

Introduction
Den protozo parasitt, Trypanosoma brucei, er en utløsende agent for sykdommer som påvirker mennesker (via Trypanosoma brucei gambiense og Trypanosoma brucei rhodesiense) og dyr (via Trypanosoma brucei brucei) i hele Afrika sør for Sahara. Det er overført til pattedyrverts gjennom spytt av Tsetsefluer vektoren. Både mennesker og dyr sovesyke forårsake en alvorlig økonomisk byrde i endemiske områder, og noen stoffer er tilgjengelig eller under utvikling for behandling av enten sykdom. Forstå mekanismene i immununndragelser er avgjørende for utvikling av legemidler for trypanosomiasis. Antigenisk variasjon av det tette, Variant Surface glykoprotein (VSG) pels som dekker den parasitt er en av de primære middel som T. brucei omgår pattedyr antistoffrespons. Omtrent 2000 varianter av VSG-genet eksisterer i trypanosom genomet, men bare en er transkribert til enhver tid fra en av ~ 15 ekspresssjons nettsteder. "Bytte" av uttrykte VSG kan skje enten ved å kopiere en VSG gen inn i aktivt uttrykk nettstedet, eller ved transkripsjonen aktivering av en tidligere taus uttrykk hotellet (anmeldt i 1).
Selv om mye arbeid har vært viet til å forstå antigen variasjon i T. brucei, hvilke faktorer som påvirker bytter frekvens er fortsatt dårlig forstått. Studier har hittil vært hindret av det faktum at mens svitsjing skjer stokastisk in vitro, taktfrekvenser er meget lav, i størrelsesorden av en på rundt 10 6 celler 2. Dette gjør det vanskelig å måle hvorvidt en gitt faktor øker eller synker bytter frekvens, ettersom vekslingen er vanskelig å oppdage i første omgang. Før 2009, metoder for å isolere skiftere i en gitt befolkning var lang og arbeidskrevende. Disse inkluderte aging trypanosomer gjennom mus immunisert mot den dominerende VSG og deretter høsting celleren dag senere 3, eller telle hundrevis av celler ved immunofluorescens 4,5. En annen strategi er avhengig av valg for legemiddelresistens å isolere skiftere 6. Fordi afrikanske trypanosomer dyrket in vitro er vanligvis består av en stor befolkning som uttrykker en større variant, og en mye mindre befolkning på skiftere uttrykker alternative varianter, henviser vi til den store varianten gjennom dette papiret som den dominerende, som starter VSG. Ved å gjøre det vi ikke på noen måte ønsker å antyde at denne store varianten har større trenings enn andre varianter i befolkningen.
Her beskriver vi en fremgangsmåte som på en pålitelig måte kan måle antall trypanosomer som uttrykker et ikke-dominante VSG i en gitt populasjon i med 3 - 4 timer. Denne fremgangsmåten er spesielt nyttig når man ønsker å fastslå hvorvidt et gitt genetisk manipulering eller behandling øker antall svitsjede celler i en populasjon. I stedet for å bli kvitt populasjon av celler som uttrykker dominant, som starter VSG gjennom narkotika eller ved immunologiske midler, disse cellene fjernes ved først å belegge dem med magnetiske kuler koblet til antistoff mot den dominerende VSG og deretter isolere dem på en magnetisk kolonne. Den slått populasjon blir deretter samlet inn i gjennomstrømnings og farget igjen med en fluorofor-merket anti-VSG-antistoff for å identifisere fremmedstoffer. Kvantifisering blir oppnådd ved å legge til et definert antall absolutte telling perler til hver prøve, slik at forholdet mellom perlene til cellene kan bestemmes og brukes til å kvantifisere antallet av omformerenheter i populasjonen 7.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
MERK: Gjennom prosedyren, er det nødvendig å holde cellene på is. Kalde medier bør også brukes hele.
1. Prøve Høsting
- Vokse Lister 427 påkjenning blodet trypanosomer til en tetthet på 0,5 - 1 million / ml. Det er best å starte kulturer med et lite antall parasitter. Spinn ned 50 x 10 6 celler / prøve i 10 min ved 1500 x g. Sørg for å la 1 x 10 6 celler i kultur for senere bruk som positive og negative antistoffkontroll.
MERK: Denne protokollen kan også brukes på trypanosomer isolert fra dyreblod (se omtale for detaljer). - Pipette eller helle av det meste av supernatanten (la ca 750 ul). Overfør cellene til et 1,5 ml mikrosentrifugerør.
- Spin-celler ved 4 ° C i 4 minutter ved 5200 x g i en mikrosentrifuge og fjern supernatanten.
2. Magnetic Merking
- Resuspender celler i 150 ul kulturmedium (HMI-9 with serum for eksempel) + primær anti-VSG-antistoff ved passende fortynning.
MERK: anti-VSG antistoff er gjort i huset og brukes ved en fortynning på 1:50. Det er viktig å bruke en primær anti-VSG-antistoff som ikke er merket med en fluorofor. - Vortex-celler ved hjelp av en vortex-adapter 60 med hastighet 6 - 8 i et kaldt rom ved 4 ° C i 10 min. Vask med 800 - 1000 mL kald HMI-9 med serum.
- Sentrifugering ved 4 ° C i 4 minutter ved 5200 x g og supernatanten aspireres. Vask med 1 ml HMI-9 med serum og spinne som ovenfor. Aspirer supernatanten. Resuspender pellet i 100 mL av HMI-9 med serum.
- Legg 110 mL av magnetisk aktivert celle sortering (MSC) mikroperler. Bruke anti-mus-anti-kanin eller anti-biotin-perler som er hensiktsmessig for den primære anti-VSG-antistoff. Bland godt. Vortex-celler i et kaldt rom ved 4 ° C i 10 min.
- Mens cellene virvling, sette opp en magnetisk-aktivert separasjon kolonne / prøve på en separasjon magnet. Sørg for å ha en receptacle (vi bruker et 15 ml sentrifugerør) under kolonnen for å samle strømningen gjennom. Sett opp anordningen i et kaldt rom.
- Tilsett 2 ml av HMI-9 med serum til prime kolonne. Etter priming sted et 15 ml sentrifugerør under kolonnen for å samle gjennomstrømning fra hver prøve.
- Etter 10 min inkubasjon i trinn 2.4, legge 800-1000 mL kald HMI-9 med serum. Sentrifugering ved 4 ° C i 4 minutter ved 5200 x g og fjern supernatanten for å fjerne ubundet antistoff. Vask med 1 ml HMI-9 med serum og gjenta etterlot 100 ul av supernatanten.
- Flick sentrifugerørets kraftig og visuelt inspisere røret for å sikre at pelleten er fullstendig resuspendert. Tilsett 1 ml av HMI-9 med serum og pipette opp og ned for å sørge for at cellene er godt resuspendert.
3. Magnetic Separasjon
- Sørg for at søylene er malt som beskrevet ovenfor. Påfør cellene til kolonnen. Samle gjennomstrømnings med celler som uttrykker de ikke-dominante VSG. 1 ml medium + trypanosomer tar vanligvis 6 - 7 minutter til å strømme gjennom kolonnen.
- Vask 2 ganger med 1 ml av HMI-9 med serum og samle gjennomstrømning i det samme rør som i trinn 3.1.
- Dividere gjennomstrømnings (3 ml) i to 1,5 ml mikrosentrifugerør.
MERK: Disse kan kombineres senere eller halv kan brukes til å isolere RNA, etc. - Fjern kolonne fra magnet. Tilsett 3 ml HMI-9 med serum til kolonne og styrter kolonne inn i et 15 ml sentrifugerør for å oppnå celler som uttrykker start, dominerende VSG. Fjerne 300 pl av eluert materiale for senere analyse. Oppbevar disse cellene på is.
- For bruk som en positiv og en negativ kontroll, tar ca. 1 million celler fra utgangskulturen av kontrollceller å vokse ved 37 ° C (de ikke forventes å skifte ofte) og pipettere dem inn i et 1,5 ml mikrosentrifugerør.
MERK: Positive kontrollceller vil bli farget med fluorophore-merket antistoff i trinn 4.1. Negative kontrollceller remai unstained hele resten av protokollen. - Spin gjennomstrømnings prøver og positiv antistoffkontrollprøve ved 4 ° C i 4 minutter ved 5200 x g og supernatanten fjernes.
MERK: En annen 300 ul alikvot av eluert celler kan anvendes som en positiv kontroll for anti-VSG farging.
4. Farging å identifisere Forurensninger og omkobler
- Resuspender gjennomstrømnings prøver og positive kontrollceller i 100 pl HMI-9 med serum + primær, fluorofor-merket anti-VSG-antistoff ved passende fortynning.
MERK:-merket anti-VSG-antistoff er gjort i huset og anvendt i en fortynning på 1: 1000. Den fluorofor-merkes, kan anti-VSG være fra den samme kilde som den umerkede antistoffet i MACS separasjons step.There bør være 3 ml gjennomstrømnings prøven i to mikrosentrifugerør følgende trinn 3.5. - For å analysere alle de gjennomstrømnings ved strømningscytometri, kombinere pelletene i de to mikrosentrifugerør mens re-susi påvente av prøvene i 100 ul av HMI-9 med serum + anti-VSG-antistoff. Bland godt.
- Vortex-celler i et kaldt rom ved 4 ° C i 10 min. Legg 800 - 1000 mL kald HMI-9 med serum. Spin prøvene ved 4 ° C i 4 minutter ved 5200 xg og fjern supernatant for å fjerne ubundet antistoff.
- Vask celler med 1 ml HMI-9 med serum. Spin prøvene ved 4 ° C i 4 minutter ved 5200 xg og fjern supernatanten. I løpet av denne siste sentrifugering, spinne ned de eluerte-prøvene fra trinn 3.4 og det negative kontrollprøven.
- Resuspender all gjennomstrømning og eluerte prøver i: 148,5 ul HMI-9 med serum, 25 ul absolutte telling kuler og 1,5 ul propidiumjodidfarging løsning (175 pl totalt). Resuspender positive og negative kontrollprøver antistoff i 175 ul HMI-9 med serum. Sørg for å registrere antall absolutte telle perler / 25 ul for den aktuelle mye brukt.
- Kjøre alle prøvene gjennom et strømningscytometer og samle inn data. Døde celler flekken som PI
- Beregn bytter frekvenser ved hjelp av flowcytometri data.
MERK: For instruksjoner om hvordan du beregner taktfrekvenser, henvises det til supplerende filen flowcytometrisystemer Calculations.docx og tabell 1.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Metoden som beskrives her ble anvendt for å demonstrere at dobbelttrådsbrudd i den VSG uttrykket området økte antall omformerenheter i en populasjon 8. Her viser vi representative resultater fra en populasjon av trypanosomer som har blitt tilsvarende indusert for å generere en dobbel tråd pause på uttrykket området. Vi sammenligner disse trypanosomer til de som ikke er blitt indusert til å generere en dobbel tråd pause. Figur 1 viser representative flowcytometri plott fra ikke-induserte og induserte celler oppsamlet i gjennomstrømnings fra den magnetiske aktivert cellesorteringskolonne. For denne spesielle eksperimentet den dominerende starter VSG var VSG2. Den venstre paneler av Figur 1A viser frem og side scatter, noe som gjør at portene til å bli trukket rundt de absolutte telle perler og celler som viser en frem og side scatter signatur som er karakteristisk for levende celler. Vær oppmerksom på at portene kanbli trukket til å omfatte flere celler fordi døde celler kan elimineres med PI flekker; må imidlertid porten holdes den samme for alle prøvene. Den høyre panel av figur 1A viser bare de cellene som faller innenfor "live" gate på venstre hånd paneler. Celler som flekker positivt for PI (Q1 og Q2) er døde celler. Celler som flekker positivt for den dominerende VSG og negativt for PI i Q3 er forurensninger. Celler som flekker negativt for både PI og den dominerende VSG i Q4 er levende celler som har byttet. Man kan observere at den prosent av forurensninger i Q3 er lavere for disse cellene der en dobbel tråd pause har blitt indusert. Imidlertid kan prosentene i Q3 ikke benyttes til å utlede hvorvidt det er flere skiftere i en gitt populasjon. Det er bare ved å sammenlikne forholdet mellom antallet celler i Q4 til antall perler oppsamlet at man kan regne taktfrekvenser. Tabell 1 viser de tallene som oppnås fra flow Cytometry plott og metoden for å beregne prosent skiftere i disse populasjonene. Disse beregningene ble utført på 3 biologiske prøver for å oppnå de observerte svitsjefrekvenser som vises på figur 1B. Nivået av stokastiske svitsje in vitro er ganske lav, som beregnet her på et gjennomsnitt på 9,53 x 10 -5, mens indusere en pause resultater i 1 - 6 x 10 -3 celler som har byttet til det ikke-dominerende VSG.
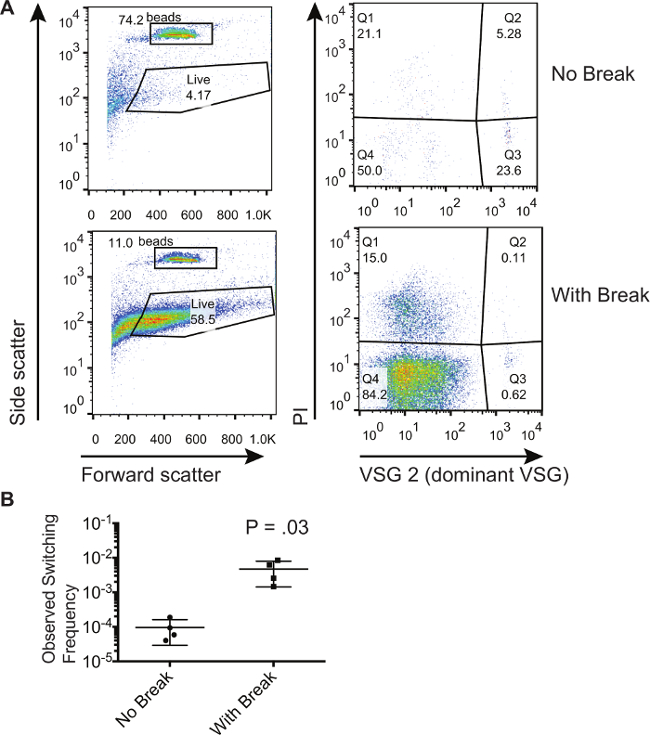
Figur 1. Isolering og kvantifisering av Switched Trypanosom populasjoner. (A) Strømningscytometri viser fraksjoner av trypanosomer i gjennomstrømningsfraksjon følgende utvalg av ikke-skiftere på en magnetisk aktivert cellesortering kolonne. En populasjon av celler med en indusert I-SCEI double-strand pause blir sammenlignet med en populasjon av celler uten en indusert break. (B) Beregnet svitsjefrekvenser for celler som har eller ikke har blitt indusert med en I-SCEI dobbel-strandet pause. Tallet på tomten representerer resultatene av en to-tailed, uparede t-test. Feilfelt representerer SD. Klikk her for å se en større versjon av dette tallet.
Supplemental fil 1. flowcytometri beregninger. Klikk her for å laste ned denne filen.

Tabell 1. Beregning av taktfrekvenser. Klikk her for å se en større versjon av denne tabellen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Med hensyn til eksperimentell teknikk, er den mest kritiske del av protokollen holde alle prøvene kald. Trypanosomer meget hurtig internal antistoff bundet til sin overflate 9, men denne fremgangsmåte er motilitet avhengig, og ikke påvirker analysen, så lenge cellene blir holdt ved 4 ° C. Alle prøver skal alltid holdes på is, og pipettering bør gjøres raskt å redusere eksponering for 25 ° C lab miljø. Cold HMI-9 med serum bør foreligge ved starten av eksperimentet og bør holdes på is i hele. Isolering av trypanosomer ved å kjøre dem over på kolonnen må gjøres i et kaldt rom, som det tar 5 - 7 min for å kjøre en 1 ml prøve gjennom kolonnen. Hvis det er flere prøver enn magnetisk separasjonsenheter som er tilgjengelige, kan magnetisk isolasjon gjøres i rekkefølge, så lenge prøvene ikke aktivt utskilte holdt på is.
Det er også viktig at prøvene, være meget godt blandet after den siste vask etter inkubasjon med mikroperlene og før isolering på magneten. Dette kan oppnås ved aggressiv flicking av sentrifugerøret eller lett risting. Ingen pellet eller klumper av celler som skal være synlig før tilsetning av prøven til separasjonskolonnen, som kan kontrolleres ved å se på lysmikroskopi nivå. Hvis klumper er til stede, graden av forurensning i den gjennomstrømnings prøve av celler som uttrykker den dominerende VSG er vanligvis mye høyere enn ønskelig. Når utfører beregningen for antallet av omformerenheter i prøven ved enden av protokollen, er det viktig å ta hensyn til om alle gjennomstrømnings ble brukt for flowcytometrisk analyse, eller om en halv ble anvendt. Hvis bare halvparten ble brukt, bør det totale antall skiftere beregnet multipliseres med to.
Når du fjerner supernatanten under vasketrinnet, er det ikke nødvendig å pipettere av hver siste dråpe, og typisk 15 - 25 pl ervenstre rundt pellet av celler. Dette er spesielt viktig i løpet av de siste stadier av farging og vasking, fordi på dette tidspunkt pellets er sjelden sett i gjennomstrømnings eksempler fordi det er så få byttet celler.
Mens den endelige anti VSG flekk blir typisk utført med renset, fluorofor-merket anti-VSG-antistoff, kan den første anti-VSG flekk før magnetisk isolering gjøres med renset antistoff eller serum. Vi har også anvendt bioreaktor supernatant avledet fra hybridomer som uttrykker anti-VSG-antistoffer. Uansett må materialet som brukes for den første flekken titreres for å sikre god separasjon av celler som gjør eller ikke uttrykker den dominerende, som starter VSG.
Det er typisk for en populasjon av celler i gjennomstrømnings å være forurenset med celler som uttrykker den dominerende VSG. Selv om dette forurensning bør være minimal, i vår erfaring er det sjelden for befolkningen å være helt blottet for celler uttrykkering den dominerende VSG. Det er også viktig å merke seg at celler som har gått gjennom isoleringsprosedyren ikke flekker som farge med anti-VSG-antistoff som de som ikke har vært gjennom prosedyren. Vi hypotese at to ting kan forklare denne forskjellen. Den første er at spinne trypanosomer fører til shedding av VSG, og det er mange spinn som er involvert i isoleringsprosedyren. Dette ville forventes å redusere intensiteten av signalet for den endelige anti VSG flekken. Den andre er at det første trinnet av protokollen omfatter farging med et primært anti-VSG-antistoff. Dersom de epitoper som gjenkjennes av antistoffet som brukes i dette første trinn er de samme som eller tilstoppe de epitoper som gjenkjennes av antistoffet i den endelige flekken, ville man forvente at intensiteten av signalet til å være lavere enn hvis bare ett antistoff ble anvendt. Av denne grunn er det viktig å titrere antistoffet som brukes i det avsluttende trinn, slik at signalet fra en positivt farget celle er omtrent tostørrelsesordener høyere enn for et negativt farget celle. På denne måten, selv om intensiteten av signalet fra en positiv farging celle reduseres i celler som har gjennomgått isoleringsprosedyren, disse positive cellene vil fortsatt være godt adskilt fra de negative cellene på flowcytometri plottet. Hvis automatisk kompensering blir brukt, bør enkelt flekk kontrollprøvene fremstilles. Antistoffer som vi har generert er tilgjengelige for kjøp fra Memorial Sloan Cancer Center monocolonal antistoff anlegget. Vi har hatt den beste suksess med å bruke umerket monoklonalt IgG-antistoff. Vi har gjennomført protokollen med polyklonale VSG antistoffer, men polyklonale antistoffer kan tidvis gjenkjenne andre VSGs så monoklonale antistoffer er foretrukket. VSG identitet kan bestemmes ved sekvensering VSG cDNA generert fra amplifikasjon av konserverte sekvenser i VSG 3'UTR og skjøtes leder. Et spesielt antistoff kan bli testet for spesifisitet av tidligere isolert slåttceller som er VSG identitet er fastslått som nettopp beskrevet.
Protokollen presentert her har en rekke fordeler. Den kan brukes både for å isolere skiftere for senere analyse og for å kvantifisere antallet av omformerenheter i en gitt populasjon. Den kan også brukes til å isolere en spesiell variant av interesse, så lenge et antistoff er tilgjengelig for denne varianten. Det er også mulig å utføre denne protokollen ved hjelp av trypanosomer isolert fra dyr, forutsatt at de røde blodcellene blir eliminert ved anvendelse av anti-Ter119 belagte-magnetiske kuler i henhold til produsentens protokoll. Vi har ikke testet den nedre grense for antall trypanosomer er nødvendig, men vi har med hell utført ved hjelp av protokollen 2,5 - 10 millioner trypanosomer fra 250 ul blod. Endelig kan protokollen brukes i forbindelse med fluktuasjon analyse for å oppnå frekvensen for veksling i en gitt populasjon.
Isoleringsprosedyren er ganske fast, tar med 3 - 4 timer fra start til slutt, avhengig av antall prøver, og således er mer effektiv enn tidligere metoder som kreves før immunisering av mus eller spesialiserte stammer inneholdende medikamentresistensmarkører. Fremgangsmåten er begrenset av det faktum at antistoffer mot utgangs VSG er nødvendig, men. Uten slike reagenser, kan en alternativ metode, slik som VSG-seq være en mer passende valg for å måle hvilke VSGs blir uttrykt i en gitt populasjon 10.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Vi ønsker å takke George Cross for generelle råd om trypanosom biologi. Dette arbeidet ble også støttet av en Bill og Melinda Gates Foundation GCE tilskudd til DS, en NSF Graduate Research Fellowship (DGE-1325261) til MRM og en NIH / NIAID (tilskudd # AI085973) til FNP. Vi takker Galadriel Hovel-Miner for bruk av stammen inneholdende I-SCEI gen og gjenkjenningssete.
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
