

Introduction
原生动物寄生虫布氏锥虫 ,是影响人类(经冈比亚布氏锥虫和布氏锥虫罗得西亚),在整个撒哈拉以南非洲地区和动物(通过布氏锥虫布氏 )疾病的病原体。它是通过采采蝇矢量的唾液传递到哺乳动物宿主。人类和动物非洲锥虫引起流行地区严重的经济负担,而一些药物可或开发治疗疾病的两种。了解免疫逃逸机制是药物开发锥虫病的关键。茂密,变体表面糖蛋白(VSG)大衣覆盖寄生虫的抗原变异是主要手段之一,由T.布氏逃避哺乳动物抗体反应。将VSG基因的约2000个变体在锥虫基因组中存在,但只有一个在任何给定时间从一个转录〜15 EXPRES锡永的网站。所表达的VSG的'开关'可以通过复制VSG基因进入活性表达位点,或通过先前无声表达位点(1综述)的转录激活发生。
虽然大量的工作一直致力于理解抗原变异的T.布氏 ,影响开关频率的因素仍然知之甚少。迄今为止的研究受到阻碍的事实,而开关在体外随机发生,开关频率非常低,为1在约10 6个细胞2的量级。这使得难以测量给定因子增加或是否减小开关频率,因为切换是硬首先被检测到。在2009年之前,用于在给定的群体中分离切换方法是冗长且劳动密集的。这些措施包括通过传代对主要的VSG免疫小鼠锥虫,然后收获细胞一天后3,或者通过免疫4,5计数数百细胞。另一种策略依赖于选择耐药隔离切换6。因为在体外生长非洲锥虫通常由大量人口表达一个主要变型中,并表达交替变体切换的小得多的人口,我们指的是主要变体在整个本文作为主导,开始VSG。这样做,我们绝不想暗示这主要变种比在其他人群更大的变体健身。
在这里,我们描述了可以可靠地测量锥虫的在3在给定的群体中表达非主导VSG数量的方法 - 4小时。此方法是当一个人想要确定一个给定的基因操作或药物治疗是否增加群交换的细胞的数目为特别有用。相反,摆脱细胞群体表达的做minant,起动VSG通过药物或通过免疫学手段,这些细胞通过首先与耦合到抗体对占优势的VSG磁珠涂覆它们,然后在磁性柱中分离它们消除。开关人口然后在流过收集,并用荧光团标记的抗VSG抗体识别的污染物再次染色。量化是通过将在规定的绝对计数珠号每个样品,使珠细胞的比率可确定并用于量化在人口7切换的数目来实现的。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
注意:在整个过程中,有必要保持细胞在冰上。冷媒体也应在整个使用。
1.样本收获
- 利斯特增长427株血液锥虫至0.5的密度 - 100万/ ml。这是最好的开始与文化少数寄生虫。降速50×10 6细胞/样品在1500×g下10分钟。一定要留下1×10 6个细胞培养后用作阳性和阴性对照抗体。
注:此协议也可以从动物血液(详见讨论)隔离锥虫使用。 - 吸管或倒掉大部分的上清液(留下约750微升)。转移细胞到1.5ml微量离心管中。
- 旋转细胞在4℃下4分钟为5,200 XG在微量和上清。
2.磁标记
- 悬浮细胞在150微升培养基(HMI-9的Wi例如第血清)+基本抗VSG抗体在适当的稀释度。
注:抗VSG抗体是在内部提出,并在1:50的稀释使用。以使用不带有荧光团标记的初级抗VSG抗体是很重要的。 - 在冷室中在4℃下进行10分钟,8 - 使用涡旋适配器60以速度6涡流细胞。 1000微升冷HMI-9与血清 - 800清洗。
- 旋在在5200 xg离心4℃4分钟,吸出上清液。洗用1ml的HMI-9与血清并如上所述旋转。吸出上清液。重悬沉淀在100μl的HMI-9的血清。
- 添加磁性活化细胞的110微升分选(MSC)的微珠。使用抗小鼠,抗兔或抗生物素珠粒作为适合主抗VSG抗体。拌匀。涡旋细胞在冷室中在4℃下进行10分钟。
- 当细胞被涡旋,建立一个磁性激活分离柱/样本上分离磁铁。一定要有一个重新ceptacle(我们使用一个15毫升的离心管)的列下收集流过。在冷室中设置的装置。
- 加2ml HMI-9与血清素到列。列下吸代替15ml离心管,收集流过每个样品之后。
- 10分钟孵化步骤2.4后,加入800-1000微升冷HMI-9血清。旋在4℃4分钟为5,200 XG和上清以除去未结合的抗体。用1ml的HMI-9与血清和重复留下100μl的上清液洗。
- 轻弹离心管大力和目视检查管,以确保沉淀完全重悬。加入1ml的HMI-9与血清和移液器上下,以确保细胞是公重悬。
3.磁选
- 确保与上述列灌注。应用细胞列。与细胞中表达的非主导VS收集流通G. 1ml培养基的+锥虫通常需要6 - 7分钟通过该柱流过。
- 洗涤2次用1毫升的HMI-9与血清,并收集在同一个管流过如在步骤3.1。
- 分流通(3毫升)中分成两个1.5ml微量管中。
注意:这些可购买组合或半可用于分离RNA 等 - 从磁铁删除列。与血清添加3毫升HMI-9的柱和沉浸柱入15ml离心管,以获得表达的起始,主导VSG细胞。除去300μl的洗脱材料的供以后分析。置于冰上这些细胞。
- 用作阳性和阴性对照,采取从对照细胞的起始培养大约为1百万个细胞,在37℃生长(那些预计不频繁切换)并吸取它们放入一个1.5ml微量管中。
注意:阳性对照细胞将与荧光标记的抗体在步骤4.1染色。阴性对照组REMA在整个协议的其余部分未染色。 - 旋流过的样品和在5200 xg离心在4℃下4分钟阳性抗体对照样品并除去上清液。
注:洗脱细胞的另一300μl的等分试样可被用作用于抗VSG染色的阳性对照。
4.染色识别污染物和切换器
- 悬浮流通的样品和在100微升的HMI-9与血清+伯,荧光团标记,抗VSG在适当的稀释抗体的阳性对照细胞。
注意:标记的抗VSG抗体是在内部制成,以1:1的稀释比使用:1000。所述荧光团标记,抗VSG可从同一源如在MACS分离未标记的抗体step.There应在以下步骤3.5 2离心管3毫升流过的样品。 - 分析所有的流通过流式细胞术,结合粒料在两个离心管的同时重新SUS待决100微升HMI-9与血清+抗VSG抗体样品。拌匀。
- 涡旋细胞在冷室中在4℃下进行10分钟。添加800 - 1000微升冷HMI-9血清。在4℃下4分钟旋转样品在5200 xg离心并除去上清液以除去未结合的抗体。
- 洗涤细胞用1ml的HMI-9的血清。在4℃下4分钟旋转样品为5,200 XG和上清。在这个最后的旋转,从步骤3.4和阴性对照样品降速洗脱样本。
- 重悬所有流通和洗脱的样品中:148.5微升HMI-9与血清,25微升绝对算珠和1.5微升碘化丙啶染色溶液(175微升总)。悬浮阳性和阴性抗体对照样本中175微升HMI-9血清。一定要记录绝对算珠/ 25微升所使用的特定批次的数量。
- 贯穿流的所有样本仪和收集数据。作为染色PI的死细胞
- 计算开关电源采用流式细胞仪数据的频率。
注:有关如何计算的开关频率的说明,请参阅补充文件流式细胞仪Calculations.docx和表1。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
使用此处所描述的方法以证明VSG表达位点内的双链断裂的人口8增加切换的数量。在这里,我们显示已同样诱导产生在表达部位的双链断裂锥虫的群体代表性的结果。我们比较这些锥虫那些没有被诱导以产生双链断裂。 图1显示了代表性的流式细胞仪从流通从磁激活细胞分选柱收集的未诱导和诱导的细胞图。对于这个特定的实验中的主要起始VSG是VSG2。 图1A的左侧面板显示前向和侧向散射,它允许门绕绝对计数珠和细胞,用于显示正向和侧向散射签名是活细胞的特征性绘制。需要注意的是门中被吸引到包括更多的细胞,因为死细胞可以与PI染色被淘汰;然而,栅极必须保持对所有样品是相同的。 图1A的右侧面板只显示落在左侧面板中的“活”的大门内的细胞。该染色积极为PI(Q1和Q2)细胞是死细胞。即积极染色为主导VSG和消极的PI在三季度细胞污染物。这两个PI和Q4的主导VSG负染色细胞是已转活细胞。人们可以看到,在第三季度污染物的百分比是那些双链断裂已诱导细胞低。然而,在第三季度百分比不能用来推断是否有一个给定的群体更切换器。这只是在Q4 的细胞数的比率进行比较,以收集一个可以计算的开关频率的小珠的数量如表1所示从FLO得到的数字W¯¯流式细胞仪的情节和方法来计算这些人群的百分比切换。这些计算是在3个生物样品进行,以获得在图1B中显示的观察开关频率。 在体外随机切换的水平相当低,如这里在平均9.53×10 -5计算,而在1诱导断裂的结果-已切换到非主导VSG 6×10 -3的细胞。
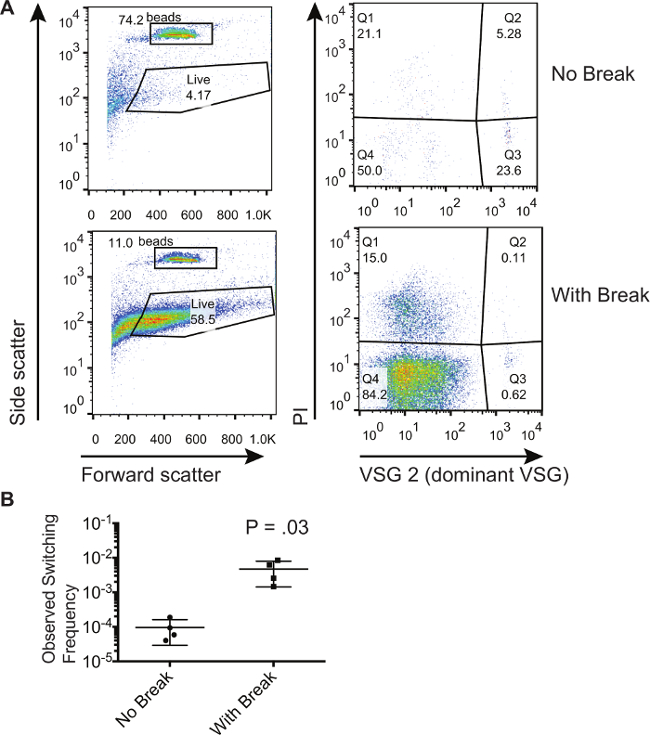
图1. 隔离和交换锥虫种群量化。 (A)流式细胞表示在下面的非切换的上一个磁激活细胞分选列选择流通级分锥虫的级分。与诱导的I-SCEI双链断裂细胞群相比较的细胞群没有诱导冰川AK。(B)的计算值的开关频率对于具有或没有被诱导与I-SCEI双链断裂细胞。在图上的数字代表一个双尾,非配对t检验的结果。误差线代表SD。 请点击此处查看该图的放大版本。
补充文件1.流式细胞仪计算。 请点击此处下载此文件。

表1.开关频率的计算。 请点击这里查看此表的放大版本。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
相对于实验技术,该协议的最关键的部件是保持所有样品冷。锥虫非常迅速内在化结合到其表面9抗体,但该方法是运动相关的,只要将细胞保持在4℃下不影响测定。所有样品应始终保持在冰上,并移液应该迅速完成,以减少暴露于25℃的实验室环境中。冷的HMI-9与血清应可在实验的开始,并应在整个保持在冰上。通过在柱运行它们的锥虫的隔离必须在冷室中进行的,因为它需要5 - 7分钟通过该柱以运行一个1ml样品。如果有比可用磁分离单元更多的样品,磁隔离可以以序列为不被积极分离样品保持在冰上进行,如长。
这也是至关重要的样品是非常充分混合自动对焦之三在与微珠和前隔离的磁铁孵化最后一次洗涤。这可通过将离心管内或光涡旋的侵略性轻弹来完成。细胞的无丸粒或团块应加入样品到分离柱,其可通过观察光镜水平进行检查之前是可见的。如果团块存在时,污染的细胞的流过样品中表达显性VSG的水平通常比期望的要高得多。当在协议结束时的样品中切换的数量进行计算,它是否被用于流式细胞仪分析所有的流过,考虑到是很重要的,或者是否使用一半。如果只使用一半,计算切换器的总数应乘以2。
当在洗涤步骤除去上清液,这是没有必要对移液器关闭每一滴,并且通常为15 - 25微升是左周围细胞的沉淀。这是在染色和洗涤的最后阶段特别关键,因为在这一点上粒料流过样品中很少见到,因为有这么几个切换细胞。
而最终反VSG污渍通常与纯化的,荧光团标记的抗VSG抗体进行之前,磁隔离的第一抗VSG染色可与纯化的抗体或血清进行。我们也使用来自表达抗VSG抗体的杂交瘤衍生的生物反应器的上清液。无论如何,用于初始污点的材料必须被滴定以确保或不表达的主导,起始VSG细胞良好的分离。
它是典型的在流通细胞群与表达显性VSG细胞污染。尽管这种污染应该是最小的,在我们的经验,这是罕见的人口是完全没有细胞的表达ING占主导地位的VSG。同样重要的是要注意,已经通过分离过程去细胞不与抗VSG抗体作为那些没有通过程序被作为明亮染色。我们假设两件事情可以解释这种差异。首先,纺丝锥虫导致VSG的脱落,而且还有所涉及的分离步骤在许多旋转。这预期会降低信号的强度的最终反VSG污点。第二个是,该协议的第一步骤涉及染色与伯抗VSG抗体。如果在该第一步骤中使用的抗体识别的表位是相同或阻塞通过在最终的染色的抗体识别的表位,人们会期望的信号的强度为比如果只有一个抗体被用于低。由于这个原因,以滴定是在最后步骤中使用的抗体,例如,从一个染色阳性细胞的信号是大致二是重要幅度比一个负染细胞更高的数量级。这样一来,即使信号从一个染色阳性细胞的强度在已经经历了分离过程的细胞被减小,这些阳性细胞将仍然很好从在流式细胞情节的阴性细胞分离。如果正在使用自动补偿,单染色的对照样品,应制备。我们已经生成的抗体可从纪念斯隆癌症中心的单克隆抗体设施购买。我们曾使用未标记的单克隆IgG抗体的最佳成功。我们已经进行了该协议使用多克隆VSG抗体,但多克隆抗体可以偶尔认识到其它的VSG所以单克隆抗体是优选的。 VSG同一性可以通过从VSG 3'UTR和剪接前导保守序列的扩增产生测序VSG的cDNA来确定。特定抗体可以特异性被测试的先前孤立切换已如刚才所述确定细胞谁的VSG身份。
这里介绍的协议具有许多优点。它可以用来既以分离供以后分析切换器和量化给定群体切换的数量。它也可以用于分离感兴趣的特定变体,只要抗体是可用于该变体。另外,也可以进行使用从动物中分离锥虫这个协议中,前提是红血细胞根据制造商的方案使用抗TER119涂覆磁性珠粒消除。我们没有测试所需锥虫数量的下限,但我们已经成功地进行了使用2.5协议年 - 1000万元锥虫从250微升血液。最后,该协议可以结合使用与波动分析以获得在给定种群的开关频率。
分离过程是相当˚FAST,服用3 - 4小时,从开始到取决于样品的数量完成,从而比以前的所需含药物抗性标记鼠或专门菌株的现有免疫方法更有效。该方法是由针对起始VSG抗体然而需要,这一事实的限制。如果没有这样的试剂,诸如VSG-SEQ的另一种方法可能是更合适的选择,以了解它们在一个给定的群体10被表达的VSG。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
我们要感谢乔治十字对锥虫生物学的一般建议。这项工作也由比尔和梅琳达·盖茨基金会GCE授予DS,一个国家科学基金会研究生研究奖学金(DGE-1325261),以MRM和NIH / NIAID(批#AI085973),以支持FNP。我们感谢凯兰崔尔伽格尔营地,矿工使用含I-SCEI基因识别位点的应变。
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
