

Summary
Umodificerede og hyperphosphorylerede tau-proteiner blev anvendt i to in vitro aggregation assays til at afsløre hyperphosphoryleringsbegivenheder-afhængige hurtig sammenlægning kinetik. Disse assays bane vejen for fremtidige skærme for forbindelser, der kan modulere tilbøjelighed hyperphosphoryleret tau at danne fibriller, der ligger til grund for progressionen af Alzheimers sygdom.
Introduction
Alzheimers sygdom (AD) er en af en stor samling af neurodegenerative lidelser kendt som tauopatier. Indbegrebet patologi underliggende tauopati er de neurofibrillære sammenfiltringer, NFT'er, i neuroner, astrocytter og mikroglia 1-4. NFT tæthed korrelerer med kognitiv svækkelse 3,5 og neurontab 6. NFT primært indeholder hyperphosphoryleret tau protein (benævnt "p-tau" nu), der danner lige eller parrede spiralfilamenter (PHF) 7,8. Tau er et mikrotubulus-associeret protein menes at lette axonal transport, som er afgørende for neuronal signalering og handel 9,10. Hver tau-molekylet indeholder 2 til 3 fosfater i normal hjerne, men phosphoryl indhold stiger med flere folder i tauopati patienter 11. Flere kinaser vil sandsynligvis bidrage til tau hyperphosphorylering herunder GSK3p (glycogensyntasekinase 3β) og CDK5 (cyclin-degige kinase 5) 12,13, men den direkte udløsende faktor for patologisk phosphorylering stadig vanskeligt at definere 14. Unormal phosphorylering i eller i nærheden af mikrotubulus-bindende motiver dissocierer tau fra mikrotubulus 15, og forårsager tau mis-lokalisering til somatodendritisk rum, hvor p-tau oligomeriserer i lige eller parrede spiralformede filamenter, som eventuelt kan polymerisere i NFT inklusioner. Den tætte forbindelse mellem tau-hyperphosphorylering, NFT formation og neurodegeneration førte til en fremherskende hypotese, at p-tau tangles fremkalde apoptotiske og andre cytotoksiske responser, og dermed er den underliggende årsag til tauopati neurodegeneration 16,17. Drug skærme og tidlige kliniske tests baseret på denne forudsætning er der lanceret 18. Denne hypotese står imidlertid udfordringer 19,20. For eksempel SantaCruz et al., Viste, at de kognitive funktioner af transgene mus kan forbedres ved at undertrykke ekspressionen af et mutanthumant tau, selvom NFT'er fortsat at danne fra eksisterende taumolekyler 21. I en Drosophila model blev NFT vist at sekvestrere toksiske cytosoliske tau at beskytte de underliggende neuronceller 22,23. Det er klart, patogenesen rolle NFT, hvis nogen, i høj grad vil påvirke retningen af tauopati terapeutiske udvikling.
I høje koncentrationer, rekombinante eller normal hjerne tau-protein spontant men langsomt polymeriserer i en PHF-lignende struktur in vitro som angivet ved binding af adskillige β-sheet foretrukne fluorescerende farvestoffer, elektronmikroskopi, og lysspredning spektroskopi 24-27. Tilføjelse heparin eller arachidonsyre, en rigelig fedtsyre i human hjerne, drastisk accelererer PHF-dannelse i tau isoform- og inducer koncentrationsafhængige måder 28-32. Interessant nok hyperphosphoryleret tau oprenset fra AD-hjerner eller fremstillet ved udtømmende in vitro phosphoryleringsreaktioner enggregates hurtigere og mere effektivt 26,33-35. Disse resultater er i fremragende aftale med de patologiske roller p-tau. Et in vitro system baseret på sammenlægning af p-tau kan således fungere som et effektivt redskab til AD drug screening.
I betragtning af den tætte sammenhæng mellem tau sammenlægning og den gradvise neurodegeneration af AD, samt det nylige sammenbrud i lægemiddeludvikling rettet Ap plak, en anden vigtig histologisk markør for AD 36-38, interessen i at opdage lægemidler, der kontrollerer tau sammenlægning er stigende. Faktisk har adskillige grupper allerede begyndt narkotika skærme ved forskellige gennemløb under anvendelse af in vitro tau aggregation reaktioner som den primære assay. Der blev fundet en række kemikalier til at udstille hæmmende eller vending aktiviteter på tau sammenlægning in vitro 39-42. Men alle aktuelle tau sammenlægning regulator skærme bruger umodificeret tau, der misser nøglen patologiske mærke fosforylation, hæve en bekymring for specificiteten og effekten af anvendelse af disse forbindelser i AD behandling.
En af de store forhindringer for at udvikle sammenlægning analyser til biokemisk karakterisering og AD lægemiddelscreening er produktionen af tilstrækkelige mængder af det patofysiologisk relevant hyperphosphoryleret tau protein. Brug af lynlåse assisteret katalyse, i hvilket 1N4R isoform af tau og GSK-3β kinase er co-udtrykkes i E. coli som leucin-zipper fusionsproteiner har vi overvinde denne udfordring (. Sui et al fremlagde, se figur 1 for de endelige produkter af tau og p-tau, se også 43 for foreløbig massespektrometri karakterisering af p-tau). Fra et panel af ni antistoffer specifikke for forskellige phosphoryleringssteder af tau, blev positive signaler ses i otte positioner (data ikke vist). Nedenfor beskriver vi protokoller og besætninger, der kan differentiere sammenlægning kinetiske differences mellem umodificeret tau og p-tau arter. Disse analyser blev ændret fra offentliggjorte protokoller, målte stigningen i fluorescens af Thioflavin T (ThT) eller Thioflavin S (THS) efter amyloid (tau aggregater) bindende 26. I den første "terminal", ikke-dye fremgangsmåde er sammenlægning reaktioner samlet og inkuberet i fravær af amyloid farvestof. På forskellige tidspunkter, er en portion af hver reaktion fjernet og blandet med lige så stort volumen af ThT-holdige buffer at stoppe aggregering og tillade ThT at binde tau aggregater. Fluorescens måles af en IAP FluoroMax-2 fluorometer. I den anden "med-dye" løbende overvågning assay, ThT eller THS er i aggregeringen reaktioner. Fluorescens kan måles løbende under hele forsøget manuelt eller ved hjælp af en multi-pladelæser. Desuden beskriver vi et assay, der anvender en næsten fysiologisk koncentration af tau og p-tau for aggregering i den løbende måling mode. Virkningen af phosphorylering forbliver let påviselig. Nedenfor vil vi beskrive trin-for-trin drift procedurer, og viser repræsentative resultater af disse analyser. Diskussion af nogle af de fordele og ulemper ved hver metode, samt potentielle lægemiddel screening applikationer vil følge.
Ved en høj koncentration, tau aggregater i amyloid-lignende strukturer spontant. Men i laboratoriet, tau fibrillization typisk accelereres ved sådanne inducere heparin (gennemsnitlig molekylvægt 6.000 g / mol) og arachidonsyre. Eksempler er vist heri, indbefatter 30 pM heparin. Dannelsen af tau amyloid aggregater overvåges af fluorescens som følge af amyloid binding ved thioflavin T (ThT) eller thioflavin S (THS). Ved binding til Tau aggregater ThT udviser en rød forskydning i fluorescens (excitation: 450 nm; topemission: 485 nm). THS på den anden side har svagt emission ved 510 nm (excitation ved 450 nm) før amyloid binding, men dette fluorescence øges betydeligt i nærværelse af en amyloidprotein såsom den aggregerede tau 44. Begge farvestoffer fungerer godt i forbindelse med afsløring tau og p-tau sammenlægning. På grund af den stærke og relativt bred emission peak af ThT (se figur 2), er der kun 30% reduktion i fluorescens enhed ved 510 nm. For nemheds skyld bruger vi den samme kombination af excitation / emission bølgelængder (dvs. 450 nm / 510 nm) til at overvåge tau aggregering ved brug af enten farvestof.
Tau aggregation kan ske i nærvær eller fravær af farvestoffet, afhængigt af formålet med analysen og tilgængeligheden af tau-protein. Begge former for reaktioner er vist nedenfor. Derudover viser vi driften af to forskellige instrumenter - en enkelt prøve fluorometer (ISA-SPEX FluoroMax-2) og en multi-plade-læser (SpectraMax M2). Læserne skal kunne tilpasse disse protokoller, der passer til deres specifikke behov og instrument tilgængelighed.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Fremstilling af reagenser
- Forbered aggregering buffer (20 mM Tris, pH 7,4, 100 mM NaCl, 1 mM EDTA). Opbevares ved stuetemperatur, stabilt i flere måneder. Supplement 1 mM dithiothreitol (DTT) før brug.
BEMÆRK: en HEPES-buffer (10 mM HEPES, pH 7,5, 0,1 mM EDTA, 5 mM DTT) producerer også lignende resultater i tau sammenlægning. - Forbered thioflavin T eller thioflavin S stamopløsning (3 mM, opløst i aggregering buffer), og filter med 0,22 um sterilt filterenhed. Opbevares ved -20 ° C i et rør, der er omfattet af aluminiumsfolie, stabilt i flere måneder.
- Heparin stamopløsning Forbered (300 uM, opløst i sammenlægning buffer). Opbevares ved -20 ° C, stabilt i måneder.
- Forbered dithiothreitol (DTT) lager (1 M, opløst i vand). Alikvot i 1,5 ml rør. Opbevares ved -20 ° C. Før sammenlægning assays, tø 1 M opløsning ved stuetemperatur. Fra denne 1.000 x bestand, forberede en portion på 100 mm, der arbejder lager med demineraliseret vand. Efterlad på isindtil den er klar.
- Fjern tau fra -80 ° C fryser. Optøs på is. Juster tau til forudbestemt koncentration med sammenlægning buffer. Spin i en mikrocentrifuge ved 20.800 xg i 10 minutter ved 4 ° C for at fjerne på forhånd dannede store aggregater. Denne pre-spinning trin øger konsistens i den efterfølgende fluorescens måling af hvert parti af proteinet prep. Supernatanten overføres til et andet rør; forlade på is indtil klar til at samle sammenlægning reaktion.
2.-dye, Terminal assay
BEMÆRK: aggregering reaktion af dette assay udføres i fravær af det fluorescerende farvestof. Efter at blande alle komponenterne, reaktionen lov til at forløbe til forudbestemte tidspunkter. Alikvoter tages derefter ud af aggregering reaktionen og blandes med ThT eller THS for amyloid binding før fluorescensaflæsning. Det oprindelige volumen af aggregeringen reaktion afhænger af antallet af tidspunkter nødvendige. Denne fremgangsmåde kan requIRE en stor mængde tau-protein, men er hurtig, enkel og kan udføres i et fluorometer eller en multi-brønds plade-læser (se diskussion). Nedenfor er trin-for-trin, brug ISA SPEX FluoroMax-2 kompakt spektrofluorometer til fluorescens kvantificering.
- Opsætning sammenlægning blandingen i 1,5 ml Eppendorf-rør som i tabel 1. Hver kolonne repræsenterer ingredienser er nødvendige for en 100 pi reaktion, hvilket er nok til én gang-punkt måling. Juster mængden for hele sammenlægning mix baseret på tidspunkter, der er nødvendige for det bestemte forsøg. Tilføj yderligere 10% af hver komponent for at give plads til pipettering fejl. Den typiske reaktion indeholdende heparin vist her kan erstattes af arachidonsyre eller aggregering buffer. Tilføj DTT 1 mM til reaktionsblandingen. Hvis hele reaktionen varer mere end én dag, supplerer frisk DTT hverdag (1 mM) for at sikre et reducerende miljø.
- Vend røret et par gange for at blande. Place hver reaktion i en 37 ° C inkubator eller vandbad. Omrøring er ikke nødvendigt for tau sammenlægning.
- Før måling af fluorescens, tænde spektrofluorometer (lampen først, derefter computer).
BEMÆRK: xenonbuelampe, der kan bruges med det samme. Men for de bedste resultater lade maskinen varme op i ca. 10 min, før du læser fluorescens. - Start softwaren på computeren.
- Vælg Real Time Display mode i Instrument Control Center, skal du indstille ekscitationsbølgelænde til 450 nm (slids til 2 nm) og emission bølgelængde til 510 nm (slids til 5 nm). Luk Real Time Display mode vinduet for at vende tilbage til instrumentet Control Center.
- Vælg Constant bølgelængde Analysis, tryk på Tilføj >> tasten i den øvre ramme for at tilføje bølgelængder sæt. Set erhvervelse parametre Standard fejl til 1. og maksimale Trials til 3, og klik derefter på Tilføj. Klik på Gå! At åbne Data Displayvindue.
- I Data Display, skal du klikke på Start Acq for at åbne dialogboksen Ny Sample kassen. Vælg "ukendt" for prøvetype.
- Til hver sammenlægning blanding 100 pi, tilsættes 98 pi sammenlægning buffer og 2 pi 3 mM Thioflavin T. Pipet flere gange for at blande.
- Overfør hele blandingen til en kuvette (FCA3, udvendig dimension, wxlxh = 12,5 mm x 12,5 mm x 45 mm). Placer kuvetten i prøveholderen i prøven rum og luk låget. Klik på Kør for at indsamle fluorescensdata. Registrere data.
- Tag kuvetten og dekanteres løsningen. Skyl kuvetten ved destilleret vand 3 gange. Tør ved at blæse luft i og uden for kuvetten.
3.-dye, Kontinuerlig tilstand analysen på en SpectraMax M2 Plate Reader
BEMÆRK: Dette assay er forskellig fra den foregående ved, at det fluorescerende farvestof ThT eller THS er inkluderet i aggregation reaktion. Dette muliggør kontinuerlig måling af det samme sæt af reaktioner. På grund af den gentagne brug af reaktionen er denne fremgangsmåde gøres bedre med en automatisk multi-brønds pladelæser (som vist nedenfor driften af SpectraMax M2). En regelmæssig fluorometer fungerer også, men hyppigheden af måling af hurtige sammenlægning reaktioner er noget begrænset på grund af den manuelle karakter af operationen.
- Opsætning sammenlægning mix i en 96-brønds plade (96 brønde sort fast plade, godt volumen 360 pi, flad bund) som i tabel 2. Hver kolonne repræsenterer ingredienserne er nødvendige for en 200 pi reaktion, hvilket er nok til en Tidsforløb måling. Bland godt ved pipettering adskillige gange. Supplement frisk 1 mM DTT hver dag under hele eksperimenter.
- Inkubér 96-brønds plade ved 37 ° C.
- På hvert tidspunkt før fluorescensmåling, tænde for multi-mode mikropladelæser og computeren. Tillad tilstrækkelig tid til machine at stabilisere sig, omkring 10 min.
- Start softwaren på computeren. Indstil temperaturen til 37 ° C, og vælg Fluorescens Intensitet (FI-Top Læs) funktionen, sæt ekscitationsbølgelænde ved 450 nm og emissionsbølgelængde på 510 nm.
- Sæt 96 brønde i skuffen, og tryk på READ for at starte målingen.
- Efter at have læst, fjerne pladen og returnere det tilbage til 37 ° C inkubator. Kopier data og indsætte i et Excel-regneark til dataanalyse og plotning.
4.-dye, Kontinuerlig tilstand analysen på en Compact Spectrofluorometer
- Opsætning sammenlægning blandingen i 1,5 ml Eppendorf-rør som i tabel 3. Hver kolonne repræsenterer ingredienserne er nødvendige for en 200 pi reaktion, hvilket er nok til én gang-punkt måling.
- Vend røret et par gange for at blande.
- Tænd spektrofluorometer og indstil software som i trin 2.3 og 2.4.
- Overfør hele blandingen toa kuvette. Placer kuvetten i prøveholderen i prøven rum og luk låget. Klik på Kør for at indsamle fluorescensdata. Registrere data.
- Fortsæt læsning med passende mellemrum, ved at klikke på Kør og registrering af data. Hvis sammenlægningen skal overvåges ved en høj frekvens (f.eks hver 30 eller 60 sek), lader reaktionen i kuvetten, og i maskinen, indtil enten målingen er færdig, eller når der er tilstrækkelig tid til at bytte reaktioner eller kuvetter.
- Tag kuvetten og dekanteres løsningen. Skyl kuvetten ved destilleret vand 3 gange. Tør ved at blæse luft i og uden for kuvetten.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Brug af rekombinant tau og p-tau (figur 1), etablerede vi to forskellige protokoller til sammenligning kinetikken for sammenlægning af tau og p-tau, at drage fordel af den stærke fluorescensemission af ThT og THS efter binding til amyloidal proteinaggregater, herunder tau og p-tau (figur 2). Med eller uden det fluorescerende farvestof i aggregeringen reaktion observerede vi konsekvent forøgelse af tau aggregation af hyperphosphorylering (figur 3-5). Denne stimulering er uafhængig af heparin (data ikke vist). I en typisk reaktion, tau og p-tau oligomerisere ved høje rater inden for de første 30 minutter før bremse betydeligt (figur 3 og 5), med p-tau udviser højere fluorescensenheder under hele forsøget. Herunder ThT i aggregeringen reaktioner forårsager betydelig forsinkelse i hastigheden for sammenlægning (figur 4). Begge isoformer nærmede plateau 160 timer efter reaktionerne var begyndt. THS, på den anden side, ikke forårsager mærkbar afmatning om sammenlægning (figur 5).

Figur 1. Oprenset tau og hyperphosphoryleret tau (p-tau), der anvendes i denne undersøgelse. Prøver blev løst ved en 10% SDS-PAGE-gel og farvet med Coomassie blue R250 (venstre) eller probes med et anti-tau monoklonalt antistof (højre panel). Lane M, molekylvægtmarkør; bane 1 og 3, ikke-phosphoryleret tau; bane 2 og 4, hyperphosphoryleret tau.

Figur 2. Emission spektre for ThT (30 uM) med eller uden binding tau aggregater. Emission overtagelse blev s dåse fra 460 nm til 600 nm (1 nm tilvækst, 0,1 sek integration, 5 nm slids bredde), excitation ved 450 nm. Tau aggregater blev opnået ved at lade 50 uM tau sammenlægning forløbe ved 37 ° CO / N (se protokol 2 for detaljer).
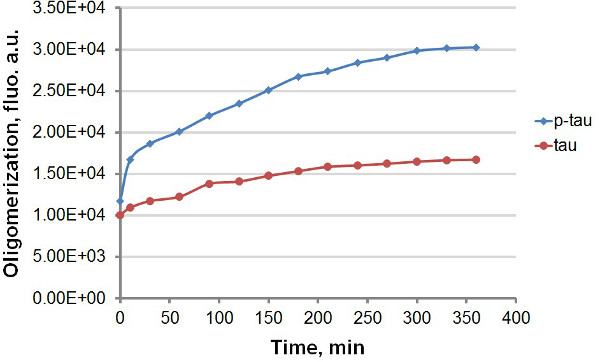
Figur 3. sammenlægning kurver for tau og p-tau i terminal assay. Sammenlægning af 50 uM tau og p-tau blev afsluttet med 30 pM heparin som inducer. På forskellige tidspunkter efter begyndelsen af reaktionen blev 100 pi af reaktionen fjernet og blandet med samme volumen af 60 uM ThT før fluorescensmåling. Fluorescens blev målt ved 450 nm excitation, 510 nm emission. "au", vilkårlige enheder. Bemærk, at tidsskala er på få minutter.
"Figur 4" src = "/ files / ftp_upload / 51537 / 51537fig4highres.jpg" />
Figur 4. sammenlægning kurver for ThT alene tau og p-tau i den løbende måling tilstand i nærvær af ThT. Hver reaktion bestod af 0 eller 50 uM tau eller p-tau, 30 pM heparin, og 30 uM ThT kollektivt buffer . Reaktioner blev inkuberet ved 37 ° C i en 96-brønds plade. På forskellige tidspunkter blev pladen fjernet fra inkubatoren og indlæst til pladelæser til fluorescens aflæsning (excitation 450 nm, emission 510 nm). Mellem aflæsninger blev pladen holdes i inkubatoren uden omrøring under et dæksel. Tilstedeværelsen af ThT betydeligt langsommere sammenlægning, men, vigtigere, hyperphosphoryleret tau stadig udstillet hurtigere aggregeringsniveau end gjorde sit umodificeret modstykke. Bemærk, at omfanget af tid i timer.
pload / 51537 / 51537fig5highres.jpg "/>
Figur 5. Mindre tau sammenlægning assays i nærværelse af ths. In vitro heparin-induceret aggregering af 6 uM tau og p-tau blev vurderet i den stadige måling mode med Thioflavin S som indikator farvestof. Ud over protein, hver reaktion indeholdt 30 pM heparin og 20 uM THS i HEPES sammenlægning buffer (10 mM HEPES, pH 7,5, 5 mM DTT, 0,1 mM EDTA). Alle ingredienser undtagen heparin blev blandet og bragt i ligevægt ved stuetemperatur. Efter tilsætning af heparin blev reaktionen overført til kuvetten og anbringes i prøveholderen. Fluorescens blev registreret straks som T 0 og fortsatte i ca. 2 timer eller indtil stigningen fluorescens bremset ned til nær nul. På grund af den relativt korte reaktionstid blev hele reaktionen udføres ved stuetemperatur i samme kuvette.
| tau | p-tau | |
| 60-100 uM tau | 50 pi | 0 pi |
| 60-100 pM p-tau | 0 pi | 50 pi |
| 300 pM heparin | 10 pi | 10 pi |
| sammenlægning buffer | 39 pi | 39 pi |
| 100 mM DTT | 1 pi | 1 pi |
Tabel 1. Sammenlægning blandingen komponenter til ikke-dye, terminal assay.
| tau | p-tau | Dye alene | |
| 60-100 uM tau | 50 pi | 0 pi | 0 pi |
| 60-100 pM p-tau | 0 pi | 50 pi | |
| 300 pM heparin | 20 pi | 20 pi | 20 pi |
| 3 mM thioflavin T- | 2 pi | 2 pi | 2 pi |
| Sammenlægning buffer | 126 pi | 126 pi | 176 pi |
| 100 mM DTT | 2 pi | 2 pi | 2 pi |
Tabel 2. Sammenlægning blandingen komponenter til med-dye, løbende analyse på en plade-læser.
| tau | p-tau | Dye alene | |
| 60-100 uM tau | 5 pi | 0 pi | 0 pi |
| 60-100 pM p-tau | 0 pi | 5 pi | 01 l |
| 300 pM heparin | 20 pi | 20 pi | 20 pi |
| 3 mM thioflavin S | 1,5 pi | 1,5 pi | 1,5 pi |
| Sammenlægning buffer | 171,5 pi | 171,5 pi | 176,5 pi |
| 100 mM DTT | 2 pi | 2 pi | 2 pi |
Tabel 3. Sammenlægning blandingen komponenter til med-dye, løbende analyse på et kompakt spektrofluorometer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Denne protokol demonstrerer forskellige assaybetingelser og instrumenter, der registrerer de phosphoryleringsafhængige hurtigt tau sammenlægning kinetik. I terminalen assay fluorescens farvestof ThT tilsættes til en portion af fjernet fra master mix på hvert tidspunkt reaktion. Amyloid binding-induceret fluorescens måles derefter 26. I den anden, med farvestof tilstand, tau aggregation udføres i nærvær af ThT eller THS, hvilket gør denne type reaktion er egnet til tidstro automatisk vurdering af vækst af tau-aggregater. Hver af disse metoder har sine fordele og ulemper.
Terminalen-mode reaktion udføres med kun de ingredienser, der er nødvendige for tau sammenlægning. Fortynding og blanding af reaktionsblandingen med thioflavin T-drastisk sinker hastigheden af fluorescens stigning væsentlige standser reaktionen til fluorescens kvantificering. Denne fremgangsmåde er således også kompatibel med manuel betjening. Men fordi reaction er praktisk opsiges ThT Desuden kan der kræves en stor mængde tau for at planlægge en sammenlægning kurve. En anden potentiel advarsel for denne metode er, at hyppig adgang til reaktionsblandingen kan indføre mikrobiel eller proteolytisk kontaminering eller protein oxidation. I modsætning hertil med farvestof tilstand tillader dannelsen af amyloid i nærvær af ThT eller ths. Udviklingen af sammenlægning kan overvåges kontinuerligt uden nogensinde at forstyrre reaktionen. Denne funktion er særligt attraktivt, når oprettelse af en automatiseret assay platform. Dog kan forskellige farvestoffer fremkalde specifikke svar. Faktisk ThT forsinker tau og p-tau aggregation betydeligt, men THS har lidt virkninger (sammenlign figur 3 og 5). Der er flere andre fluorescensfarvestoffer, herunder congorødt og thiaziner, der er blevet anvendt i histologiske og cellebiologi undersøgelser for PHF-dannelse. Mindst en rapport, at nogle af disse farvestoffer kan fremkalde tau sammenlægning i vævdyrkede celler 45. Derfor, når du vælger disse forbindelser til amyloidogenese kinetikundersøgelser, forsigtighed skal praktiseres, og at forskellige farvestoffer måske skal sammenlignes.
Med hensyn til valg af instrument, single-prøve fluorometer anvendt i den første tilgang er yderst pålidelig, men operationen kan være besværlig, når mere end et par reaktioner skal sammenlignes. Brug af flere kuvetter kan hjælpe med at undgå krydskontaminering mellem reaktioner, selv om udgifterne til disse skrøbelige kvartskuvetter kan være uoverkommelige for nogle. Omvendt kan multi-brønds mikropladelæser undersøge flere reaktioner på samme tid. Anvendelsen af engangs plader med 96 brønde er fordelagtig samt. Med et varmeelement kan en mikropladelæser være en dedikeret indretning til overvågning af flere reaktioner ved 37 ° C i en længere periode. Dog kan fordampningen være en bekymring. DiNitto et al. Overlejret af en reaktion lig med mineralsk olie for at forhindrefordampning 46.
Nogle forholdsregler skal tages for ovennævnte protokoller for at sikre ensartede og kvantitative resultater. For det første tau og p-tau spontant danne amyloide aggregater over tid, især når en høj koncentration. Det er derfor bydende nødvendigt at indefryse alle de portioner af protein preps, og tø kun den nødvendige mængde før forsøgene. Alligevel kan nogle aggregater, herunder granulerede mellemprodukter påvises ved ThT 47 har dannet under udarbejdelsen af rekombinante proteiner. En væsentlig indledende fluorescens aflæsning af en typisk sammenlægning reaktion er således almindelig. Ikke desto mindre, tilsætning af et præ-spinding trin og overføre supernatanten til et separat rør, selv uden en synlig protein pellet, kan reducere og vedligeholde ensartet indledende fluorescens af samme batch af tau og p-tau prep. For det andet arbejder stamopløsning af ThT (dvs. 60 uM) er stabil ved stuetemperatur i mindre end en uge før the fluorescens mindskes. Det er således rådes til at re-make 60 uM ThT hver flere dage.
En fremtrædende årsag underliggende studier af p-tau sammenlægning er udvikling af nye AD diagnostik og terapi. Forbindelser, der hæmmer eller vende tilbage rekombinant tau sammenlægning er blevet identificeret fra high-throughput skærme og målrettede test 18,40,41,48. Effekten af disse forbindelser til p-tau aggregation stadig at blive belyst. Disse skærme blev udført i terminalen tilstand ved afgivelse af en fælles sammenlægning mix uden farvestoffet til individuelle flerpladet brønde med forskellige forbindelser. Typisk efter O / N inkubering ThT eller THS tilføjet for fluorescensmåling, afslører den inhiberende effekt af mange forbindelser. Den med-dye fremgangsmåde nævnt ovenfor, og af Rankin et al. 49 endnu ikke er indarbejdet high-throughput skærme. Nu med hyperphosphoryleret tau rådighed for kinetiske og farmaceutiske undersøgelser af parretspiralformede filamenter dannelse, Alzheimers sygdom lægemiddelforskning er sandsynligt at fremme længere.
Endelig er det værd at bemærke, at studiet af p-tau sammenlægning er af afgørende betydning for ikke blot tauopatier, men kan også påvirke en endnu bredere befolkning. For eksempel er der rapporter om, at de neurofibrillære tråde kan påvises hos nogle patienter med kronisk traumatisk encefalopati, såsom professionelle amerikanske fodboldspillere og boksere 50-52. Lignende korrelation er også rapporteret for patienter enkelt eller gentagne traumatisk hjerneskade, herunder soldater 53. Protokoller, der er beskrevet i dette arbejde kan således hjælpe opdagelse og udvikling af nye lægemidler rettet mod p-tau aggregater i nerveceller.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q.
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S.
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M.
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B.
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
