incoerenti di proteine possono legarsi a micropiastre sferoidi dopo la lisi cellulare, introducendo variazioni nel contenuto proteico tra i pozzetti. Ciò può essere ulteriormente complicato nelle analisi XF che utilizzano sferoidi o cellule non aderenti che richiedono colle biomolecolari per legarsi, che possono contenere proteine.
Contrariamente al contenuto proteico intracellulare, il contenuto di DNA nucleare è indipendente dal tipo di cellula ed è proporzionale al numero di cellule (Figura 9D), un approccio più accurato e meno dispendioso in termini di tempo rispetto alla disaggregazione degli sferoidi per la quantificazione del numero cellulare. Al contrario, Yepéz et al.55, conducendo analisi XF in monostrati di cellule di fibroblasti, hanno scoperto che la normalizzazione dei dati XF al numero di cellule ha introdotto una maggiore dispersione dei dati rispetto a prima della normalizzazione. Il contenuto di DNA nucleare è indipendente dallo stato differenziato o dal fenotipo e quindi è più accurato per la normalizzazione dei dati sferoidi nei saggi XF rispetto al contenuto proteico. Il contenuto di DNA è stato anche una strategia collaudata per l'analisi di altri set di dati legati al metabolismo56. Tuttavia, è importante notare che il contenuto di DNA nucleare è quantificato da tutte le cellule presenti all'interno dello sferoide; pertanto, la normalizzazione al contenuto di DNA non è raccomandata per i set di dati XF in cui gli sferoidi subiscono trattamenti che possono comportare una significativa perdita di vitalità cellulare. Per tali set di dati, se possibile, è preferibile la normalizzazione alla vitalità cellulare, oppure i dati possono essere corretti al basale per la respirazione basale.
Utilizzare la capacità respiratoria di riserva come esempio per l'importanza della normalizzazione dei dati
La capacità respiratoria di riserva è una misura del tasso di capacità respiratoria mitocondriale massima meno la frequenza respiratoria mitocondriale basale (Figura 6). Tuttavia, il problema con la segnalazione di dati di questo tipo come tasso, cioè pmolO2 / min / bene all'interno di determinati esperimenti, è che i dati sono privi di normalizzazione. Anche se si normalizzano i dati sferoidi in densità cellulare / contenuto di DNA, questo spesso esclude il parametro chiave che deve essere normalizzato per la densità mitocondriale all'interno delle cellule. Dato che un cambiamento nella densità mitocondriale porterà a un cambiamento proporzionale nella respirazione basale e massimale, aumenterà anche la capacità inutilizzata. Ad esempio, se l'OCRbasale sferoide è 200 e l'OCRmax è 400, la capacità inutilizzata viene riportata come 200; se l'OCRbasale è 100 e l'OCRmax, anche la capacità di riserva è 100; tuttavia, in percentuale, sono entrambi il 50% del massimo (o il 100% del basale). Pertanto, la capacità inutilizzata non viene modificata tra questi due esempi, nonostante le differenze nei tassi di 200 e 100 quando calcolati come pmols O2 / min / pozzetto. I valori normalizzati internamente sono più affidabili e approfonditi per rendere i dati XF più comparabili tra studi e progetti. Per fare questo per la capacità respiratoria di riserva, abbiamo scelto di presentarlo come una percentuale di respirazione massima invece di un tasso assoluto. Questo potrebbe anche essere presentato come una percentuale della respirazione basale. Questo sarebbe il caso se si lavora con cellule o sferoidi. Tuttavia, dato che la posizione dello sferoide nella piastra del microwell può alterare l'OCR assoluto ma non i cambiamenti relativi con inibitori o disaccoppiatori, è più importante guardare alle risposte normalizzate internamente negli sferoidi come variazione di piega o percentuali.
I modelli sferoidi qui generati presentano una gamma di tipi di cellule e architetture che non possono essere catturati nei modelli 2D classici. Questi includono una disposizione eterogenea e spaziale delle cellule in tre dimensioni, contatti cellula-cellula migliorati (ad esempio, formazione di giunzioni gap e matrici extracellulari) e gradienti biochimici attraverso il diametro sferoide (ad esempio, gradienti di pH, accesso alla diffusione di ossigeno ai nutrienti). L'uso del flusso extracellulare per studiare la biologia sferoide in vitro potrebbe consentire di identificare bersagli ottimali per le terapie farmacologiche attraverso osservazioni di perturbazione metabolica. Questi potrebbero essere estrapolati da sferoidi in vitro a tumori in vivo e identificare percorsi che possono indirizzare il metabolismo del tumore sferoide, ad esempio l'utilizzo di carboidrati durante la crescita sferoide. Le modalità terapeutiche possono essere efficaci nel colpire gli sferoidi nelle prime fasi di crescita, ma si dimostrano meno efficaci nelle fasi successive della crescita sferoide man mano che matura la complessità della rete metabolica. Per concludere, lo sviluppo di modelli di coltura cellulare 3D e sofisticate tecnologie di analisi nella ricerca biologica continuerà ad essere un campo dinamico e in rapida evoluzione con un potenziale insuperabile. L'analisi del flusso extracellulare di sferoidi in coltura cellulare in vitro potrebbe essere impiegata come metodo di ricerca all'avanguardia per far progredire i risultati della ricerca che potrebbero essere estrapolati per comprendere meglio la biologia rilevante per l'uomo, ridurre l'uso di modelli animali nella ricerca e migliorare la ricerca incentrata sul paziente.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Gli autori non hanno conflitti di interesse da dichiarare.
Acknowledgments
N.J.C è stato supportato da un BBSRC MIBTP CASE Award con Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Ricerca sul cancro Numero 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
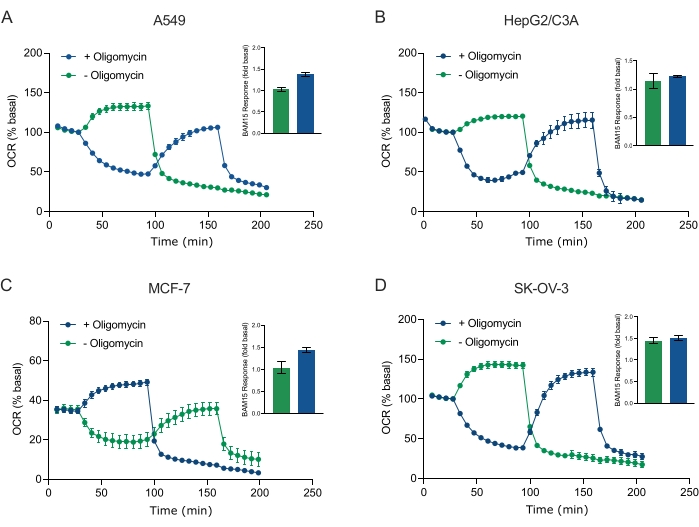
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
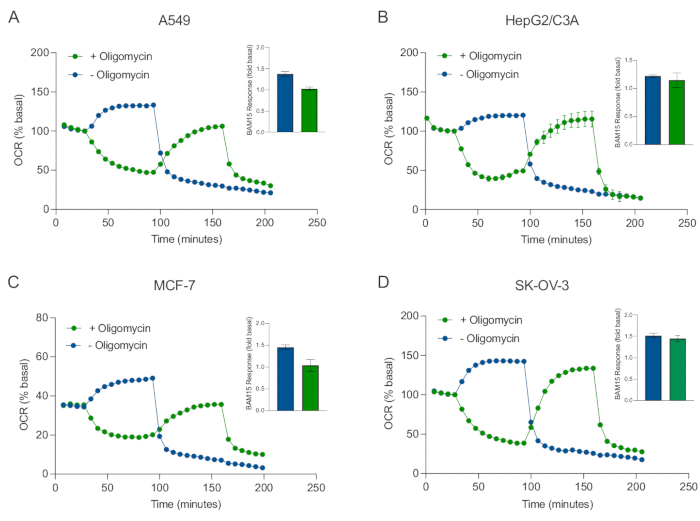
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
