

Introduction
Den protozoparasit, Trypanosoma brucei, er en agens af sygdomme, der påvirker mennesker (via Trypanosoma brucei gambiense og Trypanosoma brucei rhodesiense) og dyr (via Trypanosoma brucei brucei) i hele Afrika syd for Sahara. Den overføres til pattedyrværten gennem spyt af tsetsefluen vektor. Både menneskers og dyrs afrikanske trypanosomiasis forårsage en alvorlig økonomisk byrde i endemiske områder, og få lægemidler er tilgængelige eller under udvikling til behandling enten sygdom. Forståelse mekanismer immun unddragelse er afgørende for udviklingen af lægemidler til trypanosomiasis. Antigen variation af den tætte, Variant Surface (VSG) pels, der dækker parasitten er en af de primære måder, hvorpå T. brucei undviger den mammale antistofrespons. Ca. 2.000 varianter af VSG-genet findes i trypanosominfektion genomet, men kun én transkriberes til enhver tid fra en af ~ 15 expressionens sites. "Switching" af udtrykte VSG kan ske enten ved at kopiere en VSG gen i det aktive udtryk site, eller ved transkriptionel aktivering af en tidligere tavs udtryk websted (revideret i 1).
Selvom meget arbejde har været afsat til at forstå antigen variation i T. brucei, de faktorer, der påvirker skifte frekvens er stadig dårligt forstået. Undersøgelser til dato er blevet vanskeliggjort af, at mens skift sker stokastisk in vitro, skifte frekvenser er meget lave, i størrelsesordenen 1 i omkring 10 6 celler 2. Dette gør det vanskeligt at måle, om en given faktor stiger eller falder skifte frekvens, da skift er svært at opdage i første omgang. Før 2009, metoder til isolering omskiftere i en given population var langvarig og arbejdskrævende. Disse omfattede passage trypanosomer gennem mus immuniseret mod den dominerende VSG og derefter høste celleren dag senere 3, eller tælle hundredvis af celler ved immunofluorescens 4,5. En anden strategi er afhængig af selektion for resistens at isolere switchers 6. Fordi afrikanske trypanosomer dyrket in vitro er typisk består af en stor befolkning udtrykker en større variant, og en meget mindre population af switchers udtrykker alternative varianter, henviser vi til den store variant hele dette papir som den dominerende, startende VSG. Dermed vi ikke på nogen måde ønsker at antyde, at denne store variant har større kondition end andre varianter i populationen.
Her beskriver vi en metode, som pålideligt kan måle antallet af trypanosomer udtrykker et ikke-dominerende VSG i en given population i 3 - 4 timer. Denne fremgangsmåde er særlig nyttig til, når man ønsker at konstatere, om en given genetisk manipulation eller lægemiddelbehandling øger antallet af koblede celler i en population. I stedet for at befri populationen af celler, der udtrykker dominant startede VSG gennem narkotika eller ved immunologiske midler, er disse celler elimineres ved først at overtrække dem med magnetiske perler koblet til antistof mod den dominerende VSG og derefter isolere dem på en magnetisk søjle. Det koblede population opsamles derefter i gennemløbet og farvet igen med en fluorofor mærket anti-VSG-antistof til at identificere forureninger. Kvantificering opnås ved at tilsætte et defineret antal absolutte optælling perler til hver prøve, således at kan bestemmes forholdet mellem perler og celler og anvendes til at kvantificere antallet af omskiftere i populationen 7.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
BEMÆRK: I hele den procedure, er det nødvendigt at holde celler på is. Kolde medier bør også anvendes overalt.
1. Prøve Høst
- Grow Lister 427 stamme blodbanen trypanosomer til en densitet på 0,5 - 1 million / ml. Det er bedst at starte kulturerne med et lille antal parasitter. Spin down 50 x 10 6 celler / prøve i 10 minutter ved 1.500 x g. Vær sikker på at efterlade 1 x 10 6 celler i kultur til senere brug som positive og negative antistof kontroller.
BEMÆRK: Denne protokol kan også bruges på trypanosomer isoleret fra dyreblod (se diskussionen for detaljer). - Pipette eller hæld det meste af supernatanten (forlade ca. 750 pi). Overfør cellerne til et 1,5 ml mikrocentrifugerør.
- Spin celler ved 4 ° C i 4 minutter ved 5.200 xg i en mikrocentrifuge og fjern supernatanten.
2. Magnetisk Mærkning
- Resuspender celler i 150 pi kulturmedium (HMI-9 with serum for eksempel) + primære anti-VSG antistof ved passende fortynding.
BEMÆRK: anti-VSG-antistof er fremstillet i huset og anvendt i en fortynding på 1:50. Det er vigtigt at bruge et primært anti-VSG antistof, der ikke er mærket med en fluorofor. - Vortex celler under anvendelse af en vortex-adapter-60 ved hastighed 6 - 8 i et kølerum ved 4 ° C i 10 min. Vask med 800 - 1.000 pi kold HMI-9 med serum.
- Centrifugering ved 4 ° C i 4 minutter ved 5.200 xg og aspirere supernatanten. Vask med 1 ml HMI-9 med serum og spin som ovenfor. Aspirere supernatanten. Resuspender pellet i 100 pi HMI-9 med serum.
- Tilføj 110 pi magnetisk cellesortering (MSC) mikroperler. Bruge anti-muse, anti-kanin eller anti-biotin perler som passende til det primære anti-VSG-antistof. Bland godt. Vortex celler i et kølerum ved 4 ° C i 10 min.
- Mens celler er vortexing, oprettet en magnetisk aktiveret separation kolonne / prøve på en adskillelse magnet. Vær sikker på at have en receptacle (vi bruger en 15 ml centrifugeglas) under kolonnen til opsamling gennemløbet. Opstil apparatet i et koldt rum.
- Tilsæt 2 ml HMI-9 med serum til at prime kolonne. Efter grunding sted en 15 ml centrifugerør i kolonnen til opsamling gennemstrømning fra hver prøve.
- Efter 10 minutters inkubation i trin 2.4, tilsættes 800-1000 pi kold HMI-9 med serum. Centrifugering ved 4 ° C i 4 minutter ved 5.200 xg og fjern supernatanten for at fjerne ubundet antistof. Vask med 1 ml HMI-9 med serum og gentag forlader 100 pi af supernatanten.
- Flick centrifugerøret energisk og visuelt inspicere røret for at sikre, at pillen er fuldstændig resuspenderes. Tilsæt 1 ml HMI-9 med serum og pipette op og ned for at sikre, at celler er godt resuspenderes.
3. Magnetisk Separation
- Sørg for, at søjlerne er primet som beskrevet ovenfor. Påfør celler til kolonne. Saml gennemstrømning med celler, der udtrykker ikke-dominerende VSG. 1 ml medium + trypanosomer tager typisk 6 - 7 min at strømme gennem søjlen.
- Vask 2x med 1 ml HMI-9 med serum og indsamle gennemstrømning i det samme rør som i trin 3.1.
- Divide gennemstrømning (3 ml) i to 1,5 ml mikrocentrifugerør.
BEMÆRK: Disse kan kombineres senere, eller halvdelen kan anvendes til at isolere RNA, etc. - Fjern kolonne fra magneten. Tilsæt 3 ml HMI-9 med serum til søjle og kaste kolonne i et 15 ml centrifugerør til opnåelse celler, som udtrykker start, dominerende VSG. Fjern 300 pi eluerede materiale til senere analyse. Hold disse celler på is.
- Til anvendelse som en positiv og en negativ kontrol, tage cirka 1 million celler fra udgangs-kultur af kontrolceller vokser ved 37 ° C (dem som ikke forventes at skifte ofte) og pipette dem i et 1,5 ml mikrocentrifugerør.
BEMÆRK: Positive kontrol celler vil farves med fluorofor-mærket antistof i trin 4.1. Negativ kontrol celler remai ufarvede hele resten af protokollen. - Spin gennemstrømningssystemer prøver og positiv prøve antistofkontrol ved 4 ° C i 4 minutter ved 5.200 xg og fjern supernatanten.
BEMÆRK: En anden 300 pi alikvot af eluerede celler kan anvendes som en positiv kontrol for anti-VSG-farvning.
4. Farvning at identificere Forureninger og Skiftere
- Resuspender gennemstrømningssystemer prøver og positive kontrolceller i 100 pi HMI-9 med serum + primær, fluorofor-mærket, anti-VSG antistof ved en passende fortynding.
BEMÆRK: mærket anti-VSG-antistof er fremstillet i huset og anvendt i en fortynding på 1: 1.000. Fluoroforen-mærkede, kan anti-VSG være fra den samme kilde som det umærkede antistof i MACS separation step.There skal være 3 ml gennemstrømning prøve i to mikrocentrifugerør følgende trin 3.5. - At analysere alle gennemstrømningen ved flowcytometri, kombinere pellets i de to mikrocentrifugerør mens re-susafventning prøverne i 100 pi HMI-9 med serum + anti-VSG-antistof. Bland godt.
- Vortex celler i et kølerum ved 4 ° C i 10 min. Tilføj 800 - 1.000 pi kold HMI-9 med serum. Spin prøver ved 4 ° C i 4 minutter ved 5.200 xg og fjern supernatanten for at fjerne ubundet antistof.
- Vask cellerne med 1 ml af HMI-9 med serum. Spin prøver ved 4 ° C i 4 minutter ved 5.200 xg og fjern supernatanten. I løbet af denne sidste tur, spin ned de eluerede-prøver fra trin 3.4 og den negative kontrolprøve.
- Resuspender al gennemstrømning og eluerede prøver i: 148,5 pi HMI-9 med serum, 25 pi absolut optælling perler og 1,5 pi propidiumiodidfarvning opløsning (175 pi alt). Resuspender positive og negative antistof kontrolprøver i 175 pi HMI-9 med serum. Sørg for at registrere antallet af absolutte tælle perler / 25 pi for bestemt parti anvendes.
- Kør alle prøver gennem et flowcytometer og indsamle data. Døde celler plette som PI
- Beregn skifte frekvenser ved hjælp af flowcytometri data.
BEMÆRK: For instruktioner om, hvordan man beregner skifte frekvenser, henvises til supplerende fil flowcytometri Calculations.docx og tabel 1.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Den her beskrevne metode blev anvendt til at påvise, at dobbelt-strenget pauser i VSG udtryk stedet øget antallet af omskiftere i en population 8. Her viser vi repræsentative resultater fra en population af trypanosomer, som er blevet tilsvarende induceres til at frembringe en dobbelt-strenget pause på ekspressionsstedet. Vi sammenligner disse trypanosomer til dem der ikke er blevet induceret til at generere en dobbeltstrenget brud. Figur 1 viser repræsentative flowcytometri plots fra un-inducerede og inducerede celler indsamlet i gennemløbet fra den magnetiske cellesortering kolonne. For denne særlige eksperiment den dominerende start VSG var VSG2. Den venstre paneler af figur 1A viser frem og side scatter, som giver porte til at blive trukket rundt om de absolutte optælling perler og celler, der viser en fremadrettet og sidespredning signatur, der er karakteristisk for levende celler. Bemærk, at porte kandrages til at omfatte flere celler, fordi døde celler kan elimineres med PI farvning; dog skal porten være det samme for alle prøverne. De højre paneler af figur 1A viser kun de celler, der falder ind under "Live" gate på venstre hånd paneler. Celler, plette positivt for PI (Q1 og Q2) er døde celler. Celler, plette positivt for den dominerende VSG og negativt for PI i 3. kvartal er forurenende stoffer. Celler, plette negativt for både PI og dominerende VSG i 4. kvartal er levende celler, der har skiftet. Man kan konstatere, at procentdelen af forureninger i Q3 er lavere for de celler, hvor en dobbelt-strenget pause er blevet induceret. Imidlertid kan procenterne i Q3 ikke bruges til at udlede, om der er flere omskiftere i en given population. Det er kun ved at sammenligne forholdet mellem antallet af celler i Q4 til antallet af perler opsamlet at man kan beregne omskiftningsfrekvenser. Tabel 1 viser antallet opnået ved flow cytometri plots og metoden til beregning af de procent omskiftere i disse populationer. Disse beregninger blev udført på 3 biologiske prøver for at opnå de observerede koblingsfrekvens vist i figur 1B. Niveauet af stokastisk skift in vitro er ganske lav, som beregnet her på et gennemsnit på 9,53 x 10 -5, mens inducere en pause resulterer i 1 - 6 x 10 -3 celler, som har skiftet til den ikke-dominerende VSG.
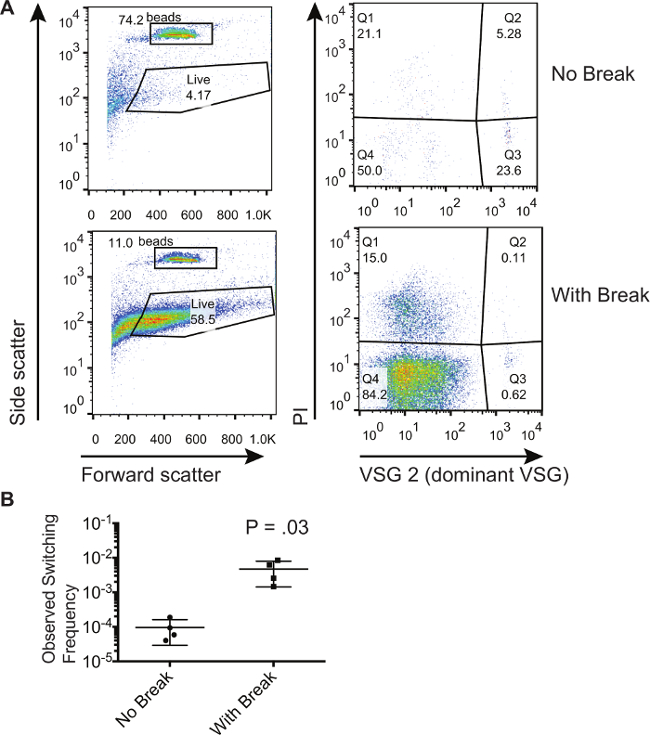
Figur 1. Isolering og Kvantificering af Switched trypanosominfektion populationer. (A) Flowcytometri viser fraktioner af trypanosomer i gennemløbsfraktionen efter udvælgelse af ikke-omskiftere på en magnetisk aktiveret cellesortering kolonne. En population af celler med en induceret I-SCEI dobbelt-strenget pause er sammenlignet med en population af celler uden en induceret break. (B) Beregnet skifte frekvenser til celler, der har eller ikke er blevet induceret med en I-SCEI dobbeltstrenget pause. Nummeret på grunden repræsenterer resultaterne af en to-halet, uparret t-test. Fejl søjler repræsenterer SD. Klik her for at se en større version af dette tal.
Supplerende Fil 1. Flowcytometri Beregninger. Klik her for at downloade denne fil.

Tabel 1. Beregning af switchfrekvenser. Klik her for at se en større version af denne tabel.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Med hensyn til eksperimentel teknik, er den mest kritiske komponent af protokollen holde alle prøverne kolde. Trypanosomer meget hurtigt internalisere antistof bundet til deres overflade 9, men denne proces er motilitet afhængig, og påvirker ikke assayet, så længe cellerne holdt ved 4 ° C. Alle prøver skal altid holdes på is, og pipettering bør ske hurtigt for at minimere eksponering for 25 ° C lab miljø. Kolde HMI-9 med serum bør være tilgængelige i starten af forsøget, og bør holdes på is i hele. Isolering af trypanosomer ved at køre dem over kolonnen skal ske i et koldt rum, som det tager 5 - 7 min at køre en 1 ml prøve gennem søjlen. Hvis der er flere prøver end magnetisk separation enheder til rådighed, kan magnetisk isolering gøres i rækkefølge, så længe prøverne ikke aktivt adskilt holdes på is.
Det er også afgørende, at prøverne være meget godt blandet AFter den sidste vask efter inkubation med mikroperlerne og før isolering på magneten. Dette kan opnås ved aggressiv flicking af centrifugeglasset eller lys hvirvelbehandling. Nr pellet eller klumper af celler skal være synlige før tilsætning af prøven til separationskolonnen, hvilket kan kontrolleres ved at se på lysmikroskopi niveau. Hvis klumper er til stede, forureningsniveauet i gennemløbet prøve af celler, der udtrykker den dominerende VSG er normalt meget højere end ønskeligt. Når der udføres beregningen for antallet af omskiftere i prøven ved slutningen af protokollen, er det vigtigt at tage hensyn til, om alle gennemløbet blev anvendt til flowcytometrisk analyse, eller om halvdelen blev anvendt. Hvis der blev anvendt kun halvdelen, bør det samlede antal omskiftere beregnede multipliceres med 2.
Når du fjerner supernatanten under vasketrin, er det ikke nødvendigt at pipette ud hver sidste dråbe, og typisk 15 - 25 pi erstarter omkring pellet af celler. Dette er især kritisk i de sidste faser af farvning og vask, for på dette tidspunkt pellets sjældent set i flow-through prøver, fordi der er så få skiftet celler.
Mens den endelige anti-VSG plet udføres typisk med oprenset, fluorofor-mærket anti-VSG-antistof, kan den første anti-VSG plet inden magnetisk isolering gøres med oprenset antistof eller serum. Vi har også brugt bioreaktor supernatant afledt af hybridomer der udtrykker anti-VSG-antistoffer. Uanset hvad, skal den bruges til den indledende pletten materiale titreres for at sikre god adskillelse af celler, der gør eller ikke udtrykker den dominerende, startende VSG.
Det er typisk for populationen af celler i gennemløbet at være kontamineret med celler, der udtrykker den dominerende VSG. Mens denne forurening bør være minimal, i vores erfaring er det sjældent, at befolkningen at være helt blottet for celler udtrykkering den dominerende VSG. Det er også vigtigt at bemærke, at celler, der er gået gennem isolation procedure, ikke plette så smukt med anti-VSG antistof som dem, som ikke har været igennem proceduren. Vi hypotesen, at to ting kunne redegøre for denne forskel. Den første er, at spinning trypanosomer medfører spredning af VSG, og der er mange spin involveret i isolation procedure. Dette ville forventes at mindske intensiteten af signalet ved den endelige anti-VSG plet. Den anden er, at det første trin i protokollen involverer farvning med et primært anti-VSG antistof. Hvis epitoper genkendt af antistoffet anvendt i dette første trin er de samme som eller okkluderer epitoperne genkendt af antistoffet i den endelige pletten, ville man forvente intensiteten af signalet til at være lavere, end hvis kun ét antistof blev anvendt. Af denne grund er det vigtigt at titrere antistof, som anvendes i det sidste trin, sådan at signalet fra en positivt farvede celle er omtrent tostørrelsesordener højere end for en negativt farvet celle. Denne måde, selv om intensiteten af signalet fra en positivt farvning celle formindskes i celler, der har været underkastet den isolation procedure, disse positive celler vil stadig være godt adskilt fra de negative celler på flowcytometri plot. Hvis der anvendes auto-kompensation, skal prøver enkelt plet kontrol være forberedt. Antistoffer, som vi har genereret er tilgængelige for køb fra Memorial Sloan Cancer Center monoklonalt antistof facilitet. Vi har haft den bedste succes med anvendelse af umærket monoklonalt IgG-antistof. Vi har udført protokollen med polyklonale VSG antistoffer, men polyklonale antistoffer kan lejlighedsvis genkende andre VSGs så monoklonale antistoffer foretrækkes. VSG identitet kan bestemmes ved sekventering VSG cDNA dannet fra amplifikation af konserverede sekvenser i VSG 3'UTR og den splejsede leder. En særligt antistof kan testes for specificitet på tidligere isolerede tændtceller, der er VSG identitet er blevet bestemt som netop beskrevet.
Protokollen præsenteres her har en række fordele. Den kan bruges både til at isolere switchers til senere analyse og kvantificere antallet af omskiftere i en given population. Den kan også anvendes til at isolere en bestemt variant af interesse, så længe et antistof er tilgængelig for denne variant. Det er også muligt at udføre denne protokol under anvendelse trypanosomer isoleret fra dyr, forudsat at de røde blodlegemer er elimineret under anvendelse af anti-TER119 overtrukne-magnetiske kugler ifølge fabrikantens protokol. Vi har ikke testet den nedre grænse for antallet af trypanosomer nødvendige, men vi har med succes gennemført protokollen hjælp 2,5 - 10 millioner trypanosomer fra 250 pi af blod. Endelig kan protokollen anvendes sammen med udsving analyse for at få frekvensen af skift i en given population.
Proceduren for isolation er ganske fast, idet 3 - 4 timer fra start til slut afhængigt af antallet af prøver, og således er mere effektiv end tidligere fremgangsmåder, der krævede forudgående immunisering af mus eller specialiserede stammer indeholdende lægemiddel-resistens-markører. Fremgangsmåden er begrænset af det faktum at antistoffer mod udgangs VSG kræves imidlertid. Uden sådanne reagenser, kan en alternativ metode, såsom VSG-seq være et mere passende valg til at måle hvilke VSGs bliver udtrykt i en given population 10.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Vi vil gerne anerkende George Cross til generel rådgivning om trypanosominfektion biologi. Dette arbejde blev også støttet af en Bill og Melinda Gates Foundation GCE tilskud til DS, en NSF Graduate Research Fellowship (DGE-1.325.261) til MRM og en NIH / NIAID (tilskud # AI085973) til FNP. Vi takker Galadriel Hovel-Miner til anvendelse af stamme indeholdende I-SCEI genet og genkendelsessted.
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
