

Introduction
寄生原虫、 トリパノソーマは 、サハラ以南のアフリカ全域( トリパノソーマブルセイを介して)や動物( トリパノソーマガンビアとトリパノソーマローデシア経由)人間に影響を与える疾患の原因物質です。なお、ツェツェバエベクターの唾液を介して哺乳動物宿主に伝達されます。人間と動物、アフリカトリパノソーマ症の両方が流行地域に深刻な経済的負担を引き起こす、といくつかの薬が利用可能であるか、またはいずれかの疾患を治療するために開発中です。免疫回避のメカニズムを理解することは、トリパノソーマ症用薬の開発のために重要です。寄生虫をカバーして密な、バリアント表面糖タンパク質(VSG)コートの抗原性の変化は、主要な手段の一つであることでT.トリパノソーマは、哺乳類の抗体応答を逃れます。 VSG遺伝子の約2,000の変異体は、トリパノソーマのゲノムに存在するが、一つだけは〜15 EXPRESの1から任意の時点で転写されますシオンサイト。表現VSGの「切り替えは、「アクティブ発現部位にVSG遺伝子をコピーすることによって、または(1件)以前にサイレント発現部位の転写活性化のいずれかによって発生する可能性があります。
多くの仕事は、Tにおける抗原変異を理解することに専念してきたがトリパノソーマは 、スイッチング周波数の影響を与える要因は、まだよく理解されていません。これまでの研究では、約10 6セル2 1の順に、スイッチングがインビトロで確率的に発生している間、スイッチング周波数が非常に低いという事実によって妨げられてきました。これは、それが困難なスイッチングが最初の場所で検出が困難であるため、スイッチング周波数を減少させ、所定の係数が大きくなるかどうかを測定することができます。 2009年以前には、所定の集団でスイッチャーを単離するための方法は、長く、労働集約的でした。これらには、が支配的VSGに対して免疫化したマウスを通じてトリパノソーマを継代した後、細胞を回収日後3、または免疫蛍光4,5によって細胞の数百を数えます。別の戦略は、スイッチャー6を単離するために、薬剤耐性の選択に依存しています。 インビトロで増殖アフリカトリパノソーマは、典型的には、1つの主要な変異体、および代替変異体を発現スイッチャのはるかに小さい人口を表現する大集団で構成されているので、我々は支配的な、出発VSGとしてこの論文を通じて、主要なバリアントを参照してください。そうすることで、私たちは決してこの主要な変異体は、集団内の他の変異体よりも大きな適応度を持っていることを暗示したいです。
4時間 - ここでは、確実に3に与えられた集団における非支配的なVSGを発現しているトリパノソーマの数を測定することができる方法について説明します。この方法は、1つの与えられた遺伝子操作または薬物治療が集団に切り替え細胞の数を増加させるかどうかを確認したい場合に特に有用です。代わりにDOを発現する細胞の集団をなくししますminantは、薬を介して、または免疫学的手段によってVSGを開始し、これらの細胞を最初に支配VSGに対する抗体に結合された磁気ビーズでそれらをコーティングした後、磁気カラム上でそれらを分離することによって除去されます。切り替え集団は、フロースルーに回収し、汚染物質を識別するためのフルオロフォア標識した抗VSG抗体で再度染色します。定量化は、細胞に対するビーズの比を決定し、集団7にスイッチャの数を定量するために使用することができるように、各サンプルの絶対計数ビーズの定義された数を加算することによって達成されます。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
注:手順を通して、それを氷上で細胞を維持することが必要です。コールドメディアも全体で使用されるべきです。
1.サンプル収穫
- 100万/ mlの - 0.5の密度にリスター427株血流のトリパノソーマを成長させます。これは、寄生虫数の少ない培養を開始するのが最善の方法です。 1500×gで10分間、50×10 6細胞/サンプルをスピンダウン。正と負の抗体コントロールとして後で使用するために文化の中で1×10 6個の細胞を残すようにしてください。
注:このプロトコルは、(詳細説明を参照)、動物の血液から単離されたトリパノソーマに使用することができます。 - ピペットまたは上清の大部分を捨てる(約750μLを残します)。 1.5ミリリットルマイクロ遠心チューブに細胞を移します。
- 微量で5200×gで4分間、4℃で細胞をスピンし、上清を除去します。
2.磁気標識
- 150μlの培養培地中で再懸濁細胞(HMI-9のwi適切な希釈で例えば番目血清)+一次抗VSG抗体。
注:抗VSG抗体は、家の中で作られ、1時50分の希釈で使用されています。フルオロフォアで標識されていない一次抗VSGの抗体を使用することが重要です。 - 10分間、4℃の低温室に8 - スピード6で渦アダプタ-60を使用して、渦細胞。血清で千μlの冷間HMI-9から800で洗います。
- 5200×gで4分間、4℃でスピンし、上清を吸引除去します。血清を1ミリリットルHMI-9で洗浄し、上記のようにスピン。上清を吸引除去します。血清とHMI-9の100μlの再懸濁ペレット。
- (MSC)マイクロビーズをソートする磁気活性化細胞の110μLを加えます。一次抗VSG抗体に応じて抗マウス、抗ウサギ、または抗ビオチンビーズを使用してください。よく混ぜます。 10分間、4℃の低温室での渦細胞。
- 細胞はボルテックスされているが、分離磁石に1磁気活性化分離カラム/サンプルを設定します。再を持っているようにしてくださいフロースルーを収集する列の下ceptacle(私たちは15ミリリットルの遠心分離管を使用します)。低温室で装置を設定します。
- プライムカラムに血清を使用して、HMI-9の2ミリリットルを追加します。フロースルー各試料から収集する列の下に15ミリリットルの遠心分離管場所をプライミングした後。
- ステップ2.4で10分間インキュベーションした後、血清をコールドHMI-9 800〜μlを添加します。 5200×gで4分間、4℃でスピンし、上清を除去し、未結合抗体を除去します。血清およびリピートは100μlの上清を残して、HMI-9の1ミリリットルで洗浄します。
- 激しく視覚的に遠心管をフリックペレットを完全に再懸濁されていることを確認するために、チューブを点検します。細胞が十分に再懸濁されていることを確認するには、上下の血清およびピペットでHMI-9の1ミリリットルを追加します。
3.磁気分離
- 上記のように列がプライミングされていることを確認します。カラムに細胞を適用します。フロースルー非支配VSを発現する細胞とを収集カラムを通って流れるように7分 - G. 1 mlの培地+トリパノソーマは、一般的に6を取ります。
- ウォッシュ血清とHMI-9の1ミリリットルで2回およびフロースルー同じチューブのステップ3.1のように集めます。
- 除算フロースルー(3ミリリットル)2 1.5ミリリットルのマイクロチューブに。
注:これらは、後に組み合わせることができるか、半分など 、RNAを単離することができます - 磁石から列を削除します。カラムに血清を使用して、HMI-9の3ミリリットルを追加し、開始、支配的なVSGを発現する細胞を得るために15ミリリットルの遠心分離管に列を急落。後の分析のために溶出した物質の300μLを削除します。氷の上でこれらの細胞を保管してください。
- 陽性および陰性対照として使用するために、37℃で増殖対照細胞の出発培養物からの約100万個の細胞を取り1.5ml微量チューブに、それらをピペット(それらは頻繁に切り替えることが予想されていません)。
注:ポジティブコントロール細胞はステップ4.1でフルオロフォア標識抗体で染色されます。ネガティブコントロール細胞REMAプロトコルの残りの部分全体に染色されていないインチ - スピンフロースルーサンプルおよび抗体陽性対照試料4℃で4分間、5200×gで、上清を除去します。
注:溶出セルの別の300μlのアリコートを、抗VSG染色の陽性対照として使用することができます。
4.染色は、汚染物質やスイッチャーを識別するために、
- 再懸濁し、フロースルーサンプルおよび100μlのHMI-9適切な希釈で血清+一次、フルオロフォア標識、抗VSG抗体とにおけるポジティブコントロール細胞。
注:標識抗VSG抗体は家で作られ、1の希釈で使用されている:1,000。フルオロフォア標識、抗VSGはstep.Thereは、ステップ3.5以下の二つのマイクロ遠心チューブ内のフロースルーサンプルの3ミリリットルであるべきMACS分離で非標識抗体と同じソースからすることができます。 - フローサイトメトリーによってフロースルーのすべてを分析するために、一方の再SUS 2マイクロ遠心チューブ内でペレットを結合血清+抗VSG抗体を使用して、HMI-9の100μl中のサンプルを保留中。よく混ぜます。
- 10分間、4℃の低温室での渦細胞。千μlの冷間HMI-9血清との - 800を追加します。スピンサンプル5200×gで4分間、4℃で、非結合抗体を除去するために、上清を除去します。
- 血清とHMI-9の1ミリリットルで細胞を洗浄。スピンサンプル5200×gで4分間、4℃で、上清を除去します。この最後のスピンの間に、ステップ3.4および陰性対照試料から溶出し、サンプルをスピンダウン。
- 血清で148.5μlのHMI-9、25μlの絶対計数ビーズと1.5μlのヨウ化プロピジウム染色溶液(175μlの合計):内のすべてのフロースルーと溶出サンプルを再懸濁します。 175μlの正および負の抗体対照試料を再懸濁HMI-9の血清で。絶対計数ビーズ/使用される特定のロットの25μlの数を記録してください。
- フローサイトメーターを介してすべてのサンプルを実行し、データを収集します。死細胞は、PIとして染色します
- フローサイトメトリーデータを使用してスイッチング周波数計算します。
注:スイッチング周波数を計算する方法については、補足ファイルを参照してくださいフローサイトメトリーCalculations.docxおよび表1。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
ここで説明する方法は、VSG発現部位内の二本鎖切断は、人口8にスイッチャーの数を増加させたことを実証しました。ここでは、同様に発現部位で二本鎖切断を生成するように誘導されているトリパノソーマの集団からの代表的結果を示します。私たちは、二本鎖切断を生成するように誘導されていないものにこれらのトリパノソーマを比較する。 図1は、フロースルーの列をソートする磁気活性化細胞からに集め、非誘導し、誘導された細胞からのプロット代表的なフローサイトメトリーを示しています。この特定の実験のために支配的な出発VSGはVSG2ました。 図1Aの左側のパネルは、前方に表示され、ゲートが生細胞の特徴である前方および側方散乱シグネチャを示す絶対計数ビーズと細胞の周りに描かれることを可能にする側方散乱、。ゲートができることに注意してください死細胞はPI染色を用いて除去することができるので、より多くの細胞を含むように描かれます。しかし、ゲートがすべての試料について同じに維持されなければなりません。 図1Aの右側のパネルには、左側のパネルにある「ライブ」のゲートに入る細胞のみを示します。 PI(Q1とQ2)に対して陽性染色細胞は死んだ細胞です。第3四半期にPIのために積極的に支配的なVSGおよび負染色細胞が汚染物質です。第4四半期にPIと支配的なVSGの両方に負染色細胞は、切り替えた生細胞です。一つは、第3四半期中の汚染物質の割合は、二本鎖切断が誘導されたそれらの細胞のために低くなっていることを観察することができます。しかし、第3四半期のパーセンテージは、所定の集団内のより多くのスイッチャーがあるかどうかを推測するために使用することはできません。それは唯一のスイッチング周波数を計算することができることを集めたビーズの数をQ4での細胞数の比を比較することである。 表1は、FLOから得られた数値を示していますワットサイトメトリープロットおよび方法は、これらの集団のパーセントスイッチャーを計算します。これらの計算は、図1Bに表示された観察されたスイッチング周波数を得るために3生物学的サンプルについて行きました。非支配的なVSGに切り替えた6×10 -3細胞- 1でブレーク結果を誘導しながら、9.53×10 -5の平均で、ここで計算されるようなin vitroでの確率論的スイッチングのレベルは、非常に低いです。
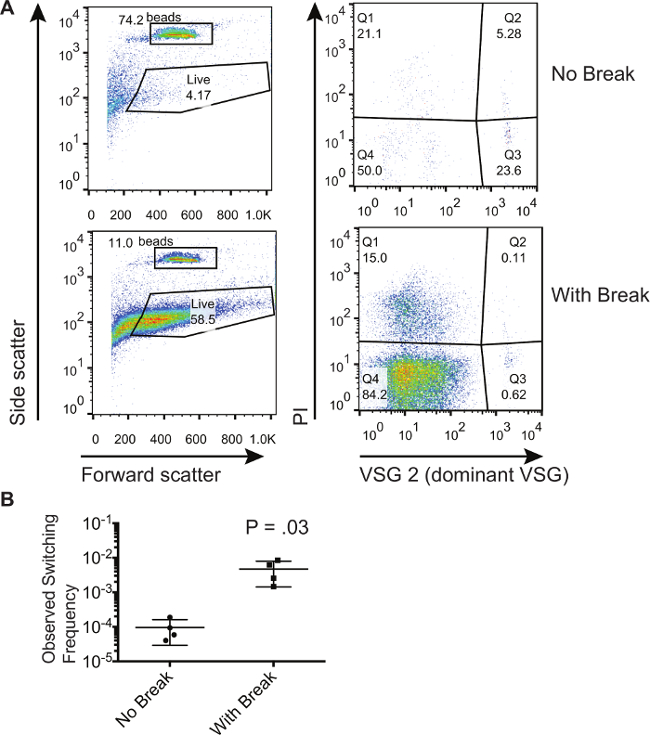
図1. 単離と交換トリパノソーマ集団の定量化。 (A)、磁気活性化細胞選別カラム上の非スイッチャの選択以下のフロースルー画分にトリパノソーマの画分を示すフローサイトメトリー。誘導されたI-SCEI二本鎖切断が誘導BREずに細胞の集団と比較すると細胞の集団AK。(B)I-SCEIの二本鎖切断を用いて誘導されていないか、またはしている細胞の周波数を切り替えて計算されます。プロット上の数は、両側検定、対応のないt検定の結果を表します。エラーバーはSDを表す。 この図の拡大版をご覧になるにはこちらをクリックしてください。
補足ファイル1.フローサイトメトリーの計算。 このファイルをダウンロードするにはこちらをクリックしてください。

スイッチング周波数の表1計算。 この表の拡大版をご覧になるにはこちらをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
実験技術に関しては、プロトコルの最も重要なコンポーネントは、冷たいすべてのサンプルを保っています。トリパノソーマは非常に迅速に、それらの表面9に結合した抗体の内在化が、このプロセスは、運動性に依存し、細胞を4℃に維持される限り、アッセイに影響を及ぼしません。全てのサンプルは、常に氷上で保存されるべきで、ピペッティングを25℃の実験室環境への暴露を最小限にするために迅速に行われるべきです。コールドHMI-9血清では、実験の開始時に利用可能であるべきであり、全体を通して氷上に保持する必要があります。カラムを通って1ミリリットルのサンプルを実行するには、7分 - それは5を取るように、列の上にそれらを実行して、トリパノソーマの単離は、低温室で行わなければなりません。利用可能な磁気分離ユニットよりも多くのサンプルが存在する場合、磁気分離であれば積極的に分離されていない試料を氷上に保持されるように、順序で行うことができます。
試料が非常によく混合AFであることも重要です磁石上のマイクロビーズと、単離前にとのインキュベーション後の最後の洗浄ター。これは、遠心分離管または光ボルテックスを積極的にフリックすることによって達成することができます。細胞のいかなるペレットまたは塊が前に光学顕微鏡レベルで見てチェックすることができます分離カラムへの試料の添加に見えてはいけません。塊が存在する場合、ドミナントVSGを発現する細胞のフロースルーサンプル中の汚染レベルは、通常、望ましいよりもはるかに高いです。プロトコルの終わりに、試料中のスイッチャーの数の計算を行う際には、すべてのフロースルーは、フローサイトメトリー分析のために使用されたかどうかを考慮することが重要である、または半分が使用されたかどうか。半分だけを使用した場合、計算されたスイッチャの総数が2で乗算されるべきです。
洗浄工程の間に上清を除去した場合、最後の一滴をオフにピペットする必要がなく、一般的に15から25μlをしています細胞のペレットを囲む左。非常に少ない切り替えセルがあるので、この時点では、ペレットはほとんどフロースルーサンプルでは見られないので、これは、染色及び洗浄の最終段階において特に重要です。
最終的な抗VSG染色は、典型的には精製された、フルオロフォア標識抗VSGの抗体を用いて行われている間、前の磁気分離の最初の抗VSG染色は、精製された抗体または血清を用いて行うことができます。我々はまた、抗VSGの抗体を発現するハイブリドーマ由来のバイオリアクター上清を使用しています。かかわらず、最初の染色のために使用される材料は、行うまたはドミナント、開始VSGを発現しない細胞の良好な分離を確実にするために滴定する必要があります。
これは、支配的なVSGを発現する細胞で汚染されるように、フロースルー中の細胞の集団に典型的です。この汚染は最小限であるべきであるが、人口が発現する細胞を完全に欠いであるために、私たちの経験では稀です支配的なVSGをる。単離手順を経た細胞の手順ではなかったものとして、抗VSG抗体と同様に明るく染色されないことに留意することも重要です。我々は2つのものが、この違いを説明することができると仮定しました。最初は、トリパノソーマ紡糸するVSGの脱落につながる、そして単離手順に関与する多くのスピンが存在することです。これは、最終抗VSG染色のための信号の強度を減少することが予想されます。第二は、プロトコルの最初のステップは、一次抗VSG抗体を用いて染色を含むことです。この最初の工程で使用した抗体によって認識されるエピトープは、最終染色抗体によって認識されるエピトープと同じかまたは閉塞されている場合、一方が信号の強度が唯一の抗体を使用した場合よりも低いことが予想されます。この理由は、陽性に染色された細胞からの信号は、大きく2であるように、最終段階で使用される抗体を滴定することが重要です負染色された細胞に比べて桁違いに高いです。この方法は、陽性に染色セルからの信号の強度が単離手順を受けた細胞において減少しても、これらの陽性細胞はまだよく、フローサイトメトリープロット上の陰性細胞から分離されます。自動補償が使用されている場合、単一の染色対照試料を調製する必要があります。我々が生成した抗体は、メモリアル・スローンがんセンターするモノクローナル抗体の施設からご購入いただけます。私たちは、非標識モノクローナルIgG抗体を用いた最高の成功を収めています。我々は、ポリクローナルVSG抗体でプロトコルを実行した、ポリクローナル抗体は、時々そのように、モノクローナル抗体が好ましいが、他のVSGを認識することができます。 VSGのアイデンティティは、VSG 3'UTRおよびスプライスさリーダーで保存された配列の増幅から生成されたシーケンシングVSG cDNAによって決定することができます。特定の抗体は、以前に単離された切り替えの特異性について試験することができますVSGのアイデンティティの細胞だけで説明したように決定されています。
ここに提示プロトコルは、多くの利点を有します。後の分析のためにスイッチャーを単離するために、所定の集団におけるスイッチャの数を定量化するための両方に使用することができます。また限り抗体は、その変異体のために利用可能であるように、関心対象の特定の変異体を単離することができます。これは、動物から単離されたトリパノソーマを使用して、この手順を行うことも可能であり、赤血球は、メーカーのプロトコールに従って、抗TER119コーティングされた磁気ビーズを使用して除去されることを条件とします。私たちは、必要なトリパノソーマ数の下限をテストしていないが、我々が正常に2.5使用してプロトコルを実施してきた - 血液250μlのから千万トリパノソーマを。最後に、プロトコルは、所与の集団内のスイッチング周波数を得るために、変動解析と組み合わせて使用することができます。
単離手順は非常にfとAST、3取って - このようにサンプル数に応じて、最初から最後まで4時間、およびは、マウスまたは薬剤耐性マーカーを含む特殊な菌株の前に予防接種を必要とし、従来の方法よりも効率的です。この方法は、出発VSGに対する抗体は、しかしながら、必要とされるという事実によって制限されます。このような試薬がなければ、このようなVSG-seqのよう別の方法は、のVSGは、所定の集団10で表現されているかを把握するために、より適切な選択かもしれません。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
私たちは、トリパノソーマ、生物学上の一般的なアドバイスのためのジョージ・クロスを承認したいと思います。この作品はまた、DS、MRMへのNSF大学院研究フェローシップ(DGE-1325261)及びFNPにNIH / NIAID(助成金#のAI085973)にビル・アンド・メリンダ・ゲイツ財団GCEの助成金によってサポートされていました。我々は、I-SCEI遺伝子と認識部位を含む菌株の使用をガラドリエルあばら家-マイナーに感謝します。
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
