E10.5 dorsal pancreatic progenitors dissociated and seeded in 3D Matrigel recapitulate pancreas development. Progenitors can be most easily followed with fluorescent reporters. In our case we used a transgenic mouse that expresses a nuclear GFP protein controlled by Pdx1 promoter (Pdx1-Ngn3-ERTM-nGFP) (Movie 1) in the absence of tamoxifen and thus without activating Neurog3 4 (Figure 2).
With the organoid medium, an initial compaction of small clusters of cells occurs in the first hours. Expansion can then be detected by the enlargement of the clusters in the first 4 days (Figure 2A). From day 5, branches form in the 20% largest organoids. Single cells do not expand and lose Pdx1 expression while the large clusters retain Pdx1 expression 28.
In these conditions, progenitors undergo a spectacular morphogenesis with the emergence of branched epithelial structures. This process takes place only when the progenitors are seeded in ≥4-cells clusters, indicating a strong requirement for community signals. Histological analysis reveals that after day 7 of culture, the resulting mini-organs are composed of pancreatic progenitors (SOX9+/HNF1B+/Pdx1+ cells: Figure 3B) and differentiated cells expressing either exocrine (Amylase+) or endocrine (Insulin+ or Glucagon+) markers (Figure 3A,C). The differentiation into endocrine cells is lower than in the endogenous pancreas (around 0.1%) but is increased to 1% when FGF1 is not added to the culture medium 28. Remarkably, not only do the seeded progenitors differentiate into the expected pancreatic lineages, but they also spontaneously adopt the normal pancreatic architecture. Although E10.5 multipotent pancreas progenitors are not polarized, the cells in culture polarize as demonstrated by the segregation of Mucin1 and aPKC in the membrane facing the central lumen and they organize into a branched tubular network. Regionalized “tip and trunk” domains emerge: HNF1B+ progenitors and endocrine cells are localized in the central region, while acinar cells are located at the periphery as a partial or complete crown of cells. The organoids can be maintained in culture for 10 days; after this period, they generally lose their architectural organization and become cystic (not shown). Passaging can be done after partial dissociation but quickly leads to cyst formation, a phenomenon that is reduced by the addition of the BMP inhibitor Noggin 28.
With the sphere medium, expansion is more frequent and is seen from 2% of single cells; nevertheless the efficiency of sphere formation correlates with the size of the seeded clusters 28. At day 2/3, a lumen is detected in the small clusters and expands thereafter, leading to largely mono-layered hollow spheres with occasional local multilayered areas (Figure 2B). These spheres collapse when retrieved from Matrigel (Figure 3D-H). Under these conditions, the resulting structures are mainly composed of pancreatic progenitors, with a small percentage of differentiated exocrine and endocrine cells at day 7 (Figure 3D-H). Progenitors in the spheres also become apically polarized, as demonstrated by the segregation of aPKC at the membrane facing the central lumen of all cells (Figure 3F). Pancreatospheres can be passaged at least twice (not shown).
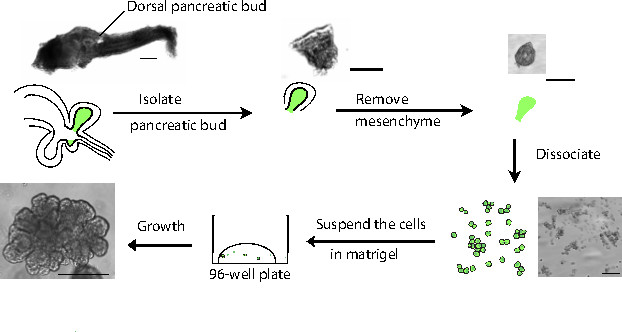
Figure 1. Schematic representation of the procedure. The gastro-intestinal tract is initially dissected from the embryo and subsequently the dorsal pancreatic bud is isolated. The mesenchyme is removed and the pancreatic progenitors are dissociated using trypsin. The resulting partially-dispersed cells are then seeded at low density in growth factor-depleted Matrigel. Scale bars: 1.00 mm except for the resulting organoid picture where the scale bar is 200 µm. Please click here to view a larger version of this figure.

Figure 2. Progress of culture over time. (A) A small cluster of pancreatic progenitors grown with the organoid medium and followed with time-lapse microscopy from 3 hr after plating, for 60 hr. In the bottom panels, an organoid after 7 days of culture. Scale bar: 200 µm; applies to all panels in A. (B) Example of a sphere followed in a 60 hr time-lapse and captured at day 7 of culture. Scale bar: 200 μm; applies to all panels in B. Please click here to view a larger version of this figure.
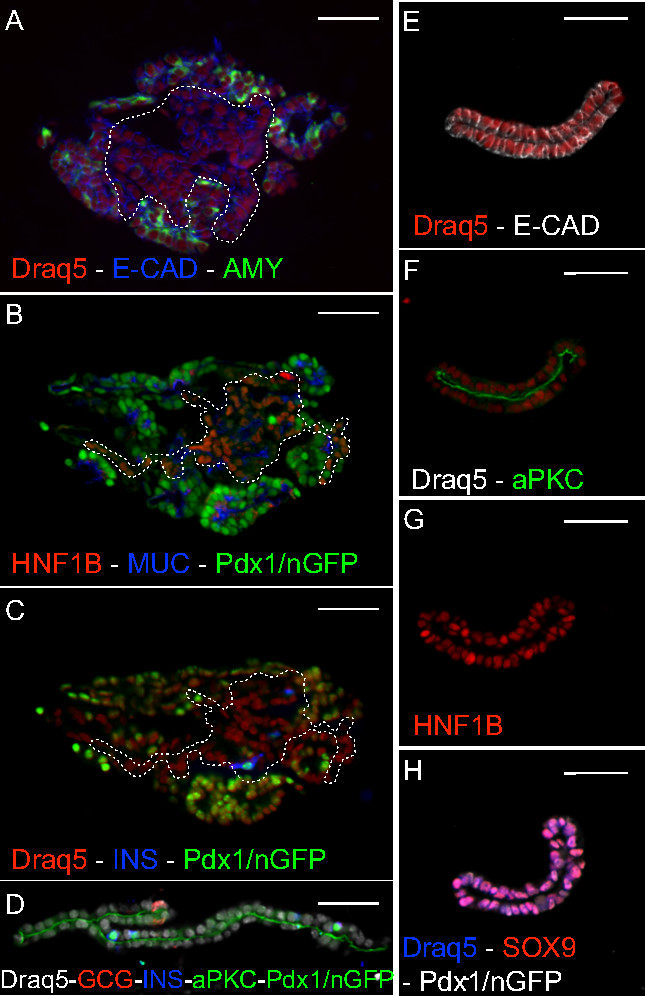
Figure 3. Histology. (A-C) Serial sections of a 7-day organoid stained for progenitor (B – HNF1B) and differentiation (A – amylase, C – insulin) markers. The organoid is composed of epithelial (A – E-cadherin) and apically polarized (B – mucin1) cells. The dashed line corresponds to the non-acinar central region (A), where HNF1B (B) and endocrine (C) cells are detected. (D-H) Sections of 7-day spheres stained for progenitor (G – HNF1B; H – SOX9) and endocrine (D – insulin and glucagon) markers. The spheres are composed of apically polarized (D, F – aPKC) epithelial (E – E-cadherin) cells. Scale bar: 50 µm. Please click here to view a larger version of this figure.
