

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Estos protocolos ayudarán a los usuarios a sondear el metabolismo energético mitocondrial en esferoides derivados de líneas celulares de cáncer 3D utilizando el análisis de flujo extracelular Seahorse.
Abstract
Los agregados celulares tridimensionales (3D), denominados esferoides, se han convertido en la vanguardia del cultivo celular in vitro en los últimos años. En contraste con el cultivo de células como monocapas bidimensionales de una sola célula (cultivo 2D), el cultivo celular esferoide promueve, regula y apoya la arquitectura celular fisiológica y las características que existen in vivo, incluida la expresión de proteínas de matriz extracelular, señalización celular, expresión génica, producción de proteínas, diferenciación y proliferación. La importancia del cultivo 3D ha sido reconocida en muchos campos de investigación, incluyendo oncología, diabetes, biología de células madre e ingeniería de tejidos. Durante la última década, se han desarrollado métodos mejorados para producir esferoides y evaluar su función metabólica y destino.
Los analizadores de flujo extracelular (XF) se han utilizado para explorar la función mitocondrial en microtejidos 3D, como los esferoides, utilizando una placa de captura de islotes XF24 o una microplaca esferoide XFe96. Sin embargo, no se han descrito en detalle distintos protocolos y la optimización del sondeo del metabolismo energético mitocondrial en esferoides utilizando la tecnología XF. Este artículo proporciona protocolos detallados para sondear el metabolismo energético mitocondrial en esferoides 3D individuales utilizando microplacas esferoides con el analizador XFe96 XF. Utilizando diferentes líneas celulares de cáncer, se ha demostrado que la tecnología XF es capaz de distinguir entre la respiración celular en esferoides 3D no solo de diferentes tamaños, sino también de diferentes volúmenes, números de células, contenido y tipo de ADN.
Las concentraciones óptimas del compuesto efector mitocondrial de oligomicina, BAM15, rotenona y antimicina A se utilizan para sondear parámetros específicos del metabolismo energético mitocondrial en esferoides 3D. Este documento también discute los métodos para normalizar los datos obtenidos de los esferoides y aborda muchas consideraciones que deben considerarse al explorar el metabolismo de los esferoides utilizando la tecnología XF. Este protocolo ayudará a impulsar la investigación en modelos avanzados de esferoides in vitro .
Introduction
Los avances en los modelos in vitro en la investigación biológica han progresado rápidamente en los últimos 20 años. Tales modelos ahora incluyen modalidades de órgano en un chip, organoides y esferoides de microtejido 3D, todos los cuales se han convertido en un enfoque común para mejorar la traducción entre estudios in vitro e in vivo. El uso de modelos in vitro avanzados, particularmente esferoides, abarca varios campos de investigación, incluida la ingeniería de tejidos, la investigación con células madre, el cáncer y la biología de enfermedades 1,2,3,4,5,6,7 y las pruebas de seguridad, incluida la toxicología genética 8,9,10, la toxicología de nanomateriales 11, 12,13,14, y pruebas de seguridad y eficacia de medicamentos 8,15,16,17,18,19.
La morfología celular normal es crítica para el fenotipo y la actividad biológica. El cultivo de células en esferoides microtisulares 3D permite a las células adoptar una morfología, función fenotípica y arquitectura, más parecida a la observada in vivo pero difícil de capturar con las técnicas clásicas de cultivo celular monocapa. Tanto in vivo como in vitro, la función celular se ve directamente afectada por el microambiente celular, que no se limita a la comunicación y programación celular (por ejemplo, formaciones de unión célula-célula, oportunidades para formar nichos celulares); exposición celular a hormonas y factores de crecimiento en los entornos inmediatos (por ejemplo, exposición a citoquinas celulares como parte de una respuesta inflamatoria); composición de matrices físicas y químicas (por ejemplo, si las células se cultivan en plástico de cultivo de tejidos rígidos o en un entorno de tejido elástico); y lo más importante, cómo el metabolismo celular se ve afectado por la nutrición y el acceso al oxígeno, así como por el procesamiento de productos de desecho metabólicos como el ácido láctico.
El análisis de flujo metabólico es una forma poderosa de examinar el metabolismo celular dentro de sistemas in vitro definidos. Específicamente, la tecnología XF permite el análisis de cambios en vivo y en tiempo real en la bioenergética celular de células y tejidos intactos. Dado que muchos eventos metabólicos intracelulares ocurren del orden de segundos a minutos, los enfoques funcionales en tiempo real son primordiales para comprender los cambios en tiempo real en el flujo metabólico celular en células y tejidos intactos in vitro.
Este artículo proporciona protocolos para cultivar líneas celulares derivadas del cáncer A549 (adenocarcinoma de pulmón), HepG2/C3A (carcinoma hepatocelular), MCF-7 (adenocarcinoma de mama) y SK-OV-3 (adenocarcinoma de ovario) como modelos de esferoides 3D in vitro utilizando enfoques de agregación forzada (Figura 1). También (i) describe en detalle cómo sondear el metabolismo energético mitocondrial de esferoides 3D individuales utilizando el analizador XF Agilent XFe96, (ii) destaca formas de optimizar los ensayos XF utilizando esferoides 3D individuales, y (iii) discute consideraciones y limitaciones importantes de sondear el metabolismo de los esferoides 3D utilizando este enfoque. Lo más importante es que este artículo describe cómo se recopilan los conjuntos de datos que permiten el cálculo de la tasa de consumo de oxígeno (OCR) para determinar la fosforilación oxidativa y, por lo tanto, la función mitocondrial en los esferoides celulares. Aunque no se analiza para este protocolo, la tasa de acidificación extracelular (ECAR) es otro parámetro que se mide junto con los datos de OCR en experimentos XF. Sin embargo, ECAR a menudo se interpreta de manera deficiente o incorrecta a partir de conjuntos de datos XF. Proporcionamos un comentario sobre las limitaciones del cálculo de ECAR siguiendo los enfoques básicos del fabricante de la tecnología.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figura 1: Flujo de trabajo gráfico para la generación de esferoides celulares, análisis de flujo extracelular y ensayos posteriores. Cuatro líneas celulares de cáncer se cultivaron selectivamente como monocapas (A), se separaron de los matraces de cultivo de tejidos y se sembraron en microplacas de 96 pocillos de unión ultrabaja para formar esferoides (B). El carcinoma de pulmón A549, el carcinoma hepático HepG2/C3A, el adenocarcinoma de ovario SK-OV-3 y las células de carcinoma de mama MCF-7 se sembraron a 1 × 103-8 × 103 células/pozo y crecieron hasta 7 días para formar esferoides individuales y optimizar la densidad de siembra de esferoides y el tiempo de cultivo mediante observación continua y mediciones planimétricas. Una vez formados, los esferoides individuales se lavaron en un medio XF libre de suero y se sembraron cuidadosamente en microplacas de ensayo de esferoides, precubiertas con poli-D-lisina (C). Los esferoides se sometieron a análisis de flujo extracelular utilizando el analizador XFe96 utilizando varios protocolos para abordar: (1) el tamaño óptimo de los esferoides para la respuesta de la respiración mitocondrial basal; (2) titulación optimizada de inhibidores respiratorios mitocondriales; (3) optimización de la colocación de esferoides dentro de pozos de microplacas. (D) Se utilizaron análisis posteriores a la XF, microscopía de contraste de fase y cuantificación del ADN esferoide para la normalización de datos y otros ensayos in vitro posteriores. Haga clic aquí para ver una versión más grande de esta figura.
1. Cultivo de líneas celulares de cáncer como esferoides 3D in vitro
| Línea celular | Descripción | Medio de cultivo | Fuente |
| A549 | Línea celular de carcinoma de pulmón | RPMI 1640 | Colección Europea de Cultivos Celulares Autenticados (ECACC) |
| Piruvato de sodio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Línea celular de carcinoma hepático, un derivado clonal de la línea celular hepG2 madre | DMEM | Colección Americana de Cultivo de Tejidos (ATCC) |
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| MCF7 | Línea celular de adenocarcinoma mamario | RPMI 1640 | Colección Europea de Cultivos Celulares Autenticados (ECACC) |
| Piruvato de sodio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Línea celular de adenocarcinoma ovárico | RPMI 1640 | Colección Europea de Cultivos Celulares Autenticados (ECACC) |
| Piruvato de sodio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| Componente | Medio de ensayo RPMI (volumen final de 50 ml) | ||
| Base Media | Agilent Seahorse XF RPMI, pH 7.4 | ||
| Glucosa (1 M de stock estéril) | 11 mM (solución madre de 0,55 ml) | ||
| L-glutamina (stock estéril de 200 mM) | 2 mM (0,5 ml de solución madre) | ||
| Piruvato de sodio (stock estéril de 100 mM) | 1 mM (0,5 ml de solución madre) |
Tabla 1: Medios de línea celular de cáncer y composiciones de medios XF.
- Cultive todas las líneas celulares utilizando la técnica estándar de cultivo de tejido aséptico y confirme que están libres de micoplasma utilizando un kit de ensayo adecuado.
- Cultivo de las líneas celulares en matraces de cultivo tisular T75 o equivalentes, utilizando el medio recomendado (Tabla 1). Cultivar las líneas celulares al 65-80% de confluencia y pasarlas regularmente hasta un máximo de 25 pasajes.
- Enjuague los matraces de cultivo celular dos veces en solución salina tamponada con fosfato modificado (DBPS) de Dulbecco.
- Separar las células de los matraces con 3 ml del reactivo de disociación celular (ver la Tabla de Materiales) durante 5 min a 37 °C y confirmar el desprendimiento por microscopía.
- Aspire la suspensión celular desprendida suavemente para asegurar una suspensión unicelular y desactivar el reactivo de disociación celular con 7 ml de medio de cultivo tisular completo.
- Recoger las células por centrifugación a 300 × g durante 5 min, desechar el sobrenadante y volver a suspender las células en medio completo.
- Cuente las células utilizando un hemocitómetro o un contador celular automatizado y titule a la densidad celular deseada requerida para la siembra.
NOTA: Para sembrar una placa entera de 96 pocillos a 100 μL/pocillo a 4 × 103 células/pozo, las células deben ajustarse a 4 × 104 células/ml en un volumen recomendado de 12 ml. - Decantar la suspensión celular en un reservorio estéril y dispensar 100 μL de la suspensión celular en cada pocillo de una microplaca repelente celular utilizando un pipettor multicanal.
NOTA: Solo los 60 pocillos internos de una microplaca deben sembrarse y el resto debe llenarse con DPBS. Esto formará una barrera de evaporación, asegurará la homogeneidad de los esferoides en toda la placa y minimizará los efectos del borde de la placa. - Centrifugar microplacas esferoides a 300 × g durante 15 min para forzar a las células a formar agregados sueltos.
- Incubar las placas a 37 °C, 5% CO2 durante un mínimo de 3 días para asegurar la formación de esferoides.
- Realizar microscopía de contraste de fase utilizando prácticas de laboratorio estandarizadas para monitorear el crecimiento de los esferoides. Reponga el medio de cultivo celular cada 3 días o dos veces por semana realizando un intercambio de medio volumen.
2. Sondeo del metabolismo energético mitocondrial de esferoides individuales utilizando la tecnología de flujo extracelular (XF)
- Preparación del ensayo (un día antes)
- Compruebe la viabilidad de los esferoides utilizando un microscopio de luz invertida con contraste de fase a un aumento de 4x para garantizar la estructura esferoide intacta, la morfología y la uniformidad general entre las muestras.
- Hidrate el cartucho del sensor.
- Alícuota ~ 20 ml del calibrante en un tubo cónico.
- Coloque el tubo cónico que contiene el calibrante en una incubadora sin CO2 a 37 °C durante la noche.
- Retire el contenido del kit de ensayo.
- Retire el cartucho del sensor de la placa de utilidad y colóquelo boca abajo en la encimera junto a la placa de utilidad.
- Pipetear 200 μL de ddH2O estéril en cada pocillo de la placa de utilidad del cartucho del sensor utilizando una pipeta P300 multicanal.
- Coloque el cartucho del sensor en la parte superior de la placa de utilidad.
- Compruebe que el nivel de agua en cada pozo es lo suficientemente alto como para sumergir las sondas del sensor.
- Transfiera el cartucho del sensor ensamblado a una incubadora de 37 °C sin CO2 y déjelo durante la noche.
NOTA: Este paso se puede realizar 12-72 h antes del comienzo del ensayo.
- Microplaca de ensayo de esferoides de capa
- Utilizando técnicas asépticas, añadir 30 μL/pocillo de solución estéril de poli-D-lisina (0,1 mg/ml) a la microplaca esferoide e incubarla durante 30 min a temperatura ambiente.
- Aspire la solución de cada pocillo de la microplaca esferoide, invierta la placa y tóquela firmemente sobre papel de seda para eliminar cualquier solución residual.
- Lavar la placa dos veces con 200 μL/pocillo de ddH2O estéril.
- Después del lavado final, invierta la microplaca y cúbrala firmemente sobre papel de seda para eliminar el agua residual.
- Deje que la placa se seque al aire durante 30 minutos antes de usarla o almacenarla a 4 °C para su uso futuro.
NOTA: La microplaca de ensayo de esferoides debe recubrirse con un adhesivo molecular para garantizar que los esferoides se fijen en la parte inferior de la microplaca. Sin un adhesivo molecular, los esferoides pueden desprenderse e interferir con los resultados del ensayo. Otros adhesivos moleculares también se pueden utilizar como una alternativa a la poli-D-lisina para placas de prerecubrimiento. Las placas preestucados se pueden almacenar a 4 °C, pero deben dejarse equilibrar a temperatura ambiente antes del comienzo del ensayo.
- Preparar el medio de ensayo XF
- Preparar el medio XF RPMI, como se detalla en la Tabla 1, y el filtro estéril con un filtro de jeringa de 0,22 μm
- Preparación del ensayo (1 h antes del ensayo)
- Precalentar el medio de ensayo XF RPMI suplementado a 37 °C.
- Precalentar la microplaca de ensayo de esferoides recubiertos en una incubadora sin CO2 a 37 °C o en un baño seco.
- Prepare el cartucho del sensor.
- Saque el tubo cónico que contiene el calibrante y el cartucho del sensor de la incubadora de aire.
- Retire el cartucho del sensor de la placa de utilidad y colóquelo boca abajo en la superficie de trabajo.
- Usando una pipeta multicanal P300, aspire el agua de la placa de servicio y deséchela.
- Vierta la solución de calibrante en un depósito de reactivo estéril y agregue 200 μL/pocillo del calibrante precalentado a la placa de utilidad utilizando una pipeta multicanal P300.
- Tome el cartucho del sensor y colóquelo de nuevo en la parte superior de la placa de utilidad, asegurándose de que los sensores estén bien sumergidos en el calibrante.
- Vuelva a transferir el cartucho del sensor montado a la incubadora sin CO2 a 37 °C hasta que esté listo para cargar las soluciones de inyección de puerto.
- Lave los esferoides con el medio de ensayo.
- Retire la placa de cultivo de esferoides de la incubadora de 37 °C, 5% de CO2 y observe los esferoides bajo el microscopio para garantizar su integridad antes de los pasos de transferencia de esferoides.
- Cargue todos los pocillos de la placa esferoide con 180 μL/pocillo de medio de ensayo precalentado, incluidos los pozos de corrección de fondo.
- Llene parcialmente una placa de Petri de 7 cm con 3 ml del medio de ensayo.
- Usando una pipeta multicanal cargada con puntas de pipeta de orificio ancho, transfiera los esferoides de la placa de cultivo de 96 pocillos a placas de Petri de 7 cm ajustando el pipeteador a un volumen de aspiración de 10-50 μL.
- Semillas de esferoides en la microplaca de ensayo de esferoides pre-recubierta.
- Usando un microscopio de disección y un aparato de caja de luz, transfiera los esferoides de la placa de Petri a la microplaca de ensayo de esferoides como se detalla a continuación.
- Ajuste el volumen de un pipeteador de un solo canal equipado con una punta de pipeta de orificio ancho a 20 μL y aspire cuidadosamente un solo esferoide. Coloque la punta directamente en el centro de cada pozo de la microplaca de ensayo de esferoides y permita que la gravedad eluya un solo esferoide en el centro de cada pozo, es decir, no expulse ningún medio de la punta de la pipeta y permita que la acción capilar retire el esferoide de la punta de la pipeta. Para confirmar la elución, el contenido del pipeteador se puede volver a pipetear en la placa de Petri de 7 cm bajo el microscopio.
NOTA: La elución por gravedad de un solo esferoide generalmente toma de 15 a 30 s dependiendo del tamaño / densidad del esferoide. Durante este tiempo, el pipeteador no debe ser removido. Cualquier pozo de corrección de fondo debe estar libre de esferoides y solo contener medio de ensayo. Bajo el microscopio, confirme la posición de cada esferoide. Idealmente, cada esferoide debe colocarse dentro del centro de cada pozo. - Una vez que todos los esferoides se hayan transferido a la microplaca de ensayo de esferoides, transfiera la placa a una incubadora sin CO2 a 37 ° C durante un mínimo de 1 h antes del ensayo.
- Ajuste el volumen de un pipeteador de un solo canal equipado con una punta de pipeta de orificio ancho a 20 μL y aspire cuidadosamente un solo esferoide. Coloque la punta directamente en el centro de cada pozo de la microplaca de ensayo de esferoides y permita que la gravedad eluya un solo esferoide en el centro de cada pozo, es decir, no expulse ningún medio de la punta de la pipeta y permita que la acción capilar retire el esferoide de la punta de la pipeta. Para confirmar la elución, el contenido del pipeteador se puede volver a pipetear en la placa de Petri de 7 cm bajo el microscopio.
- Usando un microscopio de disección y un aparato de caja de luz, transfiera los esferoides de la placa de Petri a la microplaca de ensayo de esferoides como se detalla a continuación.
3. Preparación y carga de compuestos en el cartucho del sensor para ensayos XF
| Estrategia de inyección | Compuesto (Puerto) | Volumen de arranque del micropocillo XFe96 (μL) | Concentración de pozo final deseada | Volumen de puerto (μL) | Volumen final de micropocillos XFe96 después de la inyección (μL) | Concentración de existencias de trabajo |
| 1 | Oligomicina (A) | 180 | 3 ug/ml | 20 | 200 | 30 μg/ml |
| Rotenona (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenona (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabla 2: Concentraciones de compuestos mitocondriales para sondear el metabolismo energético mitocondrial de esferoides 3D individuales utilizando el analizador XFe96.
- Prepare las concentraciones de material de trabajo de cada compuesto como se indica en la Tabla 2 utilizando un medio de ensayo XF RPMI totalmente suplementado y precalentado.
- Oriente la placa del cartucho (acoplada a la placa de utilidad) en cuanto a la columna, 1-12 de izquierda a derecha.
- Si utiliza una guía de carga, colóquela encima de la placa del cartucho de acuerdo con el procedimiento de carga del pozo, por ejemplo, si el puerto A se carga primero, asegúrese de que A sea visible en la esquina superior izquierda de la guía.
- Transfiera la solución de trabajo de cada compuesto a un depósito adecuado y, utilizando una pipeta multicanal P100 calibrada, dispense 20 μL en todos los puertos correspondientes. Repita para cada compuesto en los puertos restantes.
NOTA: Si no se utiliza algún puerto en la placa del cartucho del sensor, estos pueden dejarse vacíos o llenarse con un medio de ensayo. Si sólo se utiliza una selección de una carta de puerto específica, asegúrese de que los otros puertos correspondientes a esa letra estén cargados con un medio de ensayo; de lo contrario, se inyectará aire en el pozo, comprometiendo los resultados en esos pozos. - Después de la carga en el puerto, retire las guías de carga de la placa (si se utilizan) y prepare el analizador para cargar el cartucho del sensor.
NOTA: Si el ensayo no se está ejecutando inmediatamente después de cargar los puertos, vuelva a colocar la tapa en el cartucho del sensor y vuelva a colocar la placa en la incubadora de aire a 37 °C hasta que esté lista para cargar en la máquina.
4. Diseño de ensayos, estrategias de inyección y adquisición de datos
- Ejecución del ensayo
- Encienda el analizador y conéctelo al controlador (computadora).
NOTA: Esto se puede verificar mediante el estado de conexión del instrumento en el panel de widgets del software Wave Controller. - Navegue a la página de plantillas en el software WAVE, busque el archivo de plantilla de ensayo para el experimento y haga doble clic para abrirlo.
Nota : si la plantilla de ensayo no aparece en la vista Plantillas , importe el archivo de plantilla en la carpeta de plantilla desde una unidad de red compartida o unidad flash USB. - Para iniciar el ensayo, haga clic en la ficha Ejecutar ensayo .
NOTA: Si las definiciones de grupo se han asignado correctamente dentro del mapa de la placa, el ensayo estará listo para ejecutarse como lo indica la marca verde en el lado derecho de la página. En esta etapa, cualquier información adicional se puede ingresar en la página de resumen del ensayo o en la página que se deja en blanco; continúe con el siguiente paso. Debido a la penetración retardada de moduladores mitocondriales en esferoides microtisulares 3D (Figura 2), utilice la información del protocolo de medición descrita en la Tabla 3.
- Encienda el analizador y conéctelo al controlador (computadora).
| Período de medición | Número de inyección y puerto | Detalles de la medición | Duración del período (h:min:s) |
| Calibración | No aplicable | Los analizadores XF siempre realizan esta calibración para asegurarse de que las mediciones sean precisas | 00:20:00 (esto es un promedio y puede variar entre máquinas) |
| Equillibración | No aplicable | El equilibrio ocurre después de la calibración y se recomienda. | 00:10:00 |
| Basal | No aplicable | Ciclos = 5 | 00:30:00 |
| Mezcla = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Oligomicina / BAM15 | Inyección 1 (puerto A) | Ciclos = 10 | 01:00:00 |
| Mezcla = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Rotenona + antimicina A | Inyección 2 (Puerto B) | Ciclos = 10 | 01:00:00 |
| Mezcla = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Tiempo total: | 03:00:00 | ||
Tabla 3: Configuración del protocolo para sondear el metabolismo energético mitocondrial de esferoides 3D individuales utilizando el analizador XFe96.
- Haga clic en Iniciar ejecución para abrir el cuadro de diálogo Guardar ubicación .
- Introduzca la ubicación de guardado para el archivo de resultados y coloque el cartucho del sensor ensamblado en la bandeja térmica que aparece desde la puerta en el lateral del analizador. Espere a que la bandeja térmica se abra automáticamente y la pantalla muestre el mensaje De la placa de utilidad del calibrador de carga . Antes de seguir las indicaciones en pantalla, asegúrese de i) el ajuste adecuado del cartucho del sensor en la placa de utilidad, ii) la tapa se retira del cartucho del sensor y iii) la orientación correcta del cartucho del sensor en la placa de utilidad.
- Siga los comandos en pantalla para iniciar la calibración del cartucho del sensor.
NOTA: El tiempo necesario para completar la calibración es de aproximadamente 10-20 min (para ensayos a 37 °C). - Después de la calibración del cartucho del sensor, cargue la microplaca esferoide en el analizador siguiendo las instrucciones en pantalla del controlador de onda para iniciar el paso de equilibrio de 12 minutos.
NOTA: Las casillas verdes con marcas blancas indican una calibración "buena" para ese pozo. Si algún pozo no proporciona una calibración "buena", se indicará con una caja roja y una cruz blanca. Dichos pozos deben anotarse y excluirse de cualquier análisis después de que se complete el ensayo utilizando la pestaña de ensayo de modificación . - Espere a que el analizador comience automáticamente a adquirir mediciones de referencia después de que la máquina haya completado el paso de equilibrio (como se describe en el protocolo del instrumento).
- Para completar el experimento, siga los comandos en pantalla del controlador WAVE.
NOTA: Una vez que la microplaca esferoide se haya retirado del analizador, deseche el cartucho del sensor y reserve la placa esferoide para un análisis adicional si es necesario (por ejemplo, cuantificación de ADN de doble cadena (ds)). Si la microplaca no es necesaria para un análisis posterior, se puede desechar junto con el cartucho del sensor. - Espere a que aparezca el cuadro de diálogo de ensayo y vea los resultados o vuelva a la vista de plantillas .
5. Estrategias de normalización y análisis de datos: normalización posterior al ensayo y ensayos posteriores (pasos opcionales)
- Normalización de datos
- Para normalizar los datos de esferoides, consulte la serie de protocolos pertinentes a las estrategias de normalización de datos para calcular el tamaño y el volumen de los esferoides y cuantificar el dsDNA en ensayos de esferoides. Estos se han incluido como archivos complementarios; consulte el archivo complementario 1 y el archivo complementario 2.
- Análisis de datos
- Para exportar datos a uno de los generadores de análisis automatizados, siga los comandos de exportación de datos en el controlador WAVE y seleccione el generador de exportación que coincida con el tipo de ensayo. Alternativamente, exporte el archivo de datos y cárguelo en Seahorse Analytics.
NOTA: La desventaja de los generadores de informes y el análisis de Seahorse es que el análisis de datos se limita a cómo está diseñado el ensayo XF y no permite que se tomen promedios a través de los ciclos de medición. La exportación manual de conjuntos de datos desde el software del instrumento permite la preferencia del usuario a este respecto. Dado que la estrategia de inyección para evaluar la respiración mitocondrial de los esferoides 3D probablemente diferirá de la de una prueba típica de 'MitoStress', se han desarrollado una serie de plantillas de hojas de cálculo para ayudar a analizar estos conjuntos de datos, específicos para cultivos celulares 3D y se proporcionarán a pedido. Estos archivos de plantilla de datos proporcionarán datos sobre los parámetros respiratorios mitocondriales clave detallados y explicados en la Figura 2. - Para analizar los datos, exporte los datos como un informe de hoja de cálculo desde el software del controlador WAVE y utilice una plantilla de hoja de cálculo independiente para el análisis.
- Para exportar datos a uno de los generadores de análisis automatizados, siga los comandos de exportación de datos en el controlador WAVE y seleccione el generador de exportación que coincida con el tipo de ensayo. Alternativamente, exporte el archivo de datos y cárguelo en Seahorse Analytics.

Figura 2: Descriptores esquemáticos para parámetros derivados de análisis de datos de flujo extracelular. Abreviatura: OCR = tasa de consumo de oxígeno. Haga clic aquí para ver una versión más grande de esta figura.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Para obtener esferoides compactos y bien formados, cada línea celular se optimizó individualmente para la densidad de siembra y la duración del cultivo (Figura 3). Las líneas celulares A549, HepG2/C3A y SK-OV-3 inicialmente formaron agregados sueltos que no progresaron a esferoides redondos con perímetros claramente definidos hasta después de 7 días en cultivo. Por el contrario, las células MCF-7 podrían formar esferoides en 3 días. Hubo una clara correlación entre la densidad inicial de siembra celular y el volumen esferoide después del período de cultivo para todos los modelos esferoides. El tamaño y la morfología de los esferoides se optimizaron para la densidad de siembra. La morfología y la circularidad comenzaron a disminuir con el aumento del tamaño de los esferoides en todos los modelos. Las estrategias de siembra para líneas celulares se optimizaron a 4 × 103 células/pozo para células A549 y SK-OV-3; Las células HepG2/C3A se han optimizado previamente en otros lugares a 1 × 103 células/pozo, y las células MCF-7 se utilizaron a 4 × 103 células/pozo en todos los ensayos. En las estrategias de siembra optimizadas, el volumen de esferoides estuvo entre 5,46 × 107 μm3 (SK-OV-3) y 1,45 × 108 μm3 (A549) (Figura 3B). Todos los tipos de esferoides tuvieron una correlación lineal entre la densidad de siembra inicial y el volumen de esferoides, donde A549 y HepG2/C3A tuvieron valores de R2 de 0,957 y 0,947, respectivamente. Se encontró que los volúmenes esferoides MCF-7 y SK-OV-3 tenían una mayor correlación con la densidad de siembra inicial, R2 = 0.977 (Figura 3A).
La circularidad esferoide se calculó utilizando planimetría de imagen dentro del software de análisis FIJI utilizando los diámetros esferoides largos y cortos. La simetría esferoide perfecta tenía circularidad = 1.0; la desviación de 1.0 indicó una pérdida de circularidad (Figura 3C). La circularidad fue mayor en los esferoides MCF-7 que en otros modelos donde la circularidad se mantuvo entre 0,83 y 0,9 en todas las densidades de siembra. En comparación, el perímetro exterior de los esferoides SK-OV-3 no estaba tan claramente definido, y el volumen de esferoides fue significativamente menor incluso después de 7 días en cultivo, produciendo esferoides con una circularidad máxima de 0,61 a una densidad de siembra de 4 × 103/pozo. También se encontró que las células HepG2 / C3A forman esferoides apretados y bien formados con una morfología uniforme en el área de superficie de todos los esferoides, con una circularidad mantenida en 0.79 para las células sembradas a 1 × 103 células / pozo. Las células A549 parecieron seguir una tendencia en la que la circularidad y la morfología de los esferoides se mejoraron con la densidad de siembra; sin embargo, la circularidad no fue mayor de 0,63 a la densidad utilizada en estos experimentos.
La respiración mitocondrial basal se calculó como OCR medido a partir de esferoides sembrados a 1 × 103, 2 × 103, 4 × 103, u 8 × 103 células/pozo en microplacas de cultivo de esferoides de unión ultrabaja (Figura 3D). Para todos los tipos de esferoides, el OCR aumentó con el tamaño de los esferoides y se correlacionó linealmente con el volumen de los esferoides con R2 más alto en los esferoides MCF-7 en 0.988 y más bajo en los esferoides SK-OV-3 en 0.744 (Figura 3E). El OCR medido fue estadísticamente diferente entre todos los grupos experimentales. A549 tenía el OCR más bajo, logrando solo 18 pmol / min / pozo en el tamaño de esferoide más grande (Figura 3D). Por el contrario, los esferoides MCF-7 produjeron un OCR similar en el tamaño de esferoide más pequeño después de solo 3 días en cultivo, alcanzando un OCR basal máximo de 53 pmol / min / pozo para el tamaño de esferoide más grande (Figura 3D). HepG2/C3A produjo datos de OCR altamente consistentes con el tamaño y la morfología de los esferoides. En los esferoides HepG2/C3A sembrados de 1 × 103 células/pozo, el OCR basal alcanzó un promedio de 15 pmol/min/pozo, aumentando a un máximo de 52 pmol/min/pozo en los esferoides más grandes (Figura 3D). El OCR en los esferoides SK-OV-3 solo fue significativo entre los esferoides cultivados de 4 × 103 células/pozo y 8 × 103 células/pozo, con poca diferencia observada en los esferoides cultivados entre 1 × 103, 2 × 103, o 4 × 103 células. A pesar de las diferencias de tamaño, los datos de OCR fueron muy similares entre los esferoides HepG2/C3A y MCF-7 en todos los puntos de tamaño. En relación con el tamaño de los esferoides (μm3), el OCR basal de los esferoides MCF-7 fue comparable al de los esferoides HepG2/C3A cultivados durante 7 días a partir de 1.000 células por pozo.

Figura 3: Los parámetros de crecimiento esferoide determinan la respiración mitocondrial basal. (A) Los esferoides A549, HepG2/C3A, MCF-7 y SK-OV-3 se optimizaron para la densidad de siembra y su crecimiento se monitorizó a 1 × 103, 2 × 103, 4 × 103 y 8 × 103 células/pozo en cada una de las fotomicrografías, de arriba a la izquierda a la parte inferior derecha, respectivamente; barras de escala = 500 μm. (B) El tamaño de los esferoides se calculó utilizando datos planimétricos de fotomicrografías recolectadas y se comparó utilizando la estadística de correlación de Pearson; las líneas punteadas representan la distribución del intervalo de confianza del 95%. (C) La morfología esferoide se comparó mediante el cálculo de la circularidad. (D) El OCR se midió 5x, después de lo cual se agregó rotenona-antimicina A para dar cuenta de la frecuencia respiratoria no mitocondrial utilizando el analizador Agilent Seahorse XFe96. OCRbasal medido como OCR - OCRr/a se comparó entre la densidad de siembra (D) y el volumen esferoide (E). Los datos son promedios ± SEM de 5 a 8 réplicas de pozos por tipo de esferoide y densidad de siembra celular. Abreviaturas: OCR = tasa de consumo de oxígeno; OCRbasal = Respiración mitocondrial basal; OCRr/ a = OCR después de la adición de rotenona-antimicina A. Haga clic aquí para ver una versión más grande de esta figura.
La concentración y el curso de tiempo para la exposición a moduladores respiratorios en el análisis XF es un paso crítico en la optimización del ensayo. Los compuestos moduladores respiratorios-oligomicina, BAM15, una mezcla de rotenona-antimicina A-o un control de vehículo DMSO se inyectaron secuencialmente a través de los puertos de inyección del cartucho del sensor en los pocillos de microplacas que contienen los esferoides MCF-7 (Figura 4A). Se completaron cuatro ciclos de medición para determinar el OCR basal promedio de 30-40 pmol / min / pozo para todos los grupos de muestra. Durante el resto de la duración del ensayo, se agregaron secuencialmente moduladores respiratorios cada 5 ciclos de medición para alcanzar concentraciones finales de pozo de 0,5 μM (0,5 μg/ml de oligomicina) después de la inyección 1; 2,0 μM (2 μg/ml de oligomicina) después de la inyección 2; 5 μM (5 μg/ml de oligomicina) después de la inyección 3; y finalmente, una concentración máxima de pozo de 11 μM (11 μg/ml de oligomicina) después de la cuarta adición secuencial. Los esferoides MCF-7 no respondieron al control del vehículo durante todo el experimento (Figura 4A). El OCR basal cambió inmediatamente después de la primera inyección de cada compuesto respectivo a la concentración más baja de 0,5 μM o 0,5 μg/ml de oligomicina (Figura 4B). La OCR en los esferoides MCF-7 se redujo con oligomicina de 41 pmol/min/pozo a 23 pmol/min/pozo después de 5 ciclos de medición después de la primera inyección de 0,5 μg/ml (Figura 4B).
En respuesta a 0,5 μm bam15, el OCR se incrementó de 33 a 41 pmol/min/mucho antes de la segunda inyección (Figura 4C). Comparativamente, la combinación de rotenona más antimicina A redujo el OCR de 37 a 13 pmol/min/mucho antes de la segunda inyección (Figura 4D). Los rastros cinéticos revelaron además una disminución lineal constante (oligomicina y rotenona-antimicina A) o un aumento (BAM15) en OCR. Para todos los regímenes de dosificación de compuestos, se logró un OCR en estado estacionario dentro de 10-12 ciclos de medición completos (60-72 min) a una concentración total de pozo de 2 μM BAM15, 2 μM de rotenona, 2 μM de antimicina A y 2 μg/ml de oligomicina (Figura 4A). La tasa de consumo de oxígeno alcanzó una meseta de estado estacionario a ~ 19 pmol / min / pozo (oligomicina), 52 pmol / min / pozo (BAM15) y 10 pmol / min / pozo (rotenona-antimicina A) (Figura 4A). El aumento de la concentración compuesta de oligomicina, BAM15 o rotenona más antimicina A tampoco tuvo un efecto obvio sobre el OCR, que se mantuvo constante durante el resto del ensayo. Estos datos demuestran que tanto la concentración de compuestos como el curso temporal de exposición a compuestos moduladores respiratorios deben considerarse para la optimización del ensayo cuando se utilizan esferoides 3D.

Figura 4: Titulación de compuestos moduladores respiratorios como un paso importante para optimizar el análisis de flujo extracelular. (A) Los esferoides MCF-7 se sembraron a 4 × 103 células / pozo y se cultivaron durante 3 días antes de colocarse en pozos de una microplaca de ensayo de esferoides que contiene XF RPMI y se sondearon para OCR ± moduladores mitocondriales utilizando el analizador XFe96. La OCR se midió 5x, después de lo cual se agregaron valoraciones de control del vehículo, oligomicina (B), BAM15 (C) o rotenona-antimicina A (D) para inhibir la ATP sintasa mitocondrial, determinar la capacidad respiratoria máxima o establecer la frecuencia respiratoria no mitocondrial, respectivamente. La concentración de cada modulador mitocondrial se incrementó a lo largo de 4 estrategias individuales de inyección de titulación (0,5 μM, 1,5 μM, 3 μM y 6 μM; las unidades para oligomicina son μg/mL) para determinar el OCR máximo en estado estacionario en respuesta a la concentración óptima del compuesto. El OCR se midió durante 5 ciclos de medición entre cada inyección. Los datos son promedios ± SEM de 5 a 8 réplicas de pozos individuales. Abreviatura: OCR = tasa de consumo de oxígeno. Haga clic aquí para ver una versión más grande de esta figura.
Uno de los principales beneficios de la tecnología XF es la capacidad de sondear la función mitocondrial en células y tejidos intactos. Para examinar aspectos específicos de la función mitocondrial en células y tejidos, los moduladores mitocondriales se agregan secuencialmente a los pocillos de la microplaca de muestra a través de los 4 puertos de inyección disponibles en el cartucho del sensor. La secuencia típica de moduladores utilizados para sondear los parámetros mitocondriales en los ensayos de XF son la oligomicina, un protonóforo (por ejemplo, FCCP o BAM15) y una combinación de rotenona más antimicina A, que se agregan secuencialmente para inhibir la ATP sintasa mitocondrial, determinar la capacidad respiratoria máxima y corregir la frecuencia respiratoria no mitocondrial, respectivamente. Esta secuencia típica de adiciones de moduladores se denomina prueba MitoStress por el fabricante de la tecnología de ensayo. Dado que la oligomicina puede inhibir la respiración no estimulada por el desacoplador en algunas monocapas celulares20, examinamos esto con esferoides 3D derivados del cáncer midiendo el OCR no estimulado (OCRmax) antes (único) y después de la inyección de oligomicina (secuencial) (Figura 5A-D). OCRmax no se limitó significativamente por la adición de oligomicina en esferoides formados a partir de HEPG2/C3A o SK-OV-3 (Figura 5E y Figura 5G). Sin embargo, el OCRmáx se redujo significativamente en los esferoides A549 y MCF-7 después de una inyección secuencial de BAM15 después de la oligomicina en comparación con elOCR máximo logrado a partir de una sola inyección de BAM15 (Figura 5F y Figura 5H). A menos que se sepa lo contrario, se recomienda usar pocillos separados para tratar con oligomicina y desacoplador, con una adición final de rotenona y antimicina A cuando se explora el metabolismo energético mitocondrial de los esferoides 3D. Este enfoque aún permite el cálculo de todos los parámetros mitocondriales como con una prueba típica de MitoStress donde los compuestos se agregan secuencialmente.

Figura 5: Inyección única o secuencial de compuestos respiratorios mitocondriales. Los esferoides derivados de células cancerosas de MCF-7, HEPG2/C3A, SK-OV-3 y A549 se colocaron en pozos de una microplaca esferoide XFe96 en XF RPMI y se sondearon para OCR utilizando el analizador Agilent Seahorse XFe96. El OCR se midió 5x, después de lo cual 2 μg/ml de oligomicina (inyección Puerto A: traza verde) o 5 μM BAM15 (inyección Puerto A: traza azul o puerto de inyección B: traza verde) para inhibir la ATP sintasa mitocondrial y determinar la capacidad respiratoria máxima, respectivamente. Los datos de OCR cinética se expresan como % basal (A-D). La capacidad respiratoria máxima (OCRmax) se calculó como un factor de OCR basal mediante la ecuación: OCRmax = OCRBAM15 / OCRbasal. OCRmax se obtuvo a partir de promedios de OCR a través de ciclos de medición 8-10 después de la inyección bam15 con (barras verdes) y sin (barras azules) oligomicina. Los datos son promedios ± SEM de 3-8 pozos individuales replica a través de la microplaca de ensayo de esferoides. Abreviaturas: OCR = tasa de consumo de oxígeno. Haga clic aquí para ver una versión más grande de esta figura.
Utilizando densidades óptimas de siembra celular, concentraciones de compuestos, estrategia de inyección y período del ciclo de medición determinado en estos experimentos de optimización (Tabla 3), hemos desarrollado un protocolo detallado para sondear con precisión la respiración mitocondrial basal: OCRbasal (Figura 6A), respiración de fosforilación ADP: OCRADP (Figura 6B), respiración por fugas: OCRomy (Figura 6C), eficiencia de acoplamiento (Figura 6D ), capacidad respiratoria máxima: OCRmax (Figura 6E), y capacidad respiratoria sobrante: OCRspare (Figura 6F) utilizando esferoides 3D derivados del cáncer.

Figura 6: Sondeo de OCR con tecnología XF para establecer el metabolismo energético mitocondrial de los esferoides derivados del cáncer. Los esferoides derivados de células cancerosas de MCF-7, HEPG2 / C3A, SK-OV-3 y A549 se colocaron en pozos de una microplaca de ensayo de esferoides en XF RPMI y se sondearon para OCR utilizando el analizador Agilent Seahorse XFe96. La OCR se midió 5x, después de lo cual se agregaron 2 μg/ml de oligomicina, o 5 μM BAM15, y AR para inhibir la ATP sintasa mitocondrial, determinar la capacidad respiratoria máxima y calcular la frecuencia respiratoria no mitocondrial, respectivamente. (A) La respiración mitocondrial basal (OCRbasal) se calculó como el promedio de OCR de los 3 ciclos de medición antes de la inyección en el puerto A. (B) La eficiencia de acoplamiento de la fosforilación oxidativa se aproximó expresando OCRADP (OCRbasal- fuga deOCR) como porcentaje de OCRbasal. (C) La respiración por fosforilación de ADP (OCRADP) se midió como OCR sensible a la oligomicina, calculada a partir del OCR promediado a través de los ciclos de medición 11-13 antes de la inyección de BAM15. (D) La respiración por fugas (fuga de OCR) se midió como OCR insensible a la oligomicina, calculada a partir de la media promediada de OCR a través de los ciclos de medición 11-13. (E) La capacidad respiratoria máxima (OCRmáx.) se midió como el OCR máximo promedio medido después de la inyección de BAM15. (F) La capacidad respiratoria excedentaria se calculó expresando OCRspare (OCRmax - OCRbasal) como porcentaje de OCRbasal. La OCR después de la inyección de rotenona-antimicina A (OCRr/a) se restó de todas las tasas para corregir la OCR no mitocondrial. Los datos son promedios ± SEM de 3-8 pozos individuales replicados a través de la placa esferoide XFe96. Abreviaturas: OCR = tasa de consumo de oxígeno; AR = 2 μM rotenona-2 μM antimicina A. Haga clic aquí para ver una versión más grande de esta figura.
Los esferoides MCF-7 cultivados a partir de 4 × 103 células / pozo durante 3 días se utilizaron como modelo para determinar la transferencia, colocación y análisis óptimos dentro de las microplacas de ensayo de esferoides. Utilizando las dimensiones proporcionadas para la microplaca esferoide del fabricante, la superficie del pozo se dividió en tres áreas de zona para una colocación óptima de los esferoides (Figura 7A), donde la zona 1 se destacó como la zona óptima en el centro del pozo. Con un pipeteo cuidadoso utilizando puntas de pipeta de orificio ancho, los esferoides se transfirieron a las placas esferoides y se distribuyeron aleatoriamente a través de las superficies de los pozos por elución por gravedad (Figura 7B). Donde los esferoides se transfirieron cuidadosamente utilizando la elución por gravedad, la mayoría de los esferoides se podían encontrar típicamente en las zonas 1-2 de la microplaca, utilizando las técnicas de transferencia recomendadas por el fabricante. Donde los esferoides fueron forzados a salir de la punta de la pipeta por aspiración, los esferoides a menudo se colocaron más allá de estas zonas y no se podían ver usando microscopía.
Para comparar las posiciones de colocación de los esferoides, los esferoides MCF-7 se transfirieron a las microplacas de ensayo de esferoides en las zonas designadas 1-3 o fuera de la zona (Figura 7A). Estos 4 pocillos fueron rastreados a través de un experimento cinético OCR al inicio y después de la adición de oligomicina, BAM15 o rotenona-antimicina A (Figura 7C). La OCR se calculó a partir de la media de tres lecturas de ciclo antes de cada inyección (Figura 7B). El OCR se midió cinéticamente durante 200 min en los 4 pozos seleccionados (Figura 7C) y se corrigió la línea de base (Figura 7D). Cuando los esferoides se colocaron en la zona 3 o fuera de la zona, los OCR basales fueron significativamente más bajos que los esferoides colocados en las zonas 1 y 2 (Figura 7C). Los efectos de los compuestos respiratorios oligomicina, BAM15 y rotenona-antimicina A también difirieron dramáticamente entre los esferoides colocados en las zonas 1 y 2 en comparación con la zona 3 y las regiones fuera de la zona. Se observó un aumento en la OCR con oligomicina en esferoides colocados en la zona 3 o fuera de la zona (Figura 7E). Además, los esferoides colocados en la zona 3 o fuera de la zona experimentaron una respuesta excesivamente alta a BAM15 con OCR superior al valor basal después de la inyección de rotenona-antimicina A (Figura 7E). A pesar de un aumento de casi el doble en el OCR basal (Figura 7C) con esferoides colocados en la zona 2 frente a la zona 1, los cambios de pliegue en la respuesta a todos los compuestos respiratorios fueron muy similares (Figura 7E), lo que sugiere que es poco probable que las diferencias en el OCR basal entre los esferoides colocados en las zonas 1 o 2 sean el resultado de la colocación dentro del pozo.

Figura 7: La colocación de esferoides dentro de la microplaca de ensayo de esferoides dicta los efectos basales de OCR y modulador mitocondrial utilizando la tecnología XF. Los esferoides MCF-7 se sembraron a 4 × 103 células / pozo y se cultivaron durante 3 días antes de colocarse en los pozos de la microplaca esferoide que contiene XF RPMI y se sondearon para OCR ± moduladores mitocondriales utilizando el analizador Agilent Seahorse XFe96. (A) Fotomicrografías de las posiciones de la zona esferoide en microplacas de ensayo de esferoides después de la duración del ensayo; barra de escala = 500 μm y OCR capturado de los pozos correspondientes a lo largo del tiempo expresado como pmol/min-1/pozo-1 (B) o % basal (C). (D) Efectos moduladores mitocondriales de los esferoides MCF-7 colocados en diferentes zonas dentro de la microplaca de ensayo de esferoides; datos expresados como cambio de pliegue de basal. (E) Ejemplo de traza cinética que destaca qué mediciones de datos de OCR (círculos rojos) se utilizan para calcular la respuesta de cada modulador mitocondrial para los datos presentados en E. Los datos que se muestran provienen de respuestas individuales de pozos. Abreviatura: OCR = tasa de consumo de oxígeno. Haga clic aquí para ver una versión más grande de esta figura.
Los criterios de selección para los antecedentes son de gran importancia; el uso de pozos más externos para la corrección de fondo no es representativo de todos los pozos de microplacas, lo que puede dar lugar a suposiciones de datos incorrectas y conclusiones erróneas de los datos debido a los efectos de borde en toda la microplaca esferoide. Para evaluar esta observación, se utilizaron esferoides MCF-7 para comparar los procedimientos de corrección del ensayo para derivar los valores de OCR en respuesta a la adición de un vehículo de control, oligomicina, BAM15 o rotenona-antimicina A (Figura 8). Todos los compuestos respiratorios produjeron los perfiles cinéticos de OCR esperados para los compuestos seleccionados, revelando una tasa de respiración basal constante promedio de 20-30 pmol / min / pozo (Figura 8A). Sin embargo, cuando los datos del ensayo se analizaron utilizando los pozos más externos para la corrección de la temperatura de fondo (A1, A12, H1 y H12), los valores revelados para OCR después de la adición de compuestos respiratorios fueron especialmente bajos; El OCR arrojó valores negativos para la rotenona-antimicina A. En respuesta a estas observaciones, se realizó un análisis alternativo utilizando una serie de pozos vacíos, distribuidos aleatoriamente a través de la microplaca esferoide, como pozos de corrección de temperatura de fondo (Figura 8B). Cuando se aplicó una corrección de fondo alternativa, todos los efectos compuestos relativos sobre la OCR fueron los mismos en ambos conjuntos de análisis; sin embargo, los valores absolutos de OCR aumentaron en aproximadamente 10 pmol/min/pozo (Figura 8). Estos datos resaltan el poder y la importancia de la corrección de la temperatura de fondo en las microplacas de ensayo de esferoides y enfatizan la importancia de la optimización del usuario para el análisis XF.

Figura 8: Selección aleatoria de pozos para la corrección de fondo para mejorar el control de los gradientes de temperatura a través de la microplaca de ensayo de esferoides. Datos de OCR extrapolados de la Figura 2A utilizando pozos recomendados para la corrección de fondo (A) versus pozos asignados al azar para la corrección de fondo (B). Abreviatura: OCR = tasa de consumo de oxígeno. Haga clic aquí para ver una versión más grande de esta figura.
A diferencia de las monocapas celulares, los esferoides representan una agregación heterogénea de células en un espacio 3D y, por lo tanto, requieren una consideración exhaustiva con respecto al análisis, particularmente al normalizar estos datos. Este artículo presenta tres enfoques para normalizar los datos de XF adquiridos a partir de esferoides MCF-7 (Figura 9). Cuando no se normaliza, el OCR se correlaciona positivamente (R2 = 0,98) con el tamaño de los esferoides (determinado por la densidad inicial de siembra celular) significativamente cuando se compara estadísticamente con el coeficiente de correlación de Pearson, P = 0,0057 (Figura 9A). Esta relación lineal se reduce cuando el OCR se normaliza a la densidad inicial de siembra celular (R2 = 0,78) y ya no se correlaciona significativamente con el tamaño de los esferoides (P = 0,117, Figura 9B). Este es también el caso cuando se normaliza a volumen esferoide (R2 = 0,77; Coeficiente de correlación de Pearson P = 0,120, Figura 9C) y contenido de ADN nuclear (R2 = 0,58; Coeficiente de correlación de Pearson P= 0,233, Figura 9D). Estos datos resaltan la importancia de normalizar los datos de XF al sondear el metabolismo mitocondrial de los esferoides, especialmente si son de diferentes tamaños.

Figura 9: Normalización de los datos de flujo extracelular adquiridos de esferoides celulares. (A) Los datos de OCR en bruto se obtuvieron de MCF-7 cultivado durante 3 días y se trazaron utilizando el modelo de Pearson para obtener un coeficiente de correlación entre la densidad de siembra de esferoides y OCR; Valor de P establecido en 0,05. (B) Los datos brutos de OCR se normalizaron contra la densidad inicial de siembra de esferoides; (C) Volumen esferoide MCF-7 obtenido de la planimetría microscopía; y (D) el contenido de ADN de ds nuclear comparado utilizando el coeficiente de correlación de Pearson. Abreviaturas: OCR = tasa de consumo de oxígeno; ds ADN = ADN de doble cadena. Haga clic aquí para ver una versión más grande de esta figura.
| Línea celular | Densidad de siembra (pozo) | Crecimiento esferoide (días) | Volumen esferoide final (μM3) | OCR basal (pmolO2/min/pozo) | Sensibilidad para el OCR basal cumplido (SÍ/NO) |
| SKOV | 1000 | 5 | 9.52E+06 | 28 ± 3,5 | Sí |
| SKOV | 2000 | 5 | 2.38E+07 | 26 ± 1,4 | Sí |
| SKOV | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | Sí |
| SKOV | 8000 | 5 | 1.11E+08 | 50 ± 7,9 | Sí |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0,7 | No |
| HepG2 | 2000 | 5 | 2.88E+07 | 23 ± 1,8 | Sí |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1,7 | Sí |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2,8 | Sí |
| A549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Sí |
| A549 | 2000 | 5 | 3.57E+07 | 41 ± 1,6 | Sí |
| A549 | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Sí |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8,4 | Sí |
| MCF-7 | 1000 | 3 | 1.60E+07 | 29 ± 0,8 | Sí |
| MCF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1,7 | Sí |
| MCF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Sí |
| MCF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2,9 | Sí |
Tabla 4: Parámetros optimizados para determinar las mediciones basales de OCR en esferoides 3D individuales. Abreviatura: OCR = tasa de consumo de oxígeno.
Archivo Suplementario 1: Análisis de tamaño y volumen de esferoides. Haga clic aquí para descargar este archivo.
Archivo suplementario 2: Cuantificación del ADN de doble cadena a partir de esferoides en la microplaca esferoide. Haga clic aquí para descargar este archivo.
Archivo suplementario 3: Recomendaciones para el número de réplicas necesarias para obtener conjuntos de datos de ensayo XF confiables. Haga clic aquí para descargar este archivo.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Principales conclusiones y productos
Este artículo proporciona un protocolo detallado para sondear el metabolismo energético mitocondrial de esferoides 3D individuales utilizando una serie de líneas celulares derivadas del cáncer con el analizador XFe96 XF. Se desarrolla y describe un método para el cultivo rápido de esferoides celulares A549, HepG2 /C3A, MCF7 y SK-OV-3 utilizando tecnologías repelentes celulares para la agregación forzada. Este protocolo aborda muchas consideraciones sobre el sondeo del metabolismo de los esferoides con tecnología XF, incluida (1) la optimización de los protocolos de cultivo de esferoides y el manejo y la transferencia de esferoides a microplacas de ensayo de esferoides específicas del fabricante de la tecnología desde sus recipientes de cultivo originales; (2) la concentración de compuestos respiratorios que se utilizarán y la dependencia temporal de la penetración de compuestos; (3) estrategias de inyección que se utilizarán; y (4) formas de normalizar los datos entre grupos experimentales. Todas estas consideraciones se han examinado en el presente documento y se examinan con más detalle a continuación. Estos métodos se presentan como enfoques simplificados para generar datos de flujo de oxígeno metabólico consistente utilizando esferoides 3D individuales con el analizador de flujo XFe96. Este enfoque experimental se puede utilizar como punto de partida y rúbrica para su uso en otros modelos esferoides que se implementan fácilmente dentro de un entorno de laboratorio básico.
Consideraciones
Crecimiento, tamaño y sensibilidad de los esferoides de la tecnología XF
Para establecer datos reproducibles con tecnología XF, es esencial caracterizar y optimizar el ensayo para el modelo específico. Este enfoque es relativamente simple en una monocapa básica de células; sin embargo, esto presenta desafíos adicionales al cultivar células como esferoides 3D. Durante los experimentos presentados aquí, el medio RPMI del fabricante se complementó con el uso. Si bien se observa que algunas líneas celulares, a saber, HepG2 / C3A, se cultivaron en medio de crecimiento DMEM, durante estos ensayos relativamente cortos (~ 3-5 h), la sustitución con formulaciones RPMI-DMEM tuvo un impacto limitado en el análisis XF. La formulación de los dos medios es muy similar, y los usuarios podrían "sintonizar" los medios SEAHORSE RPMI para que coincidan con la matriz de sus medios de cultivo celular a través de la suplementación, por ejemplo, aumento de la glucosa, adición adicional de fuentes de carbohidratos. Es fundamental para la formulación final de todos los tampones y medios de XF la ausencia de rojo fenol, que es probable que interfiera con las sondas fluorescentes dentro de la placa de cartucho de la sonda XF, y bicarbonato de sodio, que conducirá a la alcalinidad debido a la falta de tampón de CO2 presente en las incubadoras de cultivos celulares. Otros medios y búferes se pueden comprar y/o fabricar internamente. Por ejemplo, el tampón Krebs Ringer HEPES es un tampón simple que se puede utilizar para evaluar la respiración en muchas células diferentes, incluidos los modelos de esferoides. Sin embargo, los usuarios de ensayos XF deben tener en cuenta que un cambio en el medio/tampón y su suplementación pueden cambiar su capacidad general de almacenamiento en búfer. Esto es de particular preocupación cuando los usuarios pueden estar interesados en medir ECAR, en el que el factor tampón del medio debe evaluarse para permitir la transformación de ECAR a la tasa de eflujo de protones (PER).
Como el OCR celular medido por la tecnología XF es proporcional a la densidad celular cuando el número de células en el pozo está dentro de la sensibilidad del sistema, era importante investigar esta relación utilizando esferoides 3D individuales. Al sondear el OCR de esferoides 3D individuales cultivados a partir de 4 líneas celulares de cáncer diferentes sembradas a densidades de 1,000, 2,000, 4,000 u 8,000 células por pozo, mostramos que el analizador XFe96 es lo suficientemente sensible como para detectar cambios en la tasa de respiración mitocondrial entre esferoides 3D cultivados a partir de diferentes densidades de siembra celular (Figura 3). Mostramos que el rango óptimo de densidad de siembra celular, y por lo tanto el volumen esferoide para formar esferoides 3D para sondear OCR, difieren según el tipo de célula. Esto se muestra por la relación lineal entre el OCR y la densidad de siembra o volumen esferoide (Figura 3). Para las células A549 y HepG2/C3A, la densidad de siembra óptima para la sensibilidad al OCR fue de entre 1.000 y 8.000 células/pozo; fue de 2.000-8.000 células/pozo para MCF-7 y de 4.000-8.000 células/pozo para células SK-OV-3. Estos datos demuestran que la optimización del tamaño de los esferoides es de particular importancia cuando se evalúa el OCR utilizando la tecnología XF.
Consideraciones sobre volúmenes esferoides mínimos y máximos y OCR basal
En general, siempre habrá umbrales mínimos y máximos para los parámetros de OCR medibles recomendados por el fabricante para estos experimentos. Para el analizador XFe96, el OCR basal entre 20 pmol O2/min/pozo y 200 pmol O2/min/pozo son los límites inferior y superior, respectivamente. Este es el caso de las células monocapa y los esferoides, y donde el modelo experimental se encuentra dentro de este rango dinámico de OCR dependerá de la cantidad de material biológico disponible, por ejemplo, el número de células como monocapa o el tamaño de los esferoides. Consulte la Tabla 4 para ver un ejemplo de cómo se alcanzaron los umbrales de OCR mediante los modelos esferoides utilizados aquí. Puede ser prudente verificar el nivel de oxígeno dentro del pozo para el cual estos datos también están disponibles a partir de estas mediciones como los datos de nivel. Esto debe verse rutinariamente desde cada experimento para fines de control de calidad. Si hay agotamiento de oxígeno en el pozo, esto se hará evidente dentro de los datos. Si este fuera el caso, puede ser necesario ajustar los ciclos de medición dentro del experimento; por ejemplo, aumentar el paso de mezcla de tal manera que el nivel de oxígeno en el pozo se recupere antes del siguiente período de medición dentro del ciclo de medición. Aunque es posible, hemos encontrado que esto es muy poco probable para experimentos de un solo esferoide utilizando las líneas celulares descritas.
Elección de desacopladores mitocondriales para ensayos de flujo extracelular
Los ionóforos de protones, como el cianuro de carbonilo 4-(trifluorometoxi) fenilhidrazona (FCCP)21, el cianuro de carbonilo m-clorofenilhidrazona (CCCP)22 o BAM1523, son potentes sustancias químicas de moléculas pequeñas capaces de interrumpir el gradiente electroquímico de protones a través de las membranas mitocondriales, inhibir la producción de ATP y, en última instancia, desacoplar la respiración mitocondrial24 . Se siguen desarrollando nuevas moléculas pequeñas para estos fines, particularmente en el tratamiento de enfermedades metabólicas 25,26,27; se refieren a dos excelentes críticas28,29. Por el contrario, el desacoplamiento de la respiración oxidativa se ha relacionado con toxicidad indeseable fuera del objetivo30. Sin embargo, dentro de los ensayos celulares in vitro, la molécula FCCP despolariza el potencial de la membrana mitocondrial y ejerce efectos fuera del objetivo, como la despolarización de la membrana plasmática, interrumpiendo el flujo iónico NA+ 31; interferencia con el procesamiento de proteínas celulares32, e incluso inducción de senescencia celular33. BAM15 se introdujo originalmente en 2013 como un desacoplador mitocondrial con una influencia mínima en las membranas plasmáticas23, con actividad protonofórica en el rango micromolar en células enteras y rango nanomolar en mitocondrias aisladas 23,34.
Dada la potencia de FCCP en la despolarización de la membrana plasmática, BAM15 es un protonóforo más confiable para desacoplar la respiración en células enteras intactas en ensayos de flujo extracelular. Aunque FCCP y su contraparte, CCCP, se han utilizado durante más de 50 años para evaluar las capacidades respiratorias máximas y continúan siendo ampliamente utilizados en estudios de XF, el uso de estas moléculas pequeñas a menudo subestima la capacidad metabólica mitocondrial y celular. Esto está en parte relacionado con la razón por la que tantas publicaciones que utilizan la tecnología XF caen en la trampa de informar capacidades respiratorias adicionales negativas o subestimar las verdaderas capacidades respiratorias mitocondriales cuando se utiliza FCCP. La potencia añadida de LA FCCP en células y tejidos intactos a menudo conduce a una función mitocondrial comprometida, y las células pueden tener dificultades para operar adecuadamente para mantener una capacidad respiratoria máxima a través de múltiples ciclos de medición después de su adición, incluso a concentraciones muy bajas35. Por lo tanto, la respuesta de las células a la FCCP se puede encontrar en muchos estudios para disminuir después del período del ciclo de medición inicial. Mientras que FCCP se ha utilizado rutinariamente para el análisis de XF, BAM15 se utiliza preferentemente en casos que involucran células enteras o modelos esferoides, dado que puede mantener una capacidad respiratoria máxima en mitocondrias totalmente despolarizadas a concentraciones tan altas como 10 μM 3. Además, BAM15 induce efectos sobre la acidificación extracelular, que coincide con la oxidación de nutrientes a través de la hidratación del CO2 para formar HCO3- y H+ en mayor medida que FCCP3. Sin embargo, en el caso de mitocondrias aisladas y células permeabilizadas, cualquiera de estos desacopladores debe funcionar tan bien como BAM15 para el desacoplamiento mitocondrial si se titula a la concentración correcta.
Cinética de la penetración de compuestos y el ciclo de ensayo
Las concentraciones, la penetración y los perfiles cinéticos de los compuestos químicos utilizados para realizar una prueba típica de MitoStress con esferoides 3D utilizando el analizador XF son más complejos de abordar. Dado que los esferoides presentan una estructura 3D, la penetración de moléculas a través del diámetro del esferoide es un proceso infinitamente más complejo que a través de las monocapas celulares. Por ejemplo, la penetración cinética y, por tanto, la sensibilidad al sorafenib quimioterapéutico se determinó por la edad esferoide y, por tanto, el tamaño en un esferoide HepG2 modelo36. La capacidad de los productos químicos de moléculas pequeñas (por ejemplo, fármacos, nanopartículas) para alcanzar un objetivo biológico depende de varios factores subyacentes, incluida la complejidad del sistema para ser penetrado dinámicamente y difundido a través de37,38. Esto es particularmente cierto para los medicamentos dirigidos al tejido tumoral39. Similar a la orientación tumoral en el contexto de un esferoide 3D, el tamaño, la compacidad y otras respuestas fenotípicas, como la expresión de proteínas transportadoras de fármacos, pueden gobernar el tiempo de penetración y la concentración de un compuesto necesarios para provocar una respuesta biológica.
En este protocolo, abordamos el tema en torno al tiempo de penetración y la concentración de moléculas pequeñas en respuesta al inhibidor de la ATP sintasa oligomicina, el desacoplador protonóforo y mitocondrial BAM15, y la combinación de los inhibidores del Complejo I y complejo III rotenona y antimicina A. Al sondear el OCR de esferoides MCF-7 individuales expuestos a múltiples titulaciones de estos compuestos respiratorios comunes, demostramos que la concentración óptima de cada compuesto requerida para inducir una frecuencia respiratoria en estado estacionario cae dentro de un rango similar al de las células monocapa (Figura 4). Es importante destacar que, y a diferencia de sus contrapartes monocapa, se muestra que aumentar el número de ciclos de medición entre inyecciones es clave para lograr un OCR de estado estacionario en esferoides 3D individuales. Estos datos resaltan la importancia de la penetración de compuestos y sus respectivos perfiles cinéticos al explorar los parámetros respiratorios mitocondriales de los esferoides 3D utilizando estos enfoques. Utilizando las propiedades de optimización de esferoides, las concentraciones de compuestos y los tiempos de ciclo de medición informados por los datos presentados en la Figura 3, figura 4 y Figura 5, se estableció una prueba MitoStress validada para sondear parámetros específicos del metabolismo oxidativo mitocondrial en una variedad de esferoides 3D derivados del cáncer (Figura 6). De importancia, y al igual que algunas líneas celulares de cáncer monocapa40, la capacidad respiratoria máxima (tasa de respiración desacoplada-estimulada) de ciertos esferoides 3D derivados del cáncer fue inhibida por la oligomicina (Figura 5). Específicamente, los esferoides 3D cultivados a partir de células A549 o MCF-7 mostraron una tasa máxima de respiración significativamente menor cuando se desacoplan con BAM15 después de la inyección de oligomicina en comparación con bam15 sin oligomicina (Figura 5F y Figura 5H). Dado que este efecto puede estar presente en otros cultivos de esferoides 3D, sugerimos que, a menos que se emplee un protocolo previamente validado, la capacidad respiratoria máxima en los esferoides 3D debe estimarse sin oligomicina.
Recopilación simultánea de datos ECAR como medida del flujo glicolítico en esferoides celulares
Como se ve típicamente en la literatura o la información del fabricante de la tecnología, la tasa glucolítica de los esferoides, medida como ECAR, es un parámetro secundario que se puede capturar junto con OCR. El cálculo de ECAR por sí solo no es un parámetro útil o significativo en ningún experimento de XF, ya que no se corrige por la capacidad de amortiguación del tampón de ensayo de XF o la adición de acidificación mitocondrial, que surge de la hidratación de CO2 a HCO3- y H +. ECAR solo es perspicaz una vez que se aplican estas correcciones de datos, después de lo cual es posible proporcionar conclusiones más precisas sobre el flujo glicolítico. Para corregir la capacidad de almacenamiento en búfer para generar datos PER más significativos, se debe conocer el volumen de la microcámara para la microplaca esferoide. El fabricante no ha podido proporcionar un volumen real para esto con la microplaca esferoide y, por lo tanto, los datos PER no se pueden determinar fácilmente. De hecho, aunque estas mediciones podían lograrse empíricamente, esto estaba más allá del alcance de este manuscrito. Sin embargo, con las correcciones apropiadas y conociendo el volumen de la microcámara para un tamaño de esferoide dado presente (por ejemplo, obteniendo una medida de la densidad de esferoides) en el pozo, los datos de ECAR se volverían significativos, y se podrían hacer cálculos de PER glicolítico. Por lo tanto, los datos de XF podrían ser más informativos para investigar el metabolismo glicolítico y oxidativo en los esferoides, pero solo si estos parámetros se consideraran en profundidad.
Formación, manipulación, transferencia y movimiento de esferoides
Algunas líneas celulares son más adecuadas para la formación de esferoides que otras y pueden no formar esferoides en absoluto, por ejemplo, las células de cáncer de ovario MCF-741,42 forman esferoides altamente circulares en comparación con otras líneas celulares (Figura 3). Como otro ejemplo, se ha demostrado que las células de cáncer de páncreas Capan-1 forman mejores esferoides que Panc-1 o BxPC343. Del mismo modo, se sabe que las líneas celulares de carcinoma hepático tienen capacidades variables para formar esferoides compactos 5,44, con un cambio observado en el fenotipo, como el aumento del metabolismo de los fármacos o la producción de albúmina, como es el caso de HepG2 frente a HepG2/C3A 9,45,46 o los esferoides HepaRG 17,47,48 . Por lo tanto, los usuarios deben optimizar las técnicas de cultivo de esferoides en consecuencia y realizar experimentos de titulación para determinar la densidad óptima de siembra y el curso del tiempo de cultivo. Además, se ha demostrado que la formulación y composición de los medios de ensayo afectan la formulación de esferoides, incluida la adición de metilcelulosa, a menudo agregada a los medios para aumentar la viscosidad de la matriz 43,49,50. Por lo tanto, la composición óptima del medio celular debe determinarse empíricamente para todas las líneas celulares utilizadas.
El número de intercambios de medios a lo largo del cultivo de esferoides está determinado por la línea celular utilizada. Sin embargo, por lo general, los intercambios de medio volumen cada 2-3 días son aplicables en la mayoría de los casos para reponer nutrientes. Utilizamos el enfoque de agregado forzado para generar esferoides 3D utilizando microplacas repelentes celulares de fuentes disponibles comercialmente para el rápido desarrollo y despliegue de modelos de esferoides en estudios de análisis XF. Sin embargo, las plataformas alternativas pueden ser más adecuadas para generar esferoides de otros tipos de células, por ejemplo, enfoques colgantes o incrustados en matrices. En laboratorios de recursos limitados, los usuarios pueden desear mirar hacia la técnica de superposición de agarosa-líquido para la formación de superficies de microplacas repelentes celulares51,52 para reducir significativamente los costos económicos de los pasos iniciales de desarrollo del método esferoide. El movimiento de los esferoides entre los vasos de cultivo es necesario para realizar análisis XF y otros ensayos aguas abajo. La facilidad de transferencia suele estar dictada por el tamaño de los esferoides y la densidad general. Recomendamos usar una punta de pipeta de orificio ancho P200 o P1000 para mantener la integridad de los esferoides; Las puntas de pipeta de diámetro más pequeño corren el riesgo de interrupción mecánica del esferoide, que se puede comprar comercialmente o, con cuidado, hacer simplemente recortando el extremo de la punta de la pipeta para aumentar el orificio. Sin embargo, este enfoque puede ser susceptible de introducir furring en el plástico alrededor del extremo de la punta, lo que podría causar interrupciones mecánicas durante la manipulación. El uso de una retroiluminación o caja de luz también es útil para el manejo de esferoides y la observación bajo un microscopio de disección como un paso esencial para garantizar la transferencia exitosa de esferoides a la microplaca de ensayo de esferoides. Además, la posición del esferoide dentro del pozo de una microplaca de ensayo de esferoides es de particular importancia e impacta directamente en el OCR y el efecto compuesto durante una prueba típica de MitoStress (Figura 7), muy probablemente debido a la relación de la posición del esferoide y los fluoróforos de la sonda del sensor.
Corrección de fondo y pozos de control de temperatura
El uso de ensayos basados en microplacas es un enfoque ampliamente utilizado en varias áreas de investigación; sin embargo, su uso presenta varios desafíos prácticos. Como es cierto en otros enfoques experimentales, particularmente aquellos que utilizan el formato de matriz 96 (o superior), la geometría y el posicionamiento de las microplacas pueden influir en la temperatura y los gradientes de intercambio de gases a lo largo del tiempo, a menudo denominados "efectos de borde"53,54. Encontramos que lo mismo es cierto para la microplaca de ensayo de esferoides. Según las directrices y protocolos del fabricante, los pozos de esquina más externos: A1, A12, H1 y H12 siempre se designan como pozos de corrección de fondo y control de temperatura para el analizador XFe96. Por el contrario, con el formato de matriz de 24 pocillos, A1 y D6 se designan como pozos de control, junto con otros dos pozos distribuidos uniformemente en el centro de la placa en B4 y C3. Al realizar el análisis de esferoides XF, encontramos una desviación significativa en los datos recopilados inicialmente utilizando la guía del fabricante. Esto fue a pesar de la inclusión de los pasos necesarios para asegurar el ensayo de preequilibración a la temperatura y el contenido de CO2 antes de comenzar la adquisición de datos, a menudo arrojando valores negativos para OCR después de la inyección de ciertos inhibidores respiratorios (Figura 8).
Encontramos que estas observaciones probablemente se deban a los efectos de borde a través de la microplaca del ensayo de esferoides. En la Figura 8, encontramos que redistribuyendo los pozos de control de fondo a través de la microplaca, los datos XF se ajustaron aproximadamente 2 veces. Dos razones más probables son (1) debido a los efectos de evaporación en los pozos de borde que resultan en una cámara de volumen total más pequeña para que la sonda XFe96 muestree, y (2) a equilibrios de temperatura inadecuados entre los pozos designados para la corrección de fondo y los pozos de muestra, lo que resulta en conjuntos de datos que enmascaran o inflan en exceso el OCR. Por lo tanto, para evitar tales resultados, se recomienda, especialmente en el contexto del análisis de esferoides, que los usuarios redistribuyan los pozos designados para la corrección de fondo en la totalidad de la microplaca de ensayo de esferoides y tomen las medidas necesarias para preequilibrar su ensayo antes de adquirir datos de XF.
Normalización de datos
Además de proporcionar un protocolo detallado para sondear el metabolismo energético mitocondrial de esferoides 3D individuales con tecnología XF, este documento también presenta posibles formas de normalizar los datos de frecuencia respiratoria mitocondrial obtenidos con esferoides 3D. Utilizando datos de frecuencia respiratoria obtenidos con esferoides MCF-7 cultivados a diferentes densidades de siembra celular (Figura 3), presentamos frecuencias respiratorias mitocondriales basales de esferoides MCF-7 de tamaño y diámetro crecientes cuando se normalizan a la densidad inicial de siembra celular, volumen esferoide y contenido de dsDNA (Figura 9). El método de normalización apropiado es primordial para la interpretación precisa de los conjuntos de datos XF, particularmente cuando se comparan modelos de esferoides 3D in vitro y diferentes tipos de células. Una normalización deficiente puede conducir a resultados erróneos que simplemente no se pueden comparar entre conjuntos de datos. El contenido de proteínas no se prefiere para la normalización de los datos de XF esferoides, ya que los pretratamientos pueden afectar las tasas de síntesis de proteínas sin un efecto significativo sobre la frecuencia respiratoria. Además, cantidades significativas e inconsistentes de proteína pueden unirse a microplacas esferoides en la lisis celular, introduciendo variación en el contenido de proteínas entre los pocillos. Esto puede complicarse aún más en los análisis de XF utilizando esferoides o células no adherentes que requieren pegamentos biomoleculares para unirse, que pueden contener proteínas.
Contrariamente al contenido de proteínas intracelulares, el contenido de ADN nuclear es independiente del tipo celular y es proporcional al número celular (Figura 9D), un enfoque más preciso y menos lento que la desagregación de esferoides para la cuantificación del número celular. Por el contrario, Yepéz et al.55, realizando análisis de XF en monocapas de células de fibroblastos, encontraron que la normalización de los datos de XF al número de células introducía una mayor dispersión de datos que antes de la normalización. El contenido de ADN nuclear es independiente del estado diferenciado o fenotipo y, por lo tanto, es más preciso para la normalización de los datos de esferoides en ensayos de XF que el contenido de proteínas. El contenido de ADN también ha sido una estrategia probada para el análisis de otros conjuntos de datos vinculados al metabolismo56. Sin embargo, es importante tener en cuenta que el contenido de ADN nuclear se cuantifica a partir de todas las células presentes dentro del esferoide; por lo tanto, no se recomienda la normalización del contenido de ADN para los conjuntos de datos XF en los que los esferoides se someten a tratamientos que pueden resultar en una pérdida significativa de la viabilidad celular. Para tales conjuntos de datos, si es factible, se prefiere la normalización a la viabilidad celular, o los datos pueden corregirse de referencia a la respiración basal.
Uso de la capacidad respiratoria excedentaria como ejemplo de la importancia de la normalización de datos
La capacidad respiratoria excedentaria es una medida de la tasa de capacidad respiratoria mitocondrial máxima menos la frecuencia respiratoria mitocondrial basal (Figura 6). Sin embargo, el problema con la notificación de datos de este tipo como una tasa, es decir, pmolO2 / min / bien dentro de ciertos experimentos, es que los datos están vacíos de normalización. Incluso si se normalizan los datos esferoides a la densidad celular / contenido de ADN, esto a menudo excluye el parámetro clave que debe normalizarse para la densidad mitocondrial dentro de las células. Dado que un cambio en la densidad mitocondrial conducirá a un cambio proporcional en la respiración basal y máxima, la capacidad ociosa también aumentará. Por ejemplo, si el esferoide OCRbasal es 200 y el OCRmáximo es 400, la capacidad excedentaria se informa como 200; si OCRbasal es 100 y OCRmáximo, entonces la capacidad excedentaria también es 100; sin embargo, como porcentaje, ambos son 50% de máximo (o 100% de basales). Por lo tanto, la capacidad excedentaria no se modifica entre estos dos ejemplos, a pesar de las diferencias en las tasas de 200 y 100 cuando se calculan como pmoles O2/min/pozo. Los valores normalizados internamente son más confiables y perspicaces para hacer que los datos de XF sean más comparables entre estudios y proyectos. Para hacer esto para la capacidad respiratoria sobrante, hemos optado por presentar esto como un porcentaje de la respiración máxima en lugar de una tasa absoluta. Esto también podría presentarse como un porcentaje de la respiración basal. Este sería el caso si se trabaja con células o esferoides. Sin embargo, dado que la ubicación del esferoide en la placa de micropocillos puede alterar el OCR absoluto, pero no los cambios relativos con inhibidores o desacopladores, es más importante observar las respuestas normalizadas internamente en los esferoides como el cambio de pliegue o los porcentajes.
Los modelos esferoides generados aquí presentan una gama de tipos de células y arquitectura que no se pueden capturar en los modelos 2D clásicos. Estos incluyen la disposición espacial heterogénea de las células en tres dimensiones, los contactos célula-célula mejorados (por ejemplo, la formación de uniones de brecha y matrices extracelulares) y los gradientes bioquímicos a través del diámetro del esferoide (por ejemplo, gradientes de pH, acceso de difusión de oxígeno a los nutrientes). El uso del flujo extracelular para estudiar la biología de los esferoides in vitro podría permitir identificar objetivos óptimos para las terapias farmacológicas a través de observaciones de perturbación metabólica. Estos podrían extrapolarse de los esferoides in vitro a los tumores in vivo e identificar las vías que pueden dirigirse al metabolismo del tumor esferoide, por ejemplo, la utilización de carbohidratos durante el crecimiento de los esferoides. Las modalidades terapéuticas pueden ser efectivas para atacar los esferoides en las primeras fases de crecimiento, pero resultan menos efectivas en las fases posteriores del crecimiento de los esferoides a medida que madura la complejidad de la red metabólica. Para concluir, el desarrollo de modelos de cultivo celular 3D y tecnologías de análisis sofisticadas en la investigación biológica continuará siendo un campo dinámico y rápidamente cambiante con un potencial insuperable. El análisis de flujo extracelular de esferoides de cultivo celular in vitro podría emplearse como un método de investigación de vanguardia para avanzar en los resultados de la investigación que podrían extrapolarse para comprender mejor la biología relevante para el ser humano, reducir el uso de modelos animales en la investigación y mejorar la investigación centrada en el paciente.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Los autores no tienen conflictos de intereses que declarar.
Acknowledgments
N.J.C fue apoyado por un premio BBSRC MIBTP CASE con Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Investigación del cáncer número 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
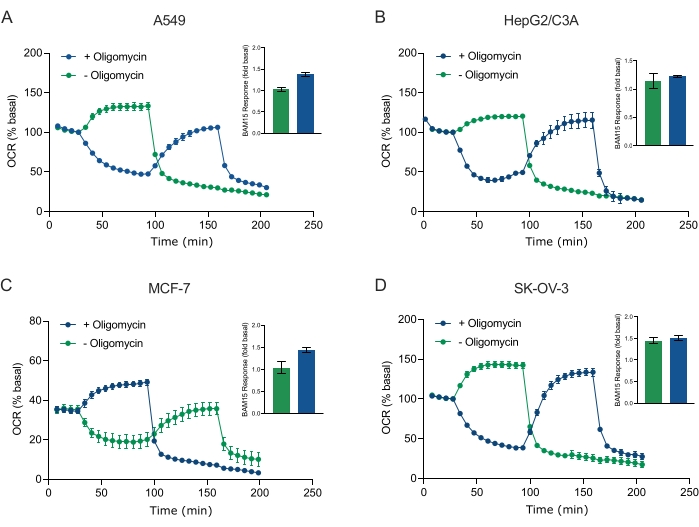
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
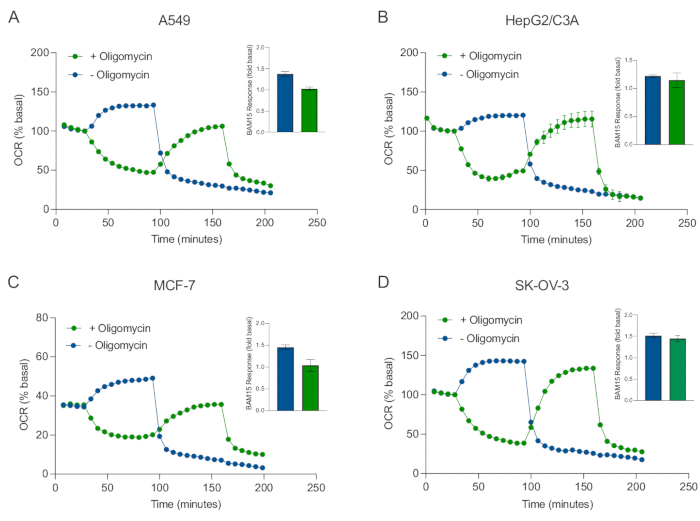
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
