

Introduction
Den protozo parasit, Trypanosoma brucei, är en orsakande medlet av sjukdomar som drabbar människor (via Trypanosoma brucei gambiense och Trypanosoma brucei rhodesiense) och djur (via Trypanosoma brucei brucei) hela Afrika söder om Sahara. Den överförs till däggdjursvärden genom saliven av tsetseflugan vektorn. Både människor och djur afrikansk sömnsjuka orsaka en allvarlig ekonomisk börda i endemiska områden, och några läkemedel är tillgängliga eller under utveckling för behandling av antingen sjukdom. Att förstå mekanismerna för immunundan är avgörande för utveckling av läkemedel för trypanosomiasis. Antigenvariation hos den täta, variant ytglykoprotein (VSG) päls som täcker parasiten är en av de primära medel genom vilka T. brucei undviker däggdjursantikroppssvar. Ca 2000 varianter av VSG-genen existerar i trypanosom-genomet, men endast en transkriberas vid varje given tidpunkt från ett på ~ 15 expressions platser. "Omkoppling" av det uttryckta VSG kan ske antingen genom att kopiera en VSG gen i aktivt uttryck webbplats eller genom transkriptionsaktivering av en tidigare tyst uttryck webbplats (översikt i ett).
Även mycket arbete har lagts ner på att förstå antigenvariation i T. brucei, är de faktorer som påverkar switchfrekvensen fortfarande dåligt kända. Studier hittills har hindrats av det faktum att medan omkoppling sker stokastiskt in vitro, switchfrekvenser är mycket låga, i storleksordningen en på cirka 10 6 celler 2. Detta gör det svårt att mäta om en viss faktor ökar eller minskar switchfrekvens, eftersom omkopplings är svåra att upptäcka i första hand. Före 2009, metoder för isolering paneler i en given population var långa och arbetskrävande. Dessa inkluderade Pass trypanosomer genom möss som immuniserats mot den dominerande VSG och sedan skörda celleren dag senare tre, eller räkna hundratals celler genom immunofluorescens 4,5. En annan strategi är beroende av val för läkemedelsresistens för att isolera paneler 6. Eftersom afrikanska trypanosomer odlas in vitro är typiskt består av en stor population som uttrycker en större variant, och en mycket mindre population av switchar uttrycker alternativa varianter, hänvisar vi till den stora varianten hela detta dokument som den dominerande, börjar VSG. På så sätt har vi på något sätt vill antyda att denna stora varianten har större kondition än andra varianter i befolkningen.
Här beskriver vi en metod som på ett tillförlitligt sätt kan mäta antalet trypanosomer som uttrycker ett icke-dominanta VSG i en given population i 3-4 h. Denna metod är särskilt användbar för när man vill fastställa huruvida en given genetisk manipulation eller läkemedelsbehandling ökar antalet omkopplade celler i en population. I stället för att befria populationen av celler som uttrycker dominant start VSG genom droger eller genom immunologiska metoder, är dessa celler elimineras genom att först belägga dem med magnetiska kulor kopplade till antikroppar mot det dominerande VSG och sedan isolera dem på en magnetisk kolonn. Den omkopplade populationen uppsamlas sedan i genomflödet och färgades igen med en fluorofor märkt anti-VSG-antikropp för att identifiera kontaminanter. Kvantifiering åstadkoms genom att tillsätta ett definierat antal absoluta räknings pärlor till varje prov så att förhållandet av pärlor till celler kan bestämmas och användas för att kvantifiera antalet switchar i populationen 7.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
OBS: Under hela förfarandet, är det nödvändigt att hålla cellerna på is. Kalla medier bör också användas i hela.
1. Prov Skörd
- Växa Lister 427-stammen blodomloppet trypanosomer till en densitet av 0,5 - 1 miljon / ml. Det är bäst att börja kulturer med ett litet antal parasiter. Spinn ner 50 x 10 6 celler / prov i 10 minuter vid 1500 x g. Var noga med att lämna 1 x 10 6 celler i odling under senare användas som positiva och negativa kontroller antikroppar.
OBS: Detta protokoll kan också användas på trypanosomer som isolerats från djurblod (se diskussion för detaljer). - Pipett eller häll bort återstoden av supernatanten (lämna ca 750 | j, l). Överföra celler till ett 1,5 ml mikrocentrifugrör.
- Snurra cellerna vid 4 ° C under 4 min vid 5200 xg i en mikrocentrifug och avlägsna supernatanten.
2. magnetisk märkning
- Resuspendera celler i 150 pl odlingsmedium (HMI-9 with serum till exempel) + primär anti-VSG antikropp vid lämplig utspädning.
OBS: Den anti-VSG antikropp görs i hus och användes vid en spädning av 1:50. Det är viktigt att använda en primär anti-VSG antikropp som inte är märkt med en fluorofor. - Virvelceller med användning av en vortex-adapter-60 vid hastighet 6-8 i ett kallt rum vid 4 ° C under 10 min. Tvätta med 800 - 1000 | j, l kall HMI-9 med serum.
- Centrifugering vid 4 ° C under 4 min vid 5200 xg och aspirera supernatanten. Tvätta med en ml HMI-9 med serum och snurra som ovan. Aspirera supernatanten. Återsuspendera pelleten i 100 | il av HMI-9 med serum.
- Lägg 110 pl magnetiskt aktiverad cellsortering (MSC) mikrokulor. Använda anti-mus, anti-kanin eller anti-biotin pärlor som är lämpligt för den primära anti-VSG antikropp. Blanda väl. Virvelceller i ett kallt rum vid 4 ° C under 10 min.
- Medan celler vortexa, inrätta en magnetisk-aktiverad separationskolonn / prov på en separation magnet. Var noga med att ha en receptacle (vi använder ett 15 ml centrifugrör) i kolumnen för att samla genomflödet. Sätt upp utrustningen i ett kallt rum.
- Tillsätt 2 ml av HMI-9 med serum för att prima kolonn. Efter grundning gör ett 15 ml centrifugrör under kolumnen för att samla genomflöde från varje prov.
- Efter 10 minuters inkubation i steg 2,4, tillsätt 800-1000 l kall HMI-9 med serum. Spin vid 4 ° C under 4 min vid 5200 xg och avlägsna supernatanten för att avlägsna obunden antikropp. Tvätta med 1 ml av HMI-9 med serum och upprepa lämnar 100 | il av den överstående vätskan.
- Flick centrifugröret kraftigt och visuellt inspektera röret för att se till att pelleten är helt återsuspenderas. Tillsätt 1 ml HMI-9 med serum och pipettera upp och ner för att se till att cellerna är väl återsuspenderas.
3. Magnetisk separation
- Se till att kolumnerna är primade såsom beskrivits ovan. Tillämpa celler till kolumn. Samla genomströmning med celler som uttrycker den icke-dominanta VSG. 1 ml medium + trypanosomer tar normalt 6-7 min för att strömma genom kolonnen.
- Tvätta 2x med 1 ml av HMI-9 med serum och samla genomströmning i samma rör som i steg 3,1.
- Divide genomströmning (3 ml) i två 1,5 ml mikrocentrifugrör.
OBS: Dessa kan kombineras senare eller halv kan användas för att isolera RNA, etc. - Ta bort kolumn från magneten. Tillsätt 3 ml av HMI-9 med serum till kolumn och störta kolumn till en 15 ml centrifugrör för erhållande av celler som uttrycker utgångs, dominant VSG. Avlägsna 300 pl eluerade materialet för senare analys. Håll dessa celler på is.
- För användning som en positiv och en negativ kontroll, ta ungefär 1 miljon celler från startkultur av kontrollceller växer vid 37 ° C (de som inte förväntas byta ofta) och pipettera dem i en 1,5 ml mikrocentrifugrör.
OBS: Positiva kontrollceller kommer att färgas med fluorofor-märkta antikroppen i steg 4,1. Negativa kontrollceller remai ofärgade under resten av protokollet. - Spin genomströmnings prover och positiv kontrollantikropp provet vid 4 ° C under 4 min vid 5200 xg och avlägsna supernatanten.
OBS: En annan 300 pl alikvot av eluerade celler kan användas som en positiv kontroll för anti-VSG färgning.
4. Färgning att identifiera miljöskadliga ämnen och Switchers
- Resuspendera genomflödes prover och positiva kontrollceller i 100 | il av HMI-9 med serum + primär, fluorofor-märkt, anti-VSG-antikropp vid lämplig utspädning.
OBS: Den märkta anti-VSG antikropp görs i hus och användes vid en spädning av 1: 1000. Fluoroforen-märkta, kan anti-VSG vara från samma källa som den omärkta antikroppen i MACS separation step.There bör vara 3 ml genomströmnings prov i två mikrocentrifugrör efter steg 3,5. - För att analysera alla genomströmning genom flödescytometri, kombinera pellets i de två mikrocentrifugrör medan re-susavvaktan på prov i 100 pl av HMI-9 med serum + anti-VSG antikropp. Blanda väl.
- Virvelceller i ett kallt rum vid 4 ° C under 10 min. Lägg 800 - 1000 | j, l kall HMI-9 med serum. Spin prover vid 4 ° C under 4 min vid 5200 xg och avlägsna supernatanten för att avlägsna obunden antikropp.
- Tvätta cellerna med 1 ml HMI-9 med serum. Spin prover vid 4 ° C under 4 min vid 5200 xg och avlägsna supernatanten. Under denna sista spinn, spinn ner de eluerade-prover från steg 3,4 och den negativa kontrollprovet.
- Resuspendera alla genomströmning och eluerade prover: 148,5 pl HMI-9 med serum, 25 il absolut räkna pärlor och 1,5 l propidiumjodidfärgning lösning (175 ul totalt). Resuspendera positiva och negativa kontrollantikroppsprover i 175 pl HMI-9 med serum. Var noga med att registrera antalet absoluta räkna pärlor / 25 ^ för särskilt parti används.
- Kör alla prover genom en flödescytometer och samla in data. Döda celler färgas som PI
- Beräkna byta frekvenser med användning av flödescytometri data.
OBS: För instruktioner om hur man beräknar omkopplingsfrekvenser, hänvisas till kompletterande fil Flödescytometri Calculations.docx och tabell 1.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Den metod som beskrivs här användes för att visa att dubbelsträngbrott inom VSG uttryck platsen ökade antalet switchar i en population 8. Här visar vi representativa resultat från en population av trypanosomer som har liknande lockats att generera en dubbelsträngbrott vid uttryck platsen. Vi jämför dessa trypanosomer till dem som inte har förmåtts att generera en dubbelsträngbrott. Figur 1 visar representativa flödescytometri tomter från un-inducerade och inducerade celler som samlats i genomflödet från magnet aktiverad cellsortering kolumn. För detta speciella experiment dominerande start VSG var VSG2. Den vänstra paneler av Figur 1A visar framåt och sidospridning, vilket gör att grindarna dras runt de absoluta räkna pärlor och celler som visar en framåt och sidospridning signatur som är kännetecknande för levande celler. Notera att grindar kandras till att omfatta fler celler eftersom döda celler kan elimineras med PI-färgning; måste emellertid grinden vara densamma för alla prover. De högra panelerna i figur 1A visar endast de celler som faller inom "Live" gate på vänstra panelerna. Celler som färgas positivt för PI (Q1 och Q2) är döda celler. Celler som färgas positivt för den dominerande VSG och negativt för PI i Q3 är föroreningar. Celler som färgas negativt för både PI och den dominerande VSG i Q4 är levande celler som har bytt. Man kan konstatera att den procent av föroreningar i Q3 är lägre för de celler där en dubbelsträngbrott har inducerats. Däremot kan de procentsatser i Q3 inte användas för att dra slutsatsen huruvida det finns fler paneler i en given population. Det är endast genom att jämföra förhållandet av antalet celler i Q4 till det antal pärlor uppsamlades att man kan beräkna omkopplingsfrekvenser. Tabell 1 visar de siffror som erhållits från flow cytometry tomter och metoden för att beräkna procent switchar i dessa populationer. Dessa beräkningar utfördes på 3 biologiska prover för att få de observerade switchfrekvenser som visas i figur 1B. Nivån av stokastiska omkoppling in vitro är ganska låg, beräknat här i genomsnitt 9,53 x 10 -5, medan framkalla ett avbrott resulterar i 1 - 6 x 10 -3 celler som har bytt till den icke-dominanta VSG.
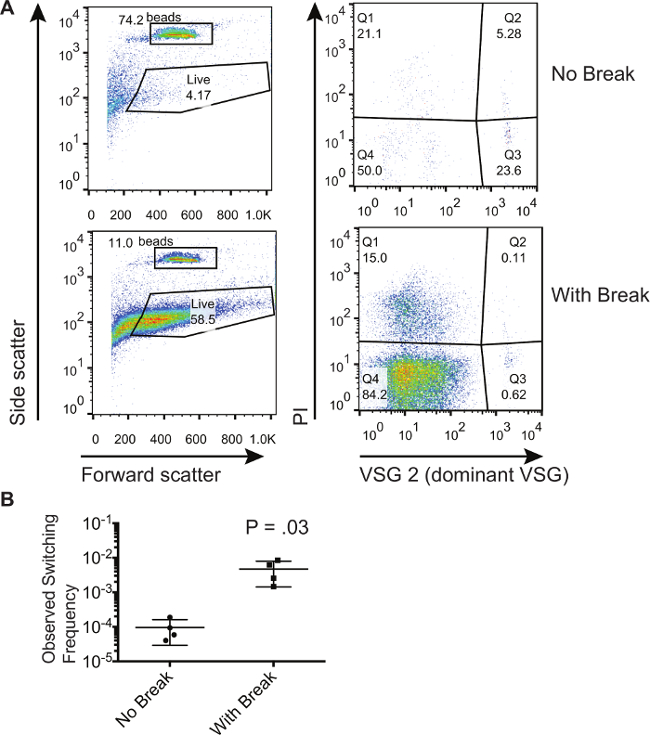
Figur 1. Isolering och kvantifiering av Switched trypanosom Populationer. (A) Flödescytometri visar fraktioner av trypanosomer i genomflödesfraktionen efter urval av icke-switchar på en magnetisk aktiverad cellsortering kolonn. En population av celler med en inducerad I-SCEI dubbelsträngbrott jämförs med en population av celler utan en inducerad break. (B) Beräknad kopplingsfrekvenser för celler som har eller inte har inducerats med en I-SCEI dubbelsträngade paus. Numret på tomten visar resultaten av en två-tailed, oparade t-test. Felstaplar representerar SD. Klicka här för att se en större version av denna siffra.
Kompletterande fil 1. Flödescytometri Beräkningar. Klicka här för att ladda ner filen.

Tabell 1. Beräkning av switchfrekvenser. Klicka här för att se en större version av denna tabell.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
När det gäller experimentell teknik, är den mest kritiska komponenten i protokollet att hålla alla prover kallt. Trypanosomer mycket snabbt internalisera antikropp bunden till deras yta 9, men denna process är motilitet beroende och inte påverkar analysen, så länge som cellerna hålls vid 4 ° C. Alla prover bör alltid hållas på is, och pipettering bör göras snabbt för att minimera exponeringen för 25 ° C labbmiljö. Cold HMI-9 med serum bör vara tillgängliga i början av experimentet och bör hållas på is hela tiden. Isolering av trypanosomer genom att köra dem över kolonnen måste göras i ett kallt rum, som det tar 5 - 7 minuter att köra en 1 ml prov genom kolonnen. Om det finns fler prover än magnetisk separations enheter tillgängliga, kan magnetisk isolering göras i ordning, så länge proverna inte aktivt separerade hålls på is.
Det är också viktigt att proverna vara mycket väl blandad after den sista tvättningen efter inkubation med mikropärlorna och före isolering på magneten. Detta kan åstadkommas genom aggressiv snärta av centrifugröret eller lätt virvelbildning. Ingen pellet eller klumpar av celler bör vara synligt före tillsats av provet till separationskolonnen, vilket kan kontrolleras genom att titta på ljusmikroskop-nivå. Om klumpar är närvarande, är vanligtvis nivån av föroreningar i prov av celler genomströmnings uttrycker dominerande VSG mycket högre än önskvärt. Vid beräkningen för antalet switchar i provet vid slutet av protokollet, är det viktigt att ta hänsyn till om alla genomflödet användes för flödescytometrisk analys, eller om hälften användes. Om bara hälften användes, bör det totala antalet switchar beräknas multipliceras med två.
När du tar bort supernatanten under tvättstegen, är det inte nödvändigt att pipett bort varenda droppe, och typiskt 15 - 25 ^ ärvänster kring pellet av celler. Detta är särskilt viktigt under slutskedet av färgning och tvätt, eftersom det vid denna punkt pellets är sällan sett i genomströmnings prover eftersom det finns så få bytte celler.
Medan det slutliga anti-VSG fläcken utförs typiskt med renat, fluorofor-märkt anti-VSG-antikropp, kan det första anti-VSG fläck före magnetisk isolering göras med renad antikropp eller serum. Vi har också använt bioreaktor supernatant härrör från hybridom som uttrycker anti-VSG antikroppar. Oavsett, måste materialet som används för den första fläcken titreras för att säkerställa god separation av celler som gör eller inte uttrycker den dominerande, börjar VSG.
Det är typiskt för den population av celler i genomströmning vara kontaminerade med celler som uttrycker den dominerande VSG. Även om denna förorening bör vara minimal, enligt vår erfarenhet är det ovanligt för befolkningen att vara helt utan celler uttryckering den dominerande VSG. Det är också viktigt att notera att celler som har gått igenom isoleringsförfarandet inte fläcken som ljust med anti-VSG antikropp som de som inte har gått igenom proceduren. Vår hypotes är att två saker skulle kunna förklara denna skillnad. Den första är att snurra trypanosomer leder till utsöndring av VSG, och det finns många snurrar inblandade i isoleringsförfarandet. Detta skulle förväntas minska intensiteten hos signalen för den sista anti-VSG fläcken. Det andra är att det första steget i protokollet innebär färgning med en primär anti-VSG antikropp. Om de epitoper som igenkännes av den antikropp som används i detta första steg är desamma som eller ockludera de epitoper som känns igen av antikroppen i den slutliga fläcken, skulle man förvänta sig intensiteten av signalen att vara lägre än om enbart en antikropp användes. Av denna anledning är det viktigt att titrera antikroppen som används i det slutliga steget, så att signalen från en positivt färgad cell är ungefär tvåtiopotenser högre än för ett negativt färgade cell. På detta sätt, även om intensiteten av signalen från en positivt färgning cell minskas i celler som har genomgått isoleringsförfarandet, dessa positiva celler kommer fortfarande att vara väl separerade från de negativa cellerna på flödescytometri tomt. Om auto-ersättning används, bör enstaka fläck kontrollprover framställas. Antikroppar som vi har genererat finns att köpa från Memorial Sloan Cancer Center monocolonal antikropps anläggning. Vi har haft den bästa framgång med hjälp av omärkt monoklonal IgG-antikropp. Vi har genomfört protokollet med polyklonala VSG antikroppar, men polyklonala antikroppar kan ibland känna igen andra VSGs så monoklonala antikroppar är att föredra. VSG identitet kan bestämmas genom sekvense VSG cDNA genererad från amplifiering av konserverade sekvenser i VSG 3'UTR och den skarvade ledare. En speciell antikropp kan testas för specificitet på tidigare isolerade omkopplasceller som finns i VSG identitet har fastställts som just beskrivits.
Protokollet som presenteras här har ett antal fördelar. Den kan användas både för att isolera switchar för senare analys och för att kvantifiera antalet switchar i en given population. Den också kan användas för att isolera en speciell variant av intresse, så länge som en antikropp är tillgänglig för den varianten. Det är även möjligt att utföra detta protokoll med användning av trypanosomer isolerade ur djur, förutsatt att röda blodkroppar elimineras med användning av anti-Ter119 belagt-magnetiska pärlor i enlighet med tillverkarens protokoll. Vi har inte testat den lägre gränsen för antalet trypanosomer krävs, men vi har framgångsrikt genomfört protokoll med 2,5 - 10 miljoner trypanosomer från 250 pl blod. Slutligen kan protokollet användas i samband med fluktuation analys för att erhålla frekvensen för att växla i en given population.
Isoleringsförfarandet är ganska fast, tar 3-4 timmar från start till mål, beroende på antalet prover, och således är mer effektiv än tidigare metoder som krävs före immunisering av möss eller specialiserade stammar innehållande läkemedelsresistensmarkörer. Metoden är begränsad av det faktum att antikroppar mot utgångs VSG krävs dock. Utan sådana reagens, kan en alternativ metod som VSG-punkter vara ett mer lämpligt val för att mäta vilka VSGs är uttrycks i en given population 10.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Vi vill tacka för George korset för allmänna råd om trypanosom biologi. Detta arbete stöddes också av en Bill och Melinda Gates Foundation GCE bidrag till DS, en NSF Graduate Research Fellowship (DGE-1.325.261) till MRM och en NIH / NIAID (bidrags # AI085973) till FNP. Vi tackar Galadriel ruckel-Miner för användning av stam innehållande I-SCEI genen och igenkänningssäte.
Materials
| Name | Company | Catalog Number | Comments |
| unlabeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| fluorophore labeled anti-VSG antibody | n/a | n/a | VSG antibody is made in house |
| HMI-9 media | n/a | n/a | HMI-9 is made in house |
| Propidium Iodide | BD Pharmingen | 556463 | |
| CountBright absolute counting beads | Thermofisher Scientific | C36950 | |
| LD columns | Miltenyi Biotech | 130-042-901 | |
| MACS magnet | Miltenyi Biotech | 130-042-303 | |
| MACS magnetic separator | Miltenyi Biotech | 130-042-302 | |
| vortex adapter-60 | ThermoFisher scientific | AM10014 | |
| flow cytometer | Coulter | n/a | |
| flow cytometer analysis software | FloJo | n/a |
References
- Horn, D. Antigenic variation in African trypanosomes. Molecular & Biochemical Parasitology. 195 (2), 123-129 (2014).
- Lamont, G. S., Tucker, R. S., Cross, G. A. Analysis of antigen switching rates in Trypanosoma brucei. Parasitology. 92 (Pt 2), 355-367 (1986).
- Rudenko, G., McCulloch, R., Dirks-Mulder, A., Borst, P. Telomere exchange can be an important mechanism of variant surface glycoprotein gene switching in Trypanosoma brucei. Mol Biochem Parasitol. 80 (1), 65-75 (1996).
- Turner, C. M., Barry, J. D. High frequency of antigenic variation in Trypanosoma brucei rhodesiense infections. Parasitology. 99 (Pt 1), 67-75 (1989).
- Miller, E. N., Turner, M. J. Analysis of antigenic types appearing in first relapse populations of clones of Trypanosoma brucei. Parasitology. 82 (1), 63-80 (1981).
- Horn, D., Cross, G. A. Analysis of Trypanosoma brucei vsg expression site switching in vitro. Mol Biochem Parasitol. 84 (2), 189-201 (1997).
- Figueiredo, L. M., Janzen, C. J., Cross, G. A. M. A histone methyltransferase modulates antigenic variation in African trypanosomes. PLoS Biol. 6 (7), e161 (2008).
- Boothroyd, C. E., Dreesen, O., et al. A yeast-endonuclease-generated DNA break induces antigenic switching in Trypanosoma brucei. Nature. 459 (7244), 278-281 (2009).
- Engstler, M., Pfohl, T., et al. Hydrodynamic Flow-Mediated Protein Sorting on the Cell Surface of Trypanosomes. Cell. 131 (3), 505-515 (2007).
- Mugnier, M. R., Cross, G. A. M., Papavasiliou, F. N. The in vivo dynamics of antigenic variation in Trypanosoma brucei. Science. 347 (6229), 1470-1473 (2015).
