

Summary
非修飾および高リン酸化タウタンパク質は、高リン酸化依存速い凝集速度論を明らかに二つのインビトロ凝集アッセイに使用した。これらのアッセイは、アルツハイマー病の進行の基礎となる原線維を形成する過リン酸化タウの性向を調節することができる化合物のための将来のスクリーンのための道を開く。
Introduction
アルツハイマー病(AD)は、タウオパチーとして知られる神経変性疾患の大規模なコレクションの一つである。典型的な病理学の基礎となるタウオパチーは、ニューロン、アストロサイトとミクログリア1-4で神経原線維変化、NFTの、です。 NFT密度は、認知機能障害3,5及びニューロン損失6と相関する。 NFTはまっすぐまたは対らせんフィラメント(PHF)7,8を形成(以下、「P-タウ」とも呼ばれる)は、主に過剰リン酸化タウタンパク質が含まれています。タウは、ニューロンのシグナル伝達および輸送9,10に必須である軸索輸送を促進すると考えられ、微小管関連タンパク質である。各タウ分子は、正常な脳で2〜3リン酸を含むが、タウオパチー患者11内の複数の折り目によってホスホリル含有量が増加する。複数のキナーゼは、GSK3β(グリコーゲン合成酵素キナーゼ3β)及びCDK5(サイクリン·ドなどのタウの過剰リン酸化に貢献する可能性があるペンダントキナーゼ5)12,13が、病理学的リン酸化のための直接の引き金は、とらえどころのない14のまま。微小管結合モチーフまたは周辺の異常なリン酸化は、微小管15からタウを解離し、p-タウは、最終的にNFT介在物に重合させることができる直鎖または対らせんフィラメントにオリゴマー化する細胞体コンパートメントにタウ誤局在を引き起こす。タウの過剰リン酸化、NFT形成、及び神経変性の間の緊密な提携は、p-タウのもつれがアポトーシスおよび他の細胞毒性反応を誘発するため、タウオパチーの神経変性16,17のための根本的な原因であることが流行しているという仮説につながった。この前提に基づく薬物スクリーニングと初期の臨床試験は、18を立ち上げてきた。しかし、この仮説は19,20挑戦に直面している。例えば、サンタクルスらは、トランスジェニックマウスの認知機能は、変異体の発現を抑制することにより改善することができることを示したのNFTは、既存のタウ分子21から形成するために続けていても、人間のタウ、。 ショウジョウバエモデルでは、NFTは、基礎となる神経細胞22,23を保護するために有毒な細胞質ゾルタウを隔離することが示された。明らかに、NFTの病因の役割は、もしあれば、大いにタウオパチー治療薬の開発の方向性に影響を与えます。
いくつかのβシート好ましい蛍光色素、電子顕微鏡法、光散乱分光法24-27の結合によって示されるように、高濃度では、組換えまたは正常な脳のタウタンパク質は自然に、ゆっくり、 インビトロでの PHF様構造へと重合する。追加ヘパリンまたはアラキドン酸、人間の脳に豊富脂肪酸は、大幅にタウisoform-と誘導物質の濃度依存マナー28-32でPHFの形成を促進する。興味深いことに、過リン酸化タウは、AD脳から精製またはインビトロリン酸化反応A に網羅して調製より速くggregatesと、より効率的に26,33-35。これらの結果は、p-タウの病理学的役割とよく一致している。 p-タウの凝集に基づくインビトロシステムは、このようにADの薬物スクリーニングのための強力なツールとして機能することができる。
タウ凝集およびADの進行神経変性、ならびにAβ斑をターゲットに医薬品開発の最近の失敗、AD 36-38のもう一つの重要な組織学的マーカーの間に密接な関連性を考えると、タウ凝集を制御する薬を発見するの関心が高まっている。実際、いくつかのグループがすでに一次アッセイとして体外タウ凝集反応に使用して、異なるスループットで薬物スクリーニングを開始しました。多くの化学物質は、 インビトロ 39-42 でタウ凝集阻害または逆転活性を示すことが見出された。しかし、現在のすべてのタウ凝集レギュレータスクリーンは蛍光体の主要な病理学的なマークをミス修正されていないタウを使用ylation、ADの治療におけるこれらの化合物を使用しての特異性および有効性の懸念を高める。
生化学的特性およびADの薬物スクリーニングのための凝集アッセイの開発の大きな障害の一つは、病態生理学的に関連した高リン酸化タウタンパク質の十分な量の産生である。タウの1N4RアイソフォームとGSK-3βキナーゼはEに共発現されているジッパーアシスト触媒反応システムを用いて、 大腸菌は、ロイシンジッパー融合タンパク質として、私たちは、この課題を克服している(。隋ら 、提出され、タウとp-タウの最終製品については、図1を参照してください。また、p-タウの予備質量分析の特性評価のための43を参照)。タウの異なるリン酸化部位に特異的な9抗体のパネルから、陽性シグナルは、8つの位置で見られた(データは示さず)。以下では、集約運動Dを区別できるプロトコルおよびインストルメンテーションを記述する変更されていないタウとpタウ種間ifferences。これらのアッセイは、26アミロイド結合の際に、チオフラビンT(THT)またはチオフラビンS(THS)の蛍光の増加(タウ凝集体)を測定した公開されたプロトコルから変更された。最初の「端末」は、無色素のアプローチは、凝集反応が組み立てられ、アミロイド色素の非存在下でインキュベートした。異なる時点で、各反応のアリコートを除去し、タウ凝集体を結合するためのThTの凝集を停止し、可能にするためのThT含有緩衝液の等量。蛍光は、IAP FluoroMax-2蛍光光度計で測定される。第二」の色素「継続的なモニタリングアッセイでは、ThTのまたはTHSは凝集反応に含まれる。蛍光は、手動で、実験全体を通じて連続的に測定またはマルチプレートリーダーを使用することができる。加えて、我々は継続的な測定MOにおける集計のためのタウおよびp-タウの近生理的濃度を使用するアッセイを説明デ。リン酸化の効果は容易に検出可能のままである。以下では、ステップバイステップの操作手順を説明し、これらのアッセイの代表的な結果が表示されます。各アプローチの長所と短所のいくつかの議論だけでなく、潜在的な薬剤スクリーニングアプリケーションが続きます。
高濃度では、タウは自然にアミロイド様構造へと集約されます。しかし、実験室で、タウ線維化は、典型的には、ヘパリン(平均分子量6,000グラム/モル)及びアラキドン酸のような誘導因子によって促進される。ここに示した例には、30μMのヘパリンが含まれています。タウアミロイド凝集体の形成を、チオフラビンT(ThTの)またはチオフラビンS(THS)のアミロイド結合性から生じる蛍光によりモニターされる。 (;:485 nmの発光ピークを450nm励起)タウ凝集体に結合すると、ThTの蛍光の赤いシフトを示す。 THSは、一方では、アミロイドは結合の前に510ナノメートル(450nmで励起)で弱い発光を有するが、このfluorescenc電子は、このような凝集したタウ44とアミロイドタンパク質の存在下で有意に増加する。両方の色素は、タウおよびp-タウ凝集を検出するのに適しています。ためのThTの強力かつ比較的広い発光ピーク( 図2参照)から、510 nmでの蛍光単位でわずか30%の減少がある。いずれかの染料を使用する際の便宜のために、我々は、タウ凝集をモニターするために、励起/発光波長( すなわち 、450nmの/ 510 nm)の同じ組み合わせを使用する。
タウ凝集は、アッセイの目的とタウタンパク質の可用性に応じて、染料の存在下または非存在下で行うことができる。反応の両方のモードを以下に示します。単一のサンプル蛍光光度計(ISA-SPEX FluoroMax-2)と、マルチプレートリーダー(スペクトラマックスM2) - 加えて、我々は2つの異なる機器の動作を実証する。読者は彼らの特定のニーズや機器の可用性に合わせて、これらのプロトコルを適応させることができるはずです。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
試薬の調製
- 集計バッファを準備し(20mMトリス、pH7.4中、100mMのNaCl、1mMのEDTA)。ヶ月間、室温で安定して保管してください。サプリメント1mMのは、使用前に(DTT)をジチオスレイトール。
NOTE:HEPESベースの緩衝液(10mM HEPES、pH7.5で、0.1mMのEDTA、5mMのDTT)もまた、タウ凝集に類似の結果を生成する。 - 0.22μmの無菌フィルターユニットによりチオフラビンTまたはチオフラビンS原液(凝集緩衝液に溶解3mMの、)、およびフィルタを準備します。ヶ月間、アルミホイルで安定したカバーチューブで-20℃で保存。
- (300μM、集計緩衝液に溶解)をヘパリン原液を準備します。 -20°Cで保存し、ヶ月間安定。
- (水に溶解し、1M、)ジチオスレイトール(DTT)の株式を準備します。 1.5ミリリットルチューブに小分けし。 -20℃で保管してください。凝集アッセイの前に、室温で1 M溶液を解凍する。この1,000倍在庫から、脱イオン水で100 mMの作業ストックの一定分量を準備。氷の上のままに直前まで。
- ℃の冷凍庫-80℃からタウを削除します。氷上で解凍します。集約緩衝液で所定濃度にタウを調整します。予め形成された大きな凝集体を除去し、4℃で10分間20800×gでマイクロ遠心スピン。このプレ紡糸工程は、タンパク質調製物の各バッチの後続の蛍光測定の一貫性を増大させる。別のチューブに上清を移し、凝集反応を組み立てるために準備ができるまで氷上に残す。
2.無色素、ターミナルアッセイ
注:このアッセイの凝集反応は、蛍光色素の不存在下で行われる。すべての成分を混合した後、反応物を所定の時間点まで進行させる。アリコートを凝集反応から取り出しのThTまたは蛍光読み取りの前に結合アミロイドTHSと混合される。凝集反応の初期体積は、必要とされる時点の数に依存する。このアプローチは、のrequありタウタンパク質を大量にIREが簡単で高速であり、蛍光光度計またはマルチウェルプレートリーダー(説明を参照)で行うことができる。以下は、蛍光定量化のためのISA SPEX FluoroMax-2コンパクトな分光蛍光光度計を使用して、ステップバイステップの操作です。
- 表1のように1.5mlのエッペンドルフチューブ中で凝集物を設定し、各列は、一時点の測定のために十分である、100μlの反応に必要な成分を表す。特定の実験のために必要な時点に基づいて、全体の集計ミックス量を調整します。ピペット操作エラーの余地を与えるために、各コンポーネントの追加の10パーセントを追加します。ここに示されている典型的な反応を含むヘパリンはアラキドン酸または集約バッファで置き換えることができる。反応混合物を1 mMのDTTを加える。全反応が1日以上続いた場合、還元環境を確保するために新鮮なDTT、毎日(1 mm)を補う。
- 混合するチューブを数回転倒。プラ37℃のインキュベーターや水浴中CEの各反応。攪拌はタウ凝集のために必要とされていません。
- 蛍光を測定する前に、(コンピュータその後、第1ランプ)分光蛍光をオンにします。
注:すぐに使用することができ、キセノンアークランプ。しかし、最良の結果を得るためにマシンが蛍光を読む前に約10分間のウォームアップすることができます。 - コンピュータ上のソフトウェアを起動します。
- 510 nmの(5 nmのスリット)に計測器制御センターの実時間表示モード 、(2 nmのスリット)450nmの設定励起波長と発光波長を選択してください。閉じるリアルタイム表示モードウィンドウは計測器制御センターに戻ります。
- 一定の波長の分析を選択して、Enterキーを押して波長セットを追加するには、上部フレームで>>キーを追加します。 1〜3と最大トライアルへの標準誤差の集合取得パラメータ、[ 追加]をクリックします。 データ表示を開くために!GOをクリックします。窓。
- データ表示 ] ウィンドウで、 新しいサンプル]ダイアログボックスを開くには[スタート] ACQをクリックします。 サンプルタイプのために「不明」を選択します。
- すべての100μlの集約混合物に、98μlの集計バッファとT.ピペットを数回混合する3 mMのチオフラビン2μlを添加する。
- キュベット(FCA3、外装寸法は、wxlxh = 12.5ミリメートルX 12.5ミリメートルX 45ミリメートル)の混合物全体を転送します。サンプル·コンパートメント内の試料ホルダーにキュベットを置き、蓋を閉じます。蛍光データを収集するために実行]をクリックします。データを記録します。
- キュベットを取り出し、その溶液をデカント。蒸留水により3回キュベットをすすぐ。キュベット内と外の空気を吹き付けることにより乾燥した。
と色素3.をSpectraMax M2プレートリーダーで継続モードアッセイ
注:このアッセイは、蛍光色素のThTまたはTHSがaggregaに含まれているという点で、以前のものとは異なるション反応。これは、反応の同じセットの連続測定を可能にする。なぜなら反応の反復使用のために、この方法は、より良好な自動マルチウェルプレートリーダー(スペクトラマックスM2の動作を次のように)で行われる。定期的な蛍光光度計も動作しますが、高速の集約反応の測定の頻度が原因の動作の説明書性質のため、多少制限されている。
- 表2のように96ウェルプレート中で凝集ミックスを設定する(96ウェル黒色固体プレート、ウェル容量360μlを、平底)の各列は、一のために十分である、200μlの反応に必要な成分を表す時間点測定。数回ピペッティングしてよく混ぜる。実験の過程を通して、毎日新鮮な1mMのDTTを補足する。
- 37℃で96ウェルプレートをインキュベートする。
- 蛍光測定の前に、各時点で、マルチモードマイクロプレートリーダー及びコンピュータの電源を入れる。 MAのための十分な時間を許可する背骨は、約10分間安定させる。
- コンピュータ上のソフトウェアを起動します。 37℃に温度を設定し、蛍光強度(FI-トップリード)モード、510 nmで450 nmおよび放出波長に設定励起波長を選択します。
- 引き出しに96ウェルプレートを挿入し、測定を開始するために、READキーを押します。
- お読みになった後、プレートを取り外して、37℃のインキュベーターに戻し、それを返す。データ分析とプロットするためのExcelスプレッドシートにデータをコピーして貼り付け。
4と染料、継続モードアッセイコンパクト分光蛍光光度計で
- 表3のように1.5mlのエッペンドルフチューブ中で凝集物を設定し、各列は、一時点の測定のために十分である、200μlの反応に必要な成分を表す。
- 混合するチューブを数回転倒。
- 分光蛍光光度計の電源を入れ、手順2.3と2.4のようにソフトウェアを設定する。
- 混合物全体tを転送OAキュベット。サンプル·コンパートメント内の試料ホルダーにキュベットを置き、蓋を閉じます。蛍光データを収集するために実行]をクリックします。データを記録します。
- 実行]をクリックし、データを記録することにより、適切な間隔で読み続ける。凝集は、高周波数( 例えば 、毎30または60秒)で監視する場合には、いずれかの測定が終了するまで機械キュベットおよび反応のまま、または反応またはキュベットを交換するのに十分な時間がある場合。
- キュベットを取り出し、その溶液をデカント。蒸留水により3回キュベットをすすぐ。キュベット内と外の空気を吹き付けることにより乾燥した。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
組換えtauおよびp-タウ( 図1)を使用して、我々は、タウを含む、タンパク質凝集体をアミロイドへの結合の際のThTとTHSの強い蛍光発光を利用して、タウの凝集およびp-タウの動態を比較するために2つの異なるプロトコルを設立及びp-タウ( 図2)。凝集反応における蛍光色素の有無にかかわらず、我々は過剰リン酸化( 図3-5)でタウ凝集の一貫した強化を観察した。この刺激は、ヘパリンとは無関係である(データは示さず)。 ( 図3、図5)、p-タウは、実験の過程を通して、より高い蛍光単位を示すと大幅に減速する前に、最初の30分以内に速い速度での典型的な反応、タウおよびp-タウオリゴマー化で。凝集反応でのThTを含めると凝集速度( 図4)の有意な位相差を生じさせる。両方のアイソフォームに近づいPLAT反応が開始した後に160時間をEAU。 THSは、他の一方で、アグリゲーション( 図5)のかなりの減速を引き起こすことはありません。

試料を10%SDS-PAGEゲルによって解決され、クマシーブルーR250(左)で染色するか、または(右抗タウモノクローナル抗体によってプローブした。この研究で使用し、図1タウおよび過リン酸化タウ(p-タウ)精製パネル)。レーンMは分子量マーカー。レーン1および3、非リン酸化タウ。レーン2および4、過リン酸化タウ。

またはタウ凝集体を結合することなくのThT(30μM)図2.発光スペクトル。排出取得したS 、450nmでの励起、600nmの(0.1秒統合、5 nmのスリット幅1 nmの増分)に460nmでの缶詰。タウ凝集は、50μMのタウ凝集を37℃CO / N(詳細については、 プロトコール2を参照)で進行させることによって得た。
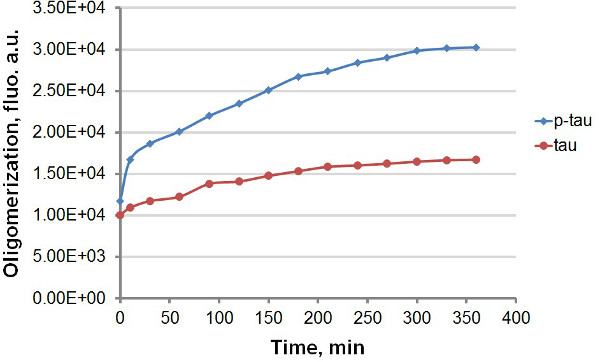
タウと端末アッセイでp-タウ図3.集計曲線が。50μMのタウ及びp-タウの凝集は、誘導物質として、30μMのヘパリンで完成しました。反応開始後の異なる時点で、反応液100μlを除去し、蛍光測定の前に60μMのThTの同容量と混合。蛍光は450nmで励起、510nmの発光で測定した。 「オ」、任意の単位。時間スケールは数分であることに注意してください。
「図4「SRC = "/ファイル/ ftp_upload / 51537 / 51537fig4highres.jpg" />
一人でのThT図4.集約曲線は、ThTの存在下での継続的な測定モードでのタウとp-tauが。各反応は0または50μMのタウまたはp-タウ、30μMヘパリンからなり、集計バッファ内の30μMのThT 。反応は、96ウェルプレート中で37℃でインキュベートした。異なる時点で、プレートをインキュベーターから取り出し、そして蛍光を読み取る(励起450nmで、発光510 nm)のためのプレートリーダーにロード。測定値の間、プレートは、カバーの下に攪拌することなくインキュベータ中に保持した。のThTの存在が大幅に変更されていない対応をしたより重要なのは、過リン酸化タウはまだ集計のより速い速度を示し、凝集を鈍化、しかし。時間のスケールは時間であることに注意してください。
PLOAD / 51537 / 51537fig5highres.jpg "/>
6μMタウおよびp-タウのTHSの存在下で、図5小規模タウ凝集アッセイ。in vitroでのヘパリン誘発凝集を指示色素としてチオフラビンSとの継続的な測定モードで評価した。タンパク質に加えて、各反応は30μMヘパリンおよびHEPES凝集緩衝液(10mM HEPES pH7.5,5mMのDTT、0.1mMのEDTA)中で20μMTHSを含んでいた。ヘパリンを除くすべての成分を混合し、室温で平衡化した。ヘパリンを添加した後、反応キュベットに移し、サンプルホルダーに入れた。蛍光はT 0とすぐに記録され、約2時間、または蛍光増加がゼロ近くに遅くまで続けた。なぜなら、比較的短い反応時間で、全体の反応は同一のキュベット中で、室温で実施した。
| タウ | p-タウ | |
| 60から100μMタウ | 50μL | 0μL |
| 60から100μMのp-タウ | 0μL | 50μL |
| 300μMヘパリン | 10μL | 10μL |
| 集計バッファ | 39μL | 39μL |
| 100 mMのDTT | 1μL | 1μL |
無染料表1.集計混合物成分、端子アッセイ。
| タウ | p-タウ | 一人で染める | |
| 60から100μMタウ | 50μL | 0μL | 0μL |
| 60から100μMのp-タウ | 0μL | 50μL | |
| 300μMヘパリン | 20μL | 20μL | 20μL |
| Tチオフラビン3 mMの | 2μL | 2μL | 2μL |
| 集計バッファ | 126μL | 126μL | 176μL |
| 100 mMのDTT | 2μL | 2μL | 2μL |
と染料表2.集計混合物成分、プレートリーダー上で継続的アッセイ。
| タウ | p-タウ | 一人で染める | |
| 60から100μMタウ | 5μL | 0μL | 0μL |
| 60から100μMのp-タウ | 0μL | 5μL | 01; L |
| 300μMヘパリン | 20μL | 20μL | 20μL |
| チオフラビンS 3 mMの | 1.5μL | 1.5μL | 1.5μL |
| 集計バッファ | 171.5μL | 171.5μL | 176.5μL |
| 100 mMのDTT | 2μL | 2μL | 2μL |
と染料表3.集計混合物成分、コンパクト蛍光光度計での継続的なアッセイ。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
このプロトコルは、リン酸化依存速いタウ凝集速度論を検出する別のアッセイ条件と楽器を示しています。端末アッセイでは、蛍光色素ThTを、各時点でのマスターミックスから除去された反応の一部に加えられる。アミロイド結合誘起蛍光次に26で測定される。第有する染料モードでは、タウ凝集は、タウ凝集体の成長のリアルタイム自動評価に適したこの種の反応をレンダリングのThTまたはTHSの存在下で行われる。これらの各メソッドには、長所と短所を持っています。
ターミナルモードの反応がタウ凝集のために必要な唯一のこれらの成分を用いて行われる。希釈し、チオフラビンTとの反応を混合して大幅に本質的に蛍光定量反応を停止させ、蛍光の増加速度を遅くする。従って、この方法は、また手動操作と互換性があります。しかし、反作用理由nは事実上のThT添加により終了し、タウ多量の凝集曲線をプロットするために必要とされ得る。この方法のための別の潜在的な注意点は、反応混合物への頻繁なアクセスは、微生物またはタンパク質分解汚染、またはタンパク質の酸化を導入することができるということである。対照的に、と染料モードのThTまたはTHSの存在下でのアミロイドの生成を可能にする。凝集の進歩は、これまでの反応を妨害することなく連続的に監視することができる。自動化されたアッセイプラットフォームを確立する場合、この機能は特に魅力的である。しかし、異なる色素は、特定の応答を誘発し得る。実際、ThTを大幅にタウおよびp-タウ凝集を遅らせるが、THSは少しの効果( 図3と図5を比較する)を有している。 PHF形成のために組織学的および細胞生物学の研究において使用されてきたコンゴレッド及びチアジン、を含むいくつかの他の蛍光色素が存在する。少なくとも一つの報告書は、これらの染料のいくつかは、組織におけるタウ凝集を誘導することができると述べている培養細胞45。アミロイド動態研究のためのこれらの化合物を選択する際にそのため、注意が実施されなければならず、異なる染料を比較する必要があること。
器具の選択に関して、第一のアプローチで使用される単一のサンプル蛍光光度計は、非常に信頼性があるが、複数の少数の反応を比較する場合に操作が面倒であってもよい。これらの脆弱な石英キュベットのコストはいくつかの法外かもしれないが、複数のキュベットを使用して、反応の間の交差汚染を避けるのを助けることができる。逆に、マルチウェルマイクロプレートリーダーは、同時に複数の反応を調べることができる。使い捨ての96ウェルプレートを使用することも同様に有利である。加熱要素を、マイクロプレートリーダーは、長期間、37℃で、複数の反応を監視するための専用の装置とすることができる。しかし、蒸発が問題になることがあります。 DiNitto ら防ぐためにミネラルオイルと同様の反応を重ね蒸発46。
いくつかの注意事項は、一貫性の定量的な結果を保証するために、上記のプロトコルのために取るべきである。まず、タウおよびp-tauが自発的に場合は特に高濃度で、時間をかけてアミロイド凝集体を形成する。それは、タンパク質調製物のすべてのアリコートを凍結し、そして実験の前にのみ必要なボリュームを解凍することが不可欠です。そうであっても、ThTの47で検出可能な粒状の中間体を含むいくつかの凝集体は、組換えタンパク質の製造の間に形成された可能性があります。典型的な凝集反応のかなりの初期蛍光読み取りは、このように一般的です。それにもかかわらず、プレスピニング工程を追加しても、目に見えるタンパク質ペレットことなく、別のチューブに上清を移す、軽減、およびタウおよびp-タウプレップ同じバッチの一貫性、初期蛍光を維持することができる。第二に、のThTの作業ストック溶液( すなわち 、60μm)は目の前の週未満の室温で安定しているEの蛍光が減少する。それは、このように60μMのThT数日ごとに再確認することをお勧めします。
p-タウ凝集の研究の基礎となる一つの著名な理由は、新規ADの診断と治療法の開発である。組換えタウ凝集を阻害するか、または戻す化合物は、高スループットスクリーニング及び標的テスト18,40,41,48から同定されている。 P-タウ凝集のためのこれらの化合物の有効性はまだ解明されていない。これらの画面は、異なる化合物との個々のマルチプレートのウェルに染料なしに共通の凝集ミックスを分配することによって、端末モードで行った。典型的には、O / Nインキュベーション後のThTまたはTHSは、多くの化合物の阻害力を明らかにし、蛍光測定のために添加される。と染料のアプローチは、上方及びランキンらにより言及49がまだハイスループットスクリーニングに組み込まれる。今、ペアの運動や医薬品の研究のための利用可能な過リン酸化タウとらせん状フィラメントの形成は、アルツハイマー病の創薬は遠く前進する可能性がある。
最後に、p-タウ凝集の研究だけでなく、タウオパチーに非常に重要であるだけでなく、より広い人口に影響を与えることができることは注目に値する。たとえば、神経原線維変化は、そのようなプロのアメリカンフットボール選手とボクサー50〜52として、慢性外傷性脳症、一部の患者で検出可能であるという報告があります。同様な相関関係はまた、兵士53を含む単一または反復外 傷性脳損傷患者について報告されている。この作品に記載されているプロトコルは、このように神経細胞におけるp-タウ凝集体をターゲットに新たな治療法の発見と開発を助けるかもしれない。
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q. Neurodegenerative tauopathies. Annu Rev Neurosci. 24, 1121-1159 (2001).
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S. Cytoskeletal pathologies of Alzheimer disease. Cell Motil Cytoskeleton. 66 (8), 635-649 (2009).
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M. Alzheimer's disease. N Engl J Med. 362 (4), 329-344 (2010).
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B. Genetic modifiers of tauopathy in Drosophila. Genetics. 165 (3), 1233-1242 (2003).
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
