

Summary
여기에 제시된 것은 높은 정밀도와 분류학적 분해능으로 수생 약탈 진핵생물의 방목 속도를 정량화하는 단일 세포, epifluorescence 현미경 기반 기술에 대한 프로토콜입니다.
Abstract
포식과 그 효과와 같은 영양 상호 작용을 해명하는 것은 생태학에 있는 많은 연구원을 위한 빈번한 작업입니다. 미생물 공동체의 연구는 많은 한계를 가지고 있으며, 육식 동물, 먹이 및 약탈 비율을 결정하는 것은 종종 어렵습니다. 여기에 제시된 것은 형광으로 표지된 먹이를 트레이서로 추가하여 수생 육식 진핵생물의 방목 속도의 안정적인 정량화와 더 높은 영양 수준으로의 영양 전달 추정을 가능하게 하는 최적화된 방법입니다.
Introduction
이종 영양 성 원생 체는 수생 시스템의 주요 생물학적 구성 요소이며 플랑크톤 바이오 매스1,2,3의상당한 부분을 차지합니다. 그들의 풍부, 다양성 및 활동을 통제하는 요인은 생물 지구 화학 사이클링에 있는 그들의 역할을 이해하는 위해 중요합니다 (즉, 유기 탄소 및 그밖 양분의 운명 및 원핵생물에서 더 높은 영양 수준에 에너지의 교류). 원생 동물 방목은 이러한 중요한 요소 중 하나입니다. 이종 영양 나노 깃 형성 및 섬모의 박취는 원핵 생물 풍부, 지역 사회 기능, 구조, 다양성, 심지어 세포 형태와 특정 세균 그룹의 성장 속도에 대한 강력한 하향식 제어를 부과4, 5,6. 일부 시스템에서는, protists세균 사망의 주요 원인으로 봉사6,7.
지금 얼마 동안 이용된 원생 동물 세균을 평가하기 위하여 이용된 표준 접근은, 먹이 유사체 및 epifluorescence 현미경 검사법으로 형광으로 표지된 박테리아 (FLB)의 사용을 관련시킵니다. 세포 특이적 관세포 비율은 선택된 시간 과정8에걸쳐 프로티스탄 식품 액구에서 표지된 먹이 입자의 수를 정량화함으로써 결정될 수 있다. 이 접근 방식에는 몇 가지 장점이 있습니다. 트레이서자연 육식 동물과 먹이 조립과 자연 샘플에 추가됩니다. 인큐베이션 전에 최소 샘플 조작, 추가된 FLB 추적에 의한 최소 샘플 변경, 인큐베이션 시간이 짧아 상태 조건에서 가까운 조건에서 얻은 음량 결과를 보장합니다. 또는, 세균성 프로티스트 또는 동물플랑크톤(예: 해양 해양 시스템)이 적은 환경에서는 소량(2%-3% 추적자)으로 시료에 첨가된 FLB의 소실률이 장기간 유세포측정을 통해 검출할 수 있습니다(12-24시간). 인큐베이션 실험. 그런 다음 시작 점과 끝점의 FLB 수(모든 세균의 영향을 통합)는 유세포 측정에 의해 정량화됩니다(자세한 내용은 이전 간행물9참조). 그러나, 이러한 파라미터는 특정 프로티스탄 및 동물플랑크톤 방목기 그룹 또는 종에 직접적으로 기인할 수 없는 총 집계 된 세균 비율을 나타냅니다.
전반적으로, 수생 환경에서 프로티스탄 종- 또는 형태형 특이적 세균 사망률을 정확하고 생태학적 의미로 정량화하는 것은 어려울 수 있다. 일부 프로티스트는 선택적 방목자이며, 추가된 FLB 트레이서의 크기 및 세포 형상은 먹이 섭취의 자연적인 비율을 왜곡할 수 있다10,11. 더욱이, 프로티스탄 활동 및 물질 대사는 매우 온도에 민감한12; 따라서 추가된 FLB 추적자의 양은 각 개별 샘플 유형에 대해 신중하게 조작되어야 합니다(박테리아의 자연적인 풍부성, 크기 및 형태와 세균의 일반적인 유형뿐만 아니라 온도에도 따라). 대부분의 연구는 대량 프로 티 스탄 방목 활동에 초점; 그러나, 특정 프로티스탄 종의 세균은 종종 훨씬 더 높은 정보 값을 보유하고 바람직 할 수있다. 이 경우 샘플에 존재하는 프로티스트 종에 대한 분류학적 지식과 행동에 대한 이해가 필요합니다. 따라서, 시간과 노동의 상당한 금액은 특정 프로티스탄 그룹 또는 종에 기인 하는 세균의 종 특정 비율에 대 한 사운드 결과 얻을 필요.
이러한 어려움에도 불구하고,이 방법은 자연 설정에서 프로티스탄 세균을 평가하기 위해 현재 사용할 수있는 가장 적합한 도구 남아있다. 여기에 제시된 수생 미생물 생태 학 연구에서 FLB를 추적자로 사용하기위한 포괄적이고 따르기 쉬운 방법입니다. 접근법의 모든 문제적 측면을 설명하고 개선 된 워크플로우를 설명하며, 대조적인 환경에서 두 가지 실험과 함께 섬모 종을 대조적으로 예로 설명합니다.
첫 번째 사례 연구는 체코의 중불위성 의 르지모프(Římov) 저수지에서 에필림네틱 환경에서 수행되었으며, 이는 대부분의 수면담수 체에 필적하는 방목지와 세균풍부를 보여줍니다(5,7참조). 두 번째 사례 연구는 수생 육식 식물 Utricularia 반사의트랩 내부 매우 특정 환경에서 실시되었다, 이는 모두 방목 mixotrophic 섬모의 매우 높은 숫자를 호스팅(테트라 히메나 utriculariae) 및 세균 성 세포. 두 샘플 유형 모두에서 세포 별 방목 속도 및 세균 서 주식의 계산이 표시됩니다. 결과의 생태 해석의 범위는 그 때 토론되고, 가능한 후속 연구 결과의 보기는 마침내 건의됩니다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 샘플 수집
-
저수지 물 샘플 의 수집: 첫 번째 사례 연구 (Exp I; 체질 육식 동물과 먹이 풍부 시스템에서 낮은 자연)
- 적절한 깊이에서 원하는 위치에서 물 샘플을 수집합니다. 실험실로 운반하는 동안 시료를 온도 조절 식 쿨러에 체재하여 (온도 충격을 피하며, 프로티스트의 섭취 율은 온도에 따라 달라지도록 주의해야 합니다).
참고 : 우리의 샘플링은 메소 - 영양 성 협곡 모양의 르지 모프 저수지 (사우스 보헤미아, 볼륨 34.5 x 106m 3,최대 깊이 43 m, 평균 체류 시간 100 일, dimictic)에서 수행되었습니다. 샘플링 사이트는 댐에 가까운 30m 깊이에 위치했다. 0.5 m 깊이에서 혼합 된 샘플을 2 l 프리딩거 샘플러에 의해 수집하였다. - 가능한 한 빨리 섹션 1.2를 계속합니다.
- 적절한 깊이에서 원하는 위치에서 물 샘플을 수집합니다. 실험실로 운반하는 동안 시료를 온도 조절 식 쿨러에 체재하여 (온도 충격을 피하며, 프로티스트의 섭취 율은 온도에 따라 달라지도록 주의해야 합니다).
-
육식 성 Utricularia 반사 요법 식물의 함정에서 유체의 컬렉션 : 두 번째 사례 연구 (Exp II; 높은 육식 동물과 먹이 풍부 시스템, 매우 작은 샘플 볼륨)
- 식물을 수중으로 부드럽게 흔들어 재배 용기에서 제거하고 흡수성 물질에 배치하여 여분의 물을 흡수하십시오. 식물의 견고성과 트랩의 크기에 따라 8-10 개의 식물 개인을 선택하십시오.
- 트래핑 기관을 베어링 잎 노드를 계산하여 대략 동등한 부분으로 각 촬영을 분할합니다. 각 촬영 세그먼트는 젊은, 중년, 오래된 함정을 나타내는 혼합 샘플을 수집하는 역할을합니다.
- 연동 펌프에 샘플 수집을 위해 얇은 유리 모세관과 Eppendorf 바이알을 부착합니다. 트랩 개구부에 모세관 팁을 삽입합니다. 진공 펌프를 사용하여 각 트랩에서 900 ±100 μL까지 각 트랩에서 모든 유체를 빨아들이고 각 트랩 연령 범주에 대해 수집됩니다.
- 프로티스탄 방목 실험을 위해 풀링된 트랩 유체의 200 μL의 삼중 하위 샘플을 사용하십시오. 섹션 3에 자세히 설명된 대로 즉시 처리합니다. 아래에 자세히 설명된 바와 같이 유체에 서식하는 미생물 성분의 다른 모든 분석을 위해 샘플의 나머지 ±300 μL을 보존하십시오.
- 섹션 2로 즉시 진행합니다.
2. 수집된 샘플의 고정
- Exp I 및 II: 각 샘플에서 2% 최종 부피:부피 농도를 얻기 위해 적어도 1시간 동안 포름알데히드와 함께 세균 열거용 물/트랩 유체 하위 샘플을 수정합니다(섹션 4; 약 20 mL 및 0.3 mL).
참고: 포름알데히드를 연기 후드에서만 다루고 샘플을 조작하는 동안 항상 장갑을 착용하십시오.
3. 샘플 여과
- 시료 희석(Exp I): 저수지 물 샘플에 희석이 필요하지 않습니다. (Exp II): 에피소레센스 현미경 을 통해 계산하기 전에 필터 표면에 대상 미생물의 적절한 분포를 달성하기 위해 입자가없는 MQ 물로 트랩 유체 샘플 10x-100x를 희석하십시오.
- 여과 깔때기(직경 25 mm)를 사용하여, 검은 0.2 μm 기공 크기 필터에 세균 계수용 트랩 유체의 서브샘플의 저수지 물 또는 10-30 μL(Exp II)의 1-2 mL(Exp I)을 여과한다.
- DAPI (4', 6-diamidino-2'-페닐린돌 디하이드로 클로라이드, 0.2 % 최종 농도)로 필터를 4 분 동안 얼룩.
참고: 피부와 작업 표면 오염을 피하고 장갑을 착용하십시오. - 농축 된 미생물이있는 필터를 현미경 슬라이드에 침지 오일 한 방울 (형광 현미경용)에 놓습니다. 다른 오일 드롭을 필터 중앙에 놓고 커버슬립으로 덮어 오일이 고르게 분포되도록 합니다.
- 이 시점에서, 바람직하게는 시료를 즉시 처리하거나, 대안적으로, 추가 분석이 될 때까지 몇 주에서 몇 달 동안 냉동고(-20°C)에 저장한다.
4. 필터에 세균 번호의 열거
- 슬라이드를 에피오레시크 현미경 아래에 놓습니다(불소DAPI에 해당하는 필터 세트 포함). 10 x 10 카운트 그리드를 안구 중 하나에 놓습니다. 슬라이드를 임의의 위치로 이동합니다.
- 계수 그리드 의 영역에서 세균 세포 (청색 형광)를 정량화합니다 (배율 1000x 아래). 카운트에서 계수 그리드의 왼쪽 및 위쪽 가장자리를 가로지르는 셀을 포함하면서 오른쪽 및 아래쪽 가장자리에 있는 셀을 제외합니다.
- 다른 임의 위치로 이동하고 총 계산 500 셀에 달하는 적어도 10-15 카운트 그리드에서 열거를 반복합니다.
- 필터의 총 유효 여과 영역에 대한 한 그리드 면적의 비율에 대한 지식을 바탕으로 주어진 현미경 및 배율에 대한 변환 계수를 설정합니다. 그런 다음 계산된 그리드 수로 계산된 총 셀 수를 나누어 그리드당 평균 박테리아 수를 산출합니다.
- 후자의 매개 변수에 설정된 변환 계수에 곱하고 샘플의 mL당 결과 수를 정규화하여(여과된 샘플의 부피에 따라 다름) mL당 총 세균 풍부도를 얻습니다.
5. 프로티스탄의 풍요로움을 결정하다
- 글루타랄데히드(1% 최종 농도, 고정 후 며칠에서 몇 주까지 처리되는 엽록소 함유 입자가 있는 샘플에 더 적합) 또는 사용 시료(Exp I) 또는 트랩 유체(Exp II) 하위 샘플을 수정하거나 formol-thiosulfate 탈색 기법 아래에 명시된.
참고 : 두 보존 기술은 프로티스트 8의 식품 액구에서 섭취된 물질의 예를 들어 방지합니다. - formol-thiosulfate 탈색 기술에 관해서는, 물/트랩 유체 하위 샘플의 20 mL/200 μL에 Lugol 용액의 100 μL/1 μL을 각각 추가합니다.
- 붕산 완충 포르말린0.5 mL/50 μL을 첨가한 다음, 3% 티오설페이트 나트륨(익스펙 I/Exp II)의 20 μL/2 μL을 첨가한 후 즉시 사용하십시오.
참고 : 나트륨 티오설페이트는 루골의 황색을 탈색하여 상피 현미경8하에서세포를 관찰 할 수 있게합니다. - 1 μm 모공 크기의 검은 폴리 카보네이트 필터에 (대상 protists의 수에 따라) 샘플의 알려진 볼륨을 필터링합니다.
- 프로티스트 종의 수를 추정하는 것은 계수 그리드를 사용하여 배율 600x 하에서 적어도 200개의 세포를 계수함으로써 이다(위 참조).
- 여과 깔때기의 진공 하에서 낮은 진공으로 약 2 mL로 시료의 부피를 줄입니다. 이어서, 저압을 방출하고 DAPI 플루오로크롬(4',6-디아미디노-2-페닐린돌 디하이드로클로라이드13,0.2% 최종 농도)을 2분 동안 첨가한다.
6. 플랑크톤 샘플에서 섬모의 커뮤니티 구조 결정
참고 : 담수 서식지의 섬모 지역 사회는 매우다양14,15,16,18,그리고 그들의 현미경 결정은 도전이다. 섬모 군을 기능성 길드10,14,16,17로 분류하면 원골균으로 다른 섬모 그룹을 보다 자세하게 분석할 수 있습니다.
- 다음을 결합하여 섬모 커뮤니티 구조를 평가합니다.
- Epifluorescence 현미경 검사법에 있는 DAPI 염색한 견본 (다양한 크기 및 형태학의 거시및 마이크로 핵의 밝은 형광으로 섬모 세포를 현지화하기 위하여) 형광표지박테리아의 섭취와 결합 (FLB8;세부 사항, 아래 참조), 박테리아에 먹이 섬모의 능력을 추적.
- 선택한 경우17,19에서라이브 샘플 관찰 . 섬모를 다른 분류학적 범주로 그룹화하는 데 사용되는 위의 접근 방식과 기준에 대한 자세한 내용은 이전 간행물16,17을참조하십시오.
참고: 인용된 연구에 따르면 섬섬 중에서 잡식성 종인 스티토트리히아(제네라 할테리아 및 펠라고할테리아)와올리고트리히아(즉, 리모스트롬비디움 종)가 가장 중요한 원양 소비자라고 합니다. 담수 서식지의 대부분에서 박테리오 플랑크톤10,17,18.
7. 섬모 방목 율 추정
- 평균 트레이서 수의 변화에 기초하여 박테리아에 대한 섬모의 섭취 율을 계산[즉, 인큐베이션 시간(5-15분)]과 관련된 섬모 당 FLB8과 FLB의 트레이서 양이 추가되어 총 박테리아의 5%-15%를 차지합니다.
- 다른 protistan 종 중 의 관양 비율을 비교하기 위하여는, 실제 인큐베이션 시간 및 추적자 FLB의 비율에 근거한 계산을 가진 시간당 박테리아의 수로, 처리 비율을 정규화합니다 FLB 추가했습니다.
참고: FLB 방법 적용의 일반적인 방식은 천연 샘플에서 세포 또는 종별 및 벌크 박터리 비율을 모두 추정하는 도 1에도시되어 있다. - 민물 환경에 토착 세균 균주에서 FLB의 제조8
- 적절한 크기(평균 세포 부피 = MCV)와 박테리아의 형태를 선택하여 조사중인 수중 시스템에서 박테리오 플랑크톤 / 박테리아 세포의 전형적인 크기를 효과적으로 모방하십시오.
참고: Exp I의 경우, 림노바산과 폴리뉴클레오박터 속의 연구 위치에서 분리된 균주의 혼합물을 사용하였다(즉, 호수와 연못에서 전형적인, 매우 풍부한 박테리오플랑크톤)20. 균주의 형태와 크기에 대한 자세한 내용은 이전 간행물3,17,18,21을참조하십시오. - 초기 고정단계에서 15분 동안 원심분리(5,000 x g)에의해 배양으로부터 세균 세포를 수확하고 선택한 위치에서 박테리아의 전형적인 MCV에 대응하는 혼합물에서 세포의 MCV±SD를 산출하는 수치 비율로 혼합한다.
- 인산완식염수의 10 mL에서 펠릿을 일시 중단합니다(PBS; pH = 9).
- 인산염염수 완충액내의 세포 현탁액에 노랑-녹색 형광 염료 5-(4,6-디클로로트리아진-2-yl) 아미노플루오레세인(DTAF, 단백질에 결합)을 첨가하고, 2시간 동안 60°C 수조에서 배양한다.
- 인큐베이션 후, 세포를 아래로 원심분리하고, DTAF 용액을 분해하고, PBS로 3x를 세척 및 원심분리기를 한다.
- 최종 세척 후, PP i-식염수 완충제의 20 mL에서 세포를 다시 중단한다.
- FLB 현탁액 및 파이펫 1.5 mL aliquots를 2 mL 저온 바이알로 소용돌이한 다음, 사용 될 때까지 PPi-식염수 완충액에서 냉동 (-20 °C에서)을 유지합니다.
- 다음 단계에서사용하기 위해 0.2 μm 폴리카보네이트 필터를 통해 PP i-식염수 버퍼를 사전 필터한다.
- FLB 농도를 결정하려면 입자가없는 PPi-식염수 버퍼의 2 mL에 작은 aliquot (일반적으로 20-40 μL)를 전송하고, 몇 번의 2 s 버스트에 대해 30W에서 초음파 처리한 다음 epifluorescence를 통해 열거하기 위해 0.2 μm 폴리 카보네이트 블랙 필터에 필터링하십시오. DTAF(448 nm/520-540 nm)에 대한 광학 필터 설정에서 현미경 검사법(1,000X 배율)을 제공합니다.
- 적절한 크기(평균 세포 부피 = MCV)와 박테리아의 형태를 선택하여 조사중인 수중 시스템에서 박테리오 플랑크톤 / 박테리아 세포의 전형적인 크기를 효과적으로 모방하십시오.
- 섬모 박테리아의 추정을 위한 트레이서 기술
- 자연 플랑크토닉 서식지에서 방목 실험을 위해 300 mL 샘플을 잘 헹구고 1L 플라스크에 넣고 15 분 동안 현장에서 배양하십시오 (프로티스트가 취급 충격에서 회복 할 수 있도록).
- FLB 추적기를 추가하여 총 박테리아의 5%-15%를 구성하고 계절과 수온에 따라 양을 추가합니다.
참고 : 매우 광범위하고 계절에 따라 달라지는 스펙트럼이 있으며, 여러 계급의 크기에 걸친 종별 섭취량 (즉, 시간당 10-10-1 박테리아 섬모-1)에서 10,16, 17,18,22,23,24. - 높은 섭취 율 (일반적으로 여름 동안)와 섬모의 향상된 발생의 기간에, 또한 매우 낮은 FLB 추가와 병렬 배양을 실행, 만 구성 2%-4% 추적자 FLB에 의해 섬모 액포의 과도 하 중 하 중 을 피하기 위해 총 박테리아의 (참조) 그림 2)의예입니다.
- 5-15 분 동안 FLB와 섬모 / 플랑크톤 샘플을 인큐베이션.
- 프로티스트8의식품 액구에서 섭취 된 물질의 예를 들어 방지하기 위해 샘플 고정을위한 두 가지 가능성이 있습니다. 1% 글루타랄데히드(조류와 같은 엽록소 함유 입자가 있는 시료에 더 적합한 최종 농도)를 첨가하여 배양을 종료합니다. 또는, 루골 용액의 100 μL/10 μL을 20 mL/200 μL의 물/트랩 유체 하위 샘플에 사용하고, 바로 그 다음에 붕산성 완충포르민의 0.5 mL/10 μL을 추가한 다음 3% 나트륨 티오술페이트(Exp I/Exp II)의 200 μL/2 μL을 첨가합니다.
- 고정식을 첨가한 후, 섬모 세포의 철저한 보존을 보장하기 위해 4°C에서 암흑 속에서 적어도 1시간 동안 시료를 쉬게 한다.
- 4-30 mL/10-30 μL (Exp I / Exp II, 각각; 부피는 섬모 풍부에 따라 다름) 및 DAPI (최종 농도 0.2 % wt / vol; 자세한 내용은 위의 단계 3.2 참조)에서 자연 플랑크톤 하위 샘플을 가져 가라.
- 1 μm 블랙 필터를 통과하고 epifluoresccccins현미경을 통해 검사하여 섬모 (600x 배율)를 계산하고 이전 간행물2,17에 자세히 설명된 대로 섭취 된 FLB 추적기의 수를 열거하십시오 (대부분 1000x배율). . 보존 후 7일 이내에 시료를 검사하십시오.
- 총 원생 동물 / 종 별 방목을 추정하기 위해, 모든 섬모의 평균 섭취량을 곱하거나 현장에서 검출 된 섬모 종만 곱합니다.
- Římov 물 저수지에서 의 한 체 데이터에서 셀 당 통풍구 비율의 계산의 예는 다음과 같이 설명된다:
- 세균 농도가 3.55 x 106 박테리아 /mL이고 트레이서 FLB가 추가 된 추적자가 0.25 x 106 FLB / mL이라고 가정하면 총 세균 입자의 합계가 3.8 x 106 박테리아 / mL (천연 박테리아 + FLB = 먹이 입자의 100 %)을 산출합니다. 자연 샘플에서 식어 영양 프로티스트에게 사용할 수 있습니다.
참고: 이렇게 추가된 FLB 추적자는 총 세균 입자의 6.58%(0.25/0.038의 프로젝트)를 나타냅니다. 할테리아 스p. 당 FLB의 평균 수는 5 분 배양에서 6.2 FLB이다. - 시간당 섭취량을 정상화하려면 다음 계산을 사용하십시오: (6.2 x 12)/(6.58/100) = Halteria 세포/h당 1131개의 박테리아.
참고: 할테리아 sp. 가변 수온하에서 검출된 Halteria sp. 및 트레이서 FLB의 양(백분율)이 추가된 분포의 분포에 대한 자세한 예는 그림 3을참조하십시오.
- 세균 농도가 3.55 x 106 박테리아 /mL이고 트레이서 FLB가 추가 된 추적자가 0.25 x 106 FLB / mL이라고 가정하면 총 세균 입자의 합계가 3.8 x 106 박테리아 / mL (천연 박테리아 + FLB = 먹이 입자의 100 %)을 산출합니다. 자연 샘플에서 식어 영양 프로티스트에게 사용할 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
실시예 실험은 지모프 저수지(South Bohemia, CZ)에서 실행되었는데, 이는 현장 포식자와 먹이 풍부도에서 자연이 낮은 자연지대이다. 대표적인 데이터는 피코플랑크톤(&2 μm) 입자10,16,17,18의 풍부하고 효율적인 방목인 잡식성 섬모 종 할테리아 그란디넬라에대해 보고되고 있다. ,22. 그림 3은 호르히모프 저수지에서 할테리아 sp. 세포당 FLB 수의 상자 및 수염 플롯을 나타내며(그림3A)시간당 세균 섭취율로 재계산된 것(그림3B) 4월, 5월, 8월, 9월에 실시된 4건의 개별 실험에서 발견되었습니다. 섬모 섭취율의 변동성이 높았으며, 이는 주로 수온의 시간적 차이로 인한 것입니다.
Q10 매개 변수는 미생물 프로세스가 박테리아에 대한 섬모 섭취 율을 유지 10 °C12의온도 증가와 함께 약 2.5 배 빠르게 실행한다는 사실을 반영한다는 점에 유의해야한다. 이 생리학적 규칙을 염두에 두고, FLB 및 인큐베이션 시간의 상당히 다른 비율은 다른 계절에 사용되었다(자세한 내용은 그림 3A참조). 따라서, 예상 온도 효과에 대한 보상, 실험 설정은 섬모 세포 당 약 5-10 FLB 사이의 섭취 율의 최적화 된 평균 및 중앙값을 산출했다. 일반적으로 이러한 양의 섭취된 FLB는 쉽게 계산할 수 있으며(그림 2,왼쪽 사진 2개 참조) 추적자의 정확한 추정값(주로 섬모당 1-15FLB 사이)의 정확한 추정률을 생성합니다. 그러나 수정된 FLB 추적자로 인해 (%) 샘플 배양의 다른 시간 절대 값 (시간당 ciliate 방목 박테리아의 수로 발현) 크게 달랐다 (p < 0.01, Kruskal-Wallis 테스트; 던의 다중 비교 테스트 다음, p < 0.05; 그림의 예 참조 3B)실험 중. 이 데이터는 또한 할테리아 그란디넬라의플랑크토닉 인구에서 절대 세균 비율의 전형적인 자연 변동성을 보여 주며, 평균 값과 중간 값의 가까운 일치를 보여줍니다(그림 3).
복막 섬모와 같은 샘플에서 매우 효율적인 균사질 섬모가 있는 경우, 전체 박테리아의 5%-10%의 전형적인 트레이서 량으로 FLB에 의해 무겁게 "과표지"될 수 있습니다(그림 2의오른쪽 사진 참조). 이는 섭취한 FLB의 정확한 정량화를 강력하게 제한할 수 있습니다. 이러한 경우에, 그것은 단지 에 대 한 회계 FLB의 낮은 금액으로 추가 병렬 배양을 실행 하는 것이 좋습니다 만 1.5%-3% 총 박테리아의. 그러나, 일반적으로 트레이서 양뿐만 아니라 인큐베이션 시간은 세포당 FLB의 수를 최적화하기 위해 조작될 수있다(도 2).
예시 실험 II: 표시는 매우 작은 샘플 볼륨만 실험적으로 섬모 테트라 히메나 utriculariae의 박균 속도를 추정 할 수있는 큰 육식 동물과 먹이 풍부와 시스템의 데이터입니다 25. 그것은 육식 성 자궁 내 반사 반사 식물 26,27의함정에서 독점적으로 풍부하게 사는 적당한 세균 방목기입니다. 그림 4는 시간당 세균 섭취량의 비율로 재계산되는 다른 실험 설정하에서 T. utriculariae의 세포당 FLB수의 상자 및 수염 플롯을 나타낸다(그림4). C, D) 젊고 성숙하고 오래된 함정에서 발견됩니다. 흥미롭게도 트랩에서, 섬모 T. utriculariae의 엽록체 베어링 인구가 검출된 반면, T. utriculariae의 아포클로틱 집단은 트랩에서 분리되고 혼합 세균 현탁액에 성장하여 유지되었습니다. 밀 알갱이가 어둠 속에서 들어오세요(자세한 내용은 이전 발행물26의그림 1 참조).
엽록서 를 베어링 인구는 조명 트랩에 살고; 따라서, 엽록체는 섬모 호스트에 추가적인 유기 탄소 공급원 및 산소를 제공할 수 있다. 테스트 된 가설 중 하나는 박테리아가 섬모의 암흑 성장 고립 된 하위 집단에 사용할 수있는 유기 탄소의 유일한 미립자 소스를 나타내기 때문에 아포 클로릭 섬모 가산 이 박테리아를 상당히 빨리 방목했다.
실제로, 우트리큘리아 반사요법의 젊고 성숙하고 오래된 함정에 사는 섬모의 박균 비율에 유의한 차이가 없었지만(그림 4A,C), T. utriculariae의 아포클로틱 인구는 방목 박테리아는 크게 (p < 0.01, Kruskal-Wallis 테스트; 던의 다중 비교 테스트, p < 0.05), 젊고 성숙한, 오래된 트랩에 살고있는 엽록체 베어링 섬모보다 약 3 배 빠른(그림 4C,D). 다시 한 번, 트레이서 금액과 인큐베이션시간(그림 4A, B,상단)은 셀당 FLB 수(일반적으로 1-15사이)를 최적화하기 위해 수정되었으며 평균 값은 5 FLB/섬모입니다. 이 숫자는 섬모 식품 액포에서 구별할 수 있으며 정확한 트레이서 계수를 허용합니다. 그러나, 시간당 방목된 박테리아의 절대 수로 표현, 엽록체 베어링과 아포클로릭 인구는 각각 시간당 약 350 및 1,000 박테리아 섬모를 방목. 이 실험 설정은 현저하게 다른 환경 제약 조건25,26,같은 섬모 종의 두 개의 별개의 하위 집단의 대사 및 생리학적 특성에 대한 새로운 통찰력을 가져왔다. 27.
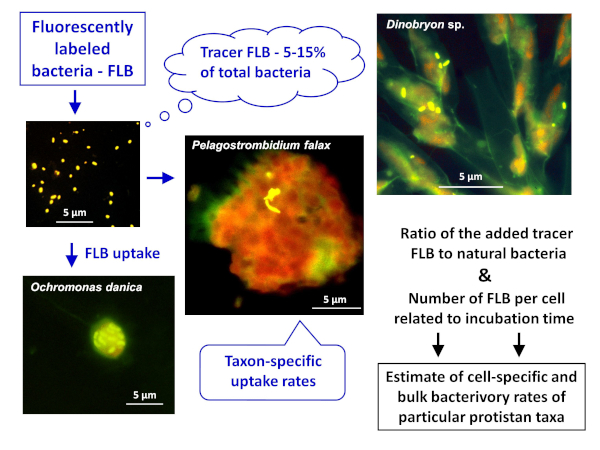
그림 1 : 형광 표지 된 박테리아 (FLB)를 사용하여 세포 및 종별 방목 속도를 샘플링의 총 천연 박테리아 수에 섭취 한 추적자 FLB의 비율에서 추정하는 워크플로우. 자세한 내용은 프로토콜의 섹션 7을 참조하십시오. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 2 : 부영양어연못의 플랑크톤에서 섬모세포의 예. 예는 섬모 세포에서 카운트 가능한 FLB를 가진 연못으로부터 도시된다 (일반적으로 세포 당 1-10 트레이서 FLB, 왼쪽 두 개의 마이크로 포토그라프) peritrichous 섬모 펠라고보르티켈라 나탄스 (오른쪽 마이크로 사진)에 비해. 짧은, 5 분 잠복기 동안에도, 그것은 트레이서 FLB에 의해 "과잉 라벨"되었다, 섭취 FLB의 양이 부정확하거나 거의불가능하게. 이 경우, 추적자 양을 총 박테리아의 1.5%-3%로 감소시킬 것이 좋습니다. 그러나, 일반적으로 트레이서 양 과 배양 시간 둘 다 세포 당 섭취된 FLB의 수를 최적화하기 위하여 조작될 수 있다. 자세한 내용은 프로토콜의 섹션 7을 참조하십시오. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 3 : 호르히모프 저수지(Exp I)(A)에서 할테리아 의 세포당 FLB 수의 박스 앤 수염 플롯을 시간당 세균 섭취량(B)으로 재계산합니다. 데이터는 4월부터 9월까지의 네 가지 예로 표시되는 다양한 계절 설정에서 감지되었습니다. 패널 A의 상단은 수온, 다른 FLB 추적자 추가 (%) 및 샘플 배양의 다른 시간에 대한 정보를 보여줍니다. 후자의 두 파라미터는 셀당 FLB 수를 최적화하기 위해 수정될 수 있으며, 평균(전체 라인) 및 중앙값(파선) 값은 섬모세포당 약 5-10 FLB(A)입니다. 막대는 모든 데이터의 25번째 및 75번째 백분위수(검사된 50-180셀)를 표시하고 수염은 1번째 및 99번째 백분위수입니다. (B)다른 작은 글자들은 연구 기간 동안 할테리아 sp. 세포 특이적 세균 비율에 유의한 차이를 나타낸다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 4 : 셀당 FLB 수의 상자 및 수염 플롯. 플롯은 엽록체 베어링 테트라히메나 우틸화 의 젊은, 성숙, 그리고 Utricularia 반사의 오래된 트랩 (Exp II)(A)의삼중 치료에서, 시간당 세균 섭취율로 재계산(C)로도시된다. 데이터는 트랩에서 분리된 T. upochloric 인구의 중복 된 아포클로틱집단의세균 섭취 율과 비교되었지만 어두운 밀 알갱이에서 자라는 혼합 세균 현탁액에서 유지되었습니다. 패널 A와 B의 상단에, 다른 FLB 추적자가 추가 (%) 및 샘플 배양의 다른 시간이 표시됩니다. 후자의 두 파라미터는 셀당 FLB 수를 최적화하기 위해 수정되었으며, 평균(전체 라인) 및 중앙값(파선) 값은 섬모 세포당 약 5-10FLB(A,B)입니다. 막대는 모든 데이터의 5번째 및 95번째 백분위수(검사된 50-100셀)를 표시하고 수염은 1번째 및 99번째 백분위수입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
수생 시스템에서 영양 상호 작용을 해독하는 것은 항상 도전28,특히 protists와 그들의 먹이, 박테리아를 포함하는 나노 플랑크톤 규모에서. 영양 섭취 경로 및 정량화에 관해서, 성공적으로 높은 영양 수준에서 사용 하는 방법의 응용 프로그램은 덜 가능, 생물 학적 상호 작용의 높은 복잡성으로 인해. 여기에는 예를 들어 안정적인 동위원소 라벨링 접근법이 포함됩니다. 이 프로토콜은 epifluoresccs 현미경 검사법과 형광표지 된 박테리아를 추적하여 탄소 흐름을 추적하고 반정화 / 추정하는 장점을 보여줍니다 (박테리아 먹이 : mixotrophic grazers29를포함한 다양한 프로티스탄 방목제) 미생물 식품 웹의 기초를 통해 경로. 이러한 장점 중 하나는 단일 셀 접근법의 높은 정확도이며, 다른 하나는 방목장 커뮤니티 구조와 다른 기능 적 길드, 종 (Exp I) 및 동일한 하위 집단을 구별하는 전례없는 해상도입니다. 종 (Exp II).
프로토콜의 중요한 단계
프로토콜에는 몇 가지 중요한 단계가 있으며, 이 단계는 방법론의 장점을 최대한 활용할 수 있도록 보장할 수 있습니다. 첫째, 실험개시 전에 연구된 환경에 대한 기본적인 이해는 항상 유익하다. 여기에는 잠재적 용 방목자의 다양성과 풍부의 현미경 스크리닝이 포함되며, 세균 먹이 크기 및 먹이 분포 둘 다 1) 물 기둥(예를 들어, 에필리온에서 저천액까지의 수직 프로파일) 및 2)의 경우 협곡 모양의 저수지, 댐 유입 이과정에서. 둘째, 수집 된 샘플로 신중하게 조작하면 대표적인 결과를 보장합니다. 온도는 프로티스트 방목속도를 포함하여 대부분의 미생물공정(12)에영향을 미치는 매우 중요한 인자이다(도3).
셋째, 샘플 내의 세균 세포 또는 방목장의 정량화에 기초하여 첨가된 트레이서의 양을 조작하면 과다 표지에 대한 문제가 제거될것이다(도 2). 섬모 종별 섭취 율에 매우 광범위한 스펙트럼이 있다는 점에 유의해야 합니다(자세한 내용은 7.2단계 참조). 따라서 프로토콜을 적절하게 적용하려면 시간 코스 섭취량이 있는 주요 섬모 종에 대한 사전 지식이 필수적입니다. 가능한 섬모 표지 (아무도 또는 너무 적은 FLB가 섬모 세포 당 채택, 통계적으로 건전하지 않은 데이터를 산출) 또는 과잉 라벨링 (FLB의 큰 숫자로 나타납니다) 피하기 위해 다른 추적자 금액으로 예비 실험을 실행하는 것이 좋습니다 "응축 FLB 구름"또는 추적자에 의해 포장 된 섬모 식품 액포에 무리를 형성, 따라서 심각하게 자신의 정확한 정량화를 제한; 그림 2)의오른쪽 위 예제를 참조하십시오. 또한 FLB를 함유한 배양 시간은 일반적으로 30분보다 짧으며, 섬모에 의한 피코플랑크톤의 평균 소화 시간은 약 1.5시간이며, 소화가 시작되기 때문에(섭취한 피코플랑크톤 세포는 전형적인 모양과 색상을 잃음) 후 45-60 분30. 마찬가지로, 정확한 결과를 얻으려면 현미경 보기 전에 필터에 대한 샘플의 최적의 희석 및 분포가 달성되어야 합니다.
수정 및 문제 해결
기술의 주요 단계, 가능한 수정 및 문제 해결 수정은 그림 1 및 그림 2에나와 있습니다. 또한, 고농도의 해로운 입자, 식물성 플랑크톤 세포 또는 플랑크톤의 식민지의 경우, 이러한 샘플 1) 개별 방목 세포가 될 수있는 단계를 달성하기 위해 그에 따라 희석되어야한다는 점에 유의해야합니다. 2) 식품 액액 함량의 정량화를 행한다.
제한
이 방법의 성공적인 적용을위한 주요 제한은 상피에서 명확한 샘플 보기를 방지 하는 금액에 부착 된 박테리아 또는 응집체와 다양 한 유기 detritus 또는 풍부한 무기/유기 입자의 존재에 있다 현미경및 추적자 양의 정확한 추정을 추가했습니다. 제시 된 추적 기 법은 주로 입자에 부착되지 않은 무료 (즉, 일시 중단 된) 박테리아와 함께 작동한다는 점에 유의해야합니다. 그러나, 우리 자신의 경험과 문학 참조에 따라 (이전 간행물2,4,8,10,16,18,21 참조 ,26),제시된 방법론은 대부분의 수중 환경에 적합하다. 영양 상태, 해악 함량 및 그라저 다양성 및 숫자가 다른 두 가지 자연적이고 대조적인 시스템의 예가제공됩니다(그림 3 및 그림 4).
기존 방법에 대한 접근 방식의 중요성
중요한 것은, 박테리버의 분류 /택시의 풍요로움과 종별 세균 비율에 대한 지식에서, 프로티스탄 분류 (또는 총 섬모 조립)의 대량 박통 비율을 계산할 수 있습니다. 이 접근법이 자연 플랑크톤 환경에 적용되면 이종영양 기종과 섬모 (박테리오 플랑크톤2,6,7의주요 방목자를 대표함) 프로티스탄 모두에 수반됩니다. 주어진 환경에서 세균 인구의 방목 유도 회전율 은16,17,18,22로추정 될 수있다. 이러한 데이터는 미생물 식품 웹에서 탄소 흐름 역학의 추정에 대한 근본적인 중요성을 보유하고 있습니다.
향후 응용 프로그램
일부 수정 사항을 사용하여 이 메서드를 성공적으로 사용할 수 있는 다른 특정 환경이 있습니다. 여기에는 활성 슬러지 시스템, 반추위 생태계, 수생 퇴적물 및 비대 어류 연못17이포함됩니다. 그러나 이러한 영양분 및 미생물이 풍부한 환경에서의 응용에는 일반적인 크기 분포 및 기타 특성을 모방할 수 있는 적절한 크기, 형태학 및 추적자 FLB 수에 관한 프로토콜을 최적화하기 위한 예비 테스트가 필요합니다. 환경에 내재된 먹이 박테리아의
현재, 이러한 접근법과 촉매 제자 증착 형광을 결합하는 데 관심이 증가하고 있는데, 이 경우 그라저 셀(예를 들어, 이종영양 기갑)의 정체가 특이적으로 검출되는 경우(CARD-FISH)가 검출되고 있다. FISH-프로브 및 섭취 율은 동일한 현미경슬라이드(31)에있는 기세포의 식품 액구에서의 FLB 함량을 기초로 한다. 이중 혼성화(32)라는 정교하고 새로운 접근법은 포식자 세포와 먹이 박테리아의 수준에서 FISH 프로브의 조합입니다 (이는 또한 계통 유전학 균주, 세균 계보 특정 FISH 프로브에 의해 특별히 표시됩니다). 이 접근법은 우아하지만 시간이 많이 걸리며 특정 기술과 경험31,32,다양한 FLB 섭취 접근법 수정의 적용이 실험실에서 일상적인 사용을 위해 더 쉽게 채택될 수 있다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 공개 할 것이 없다.
Acknowledgments
이 연구는 연구 보조금 13-00243S및 K. Š. 및 D. S.에게 각각 수여된 19-16554S의 밑에 체코 과학 재단에 의해 지원되었습니다. 이 기사는 또한 "댐 저수지의 수질 개선을위한 도구로 바이오 조작"(없음 CZ.02.1.01/0.0/0.0/16_025/0007417), 운영 프로그램 연구, 개발에 유럽 지역 개발 기금에 의해 지원되었다 교육.
Materials
| Name | Company | Catalog Number | Comments |
| 0.2-µm pore-size filters | SPI supplies, https://www.2spi.com/ | B0225-MB | Black, polycarbonate track etch membrane filters, diameter approprite for filtering apparatus used |
| 5-(4,6-dichlorotriazin-2-yl) aminofluorescein (DTAF) | Any brand | ||
| Automatic pipettes with adjustable volumes | Any brand, various sizes | ||
| Centrifuge | 22 000 x g | ||
| Cryovials | Any brand, 2 mL size | ||
| DAPI (4´,6-Diamidino-2´-phenylindole dihydrochloride) | Any brand | 1 mg ml-1 | |
| Epiflorescence microscope | Magnification from 400 x up to 1000 x | ||
| Filters appropriate for viewing in the DAPI and DTAF range | |||
| Counting grid in one of the oculars | |||
| Filtering apparatus | Usually with a diameter of 25 mm | ||
| Formaldehyde | A brand for microscopy | ||
| Glutaraldehyde | A brand for microscopy | ||
| Immersion oil for microscopy | Specific oil with low fluorescence | ||
| Lugol´s solution | Any brand or see comment | Make an alkaline Lugol' solution as follows: Solution 1 - dissolve 10 g of potassium iodide in 20 ml in MQ water, then add 5 g of iodine. Solution 2 - add 5 g of sodium acetate to 50 ml of MQ water. Add the solution 2 to the solution 1 and thoroughly mix | |
| Methanol stabilized formalin | Any brand available for microscopy purposes | ||
| Microscope slides and cover slips | Any brand produced for microscopy purposes | ||
| MQ water for diluting samples | Any brand |
||
| Phosphate-buffered saline (PBS; pH = 9) | Any brand | 0.05 M Na2HPO4-NaCl solution, adjusted to pH 9 | |
| PPi-saline buffer | Any brand | 0.02 M Na4P2O7-NaCl solution. Add 0.53 g Na4P2O7 to 100 ml of MQ water plus 0.85 g NaCl | |
| Sampling device | Appropriate for obtaining representative sample | e.g. Friedinger sampler for lake plankton | |
| Sodium thiosulfate solution | Any brand | 3% solution is used in the protocol | |
| Sonicator | Any brand | 30 W | |
| Vortex | Any brand allowing thorough mixing of the solutes and samples | ||
| Water bath | Any brand allowing temperature to be maintained at 60 °C |
References
- Azam, F., et al. The ecological role of water-column microbes in the sea. Marine Ecology Progress Series. 10, 257-263 (1983).
- Šimek, K., et al. A finely tuned symphony of factors modulates the microbial food web of a freshwater reservoir in spring. Limnology & Oceanography. 59, 1477-1492 (2014).
- Šimek, K., et al. Bacterial prey food characteristics modulate community growth response of freshwater bacterivorous flagellates. Limnology & Oceanography. 63, 484-502 (2018).
- Šimek, K., et al. Changes in bacterial community composition, dynamics and viral mortality rates associated with enhanced flagellate grazing in a meso-eutrophic reservoir. Applied & Environmental Microbiology. 67, 2723-2733 (2001).
- Jürgens, K., Matz, C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek. 81, 413-434 (2002).
- Pernthaler, J. Predation on prokaryotes in the water column and its ecological implications. Nature Reviews Microbiology. 3, 537-546 (2005).
- Berninger, U. B., Finlay, J., Kuuppo-Leinikki, P. Protozoan control of bacterial abundances in freshwaters. Limnology and Oceanography. 36, 139-147 (1991).
- Sherr, E. B., Sherr, B. F. Protistan grazing rates via uptake of fluorescently labeled prey. Handbook of Methods in Aquatic Microbial Ecology. Kemp, P. F., Sherr, B. F., Sherr, E. B., Cole, J. J. , Lewis Publishers. Boca Raton, Florida. 695-701 (1993).
- Vazquez-Dominguez, E., Peters, F., Gasol, J. M., Vaqué, D. Measuring the grazing losses of picoplankton: methodological improvements in the use of fluorescently tracers combined with flow cytometry. Aquatic Microbial Ecology. 20, 119-128 (1999).
- Šimek, K., et al. Ecological role and bacterial grazing of Halteria spp.: Small oligotrichs as dominant pelagic ciliate bacterivores. Aquatic Microbial Ecology. 22, 43-56 (2000).
- Montagnes, D. J. S., et al. Selective feeding behaviour of key free-living protists: avenues for continued study. Aquatic Microbial Ecology. 53, 83-98 (2008).
- Kirchman, D. L. Processes in Microbial Ecology. 2nd Edition. , Oxford University Press. Oxford, UK. (2018).
- Porter, K. G., Feig, Y. S. The use of DAPI for identifying and counting aquatic microflora. Limnology and Oceanography. 25, 943-948 (1980).
- Foissner, W., Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshwater Biology. 35, 375-482 (1996).
- Foissner, W., Berger, H., Schaumburg, J. Identification and ecology of limnetic plankton ciliates. Informationsberichte des Bayer Landesamtes für Wasserwirtschaft Heft. , 3-99 (1999).
- Šimek, K., et al. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: a study at the species and community level. Limnology & Oceanography. 40, 1077-1090 (1995).
- Skibbe, O. An improved quantitative protargol stain for ciliates and other planktonic protists. Archiv für. Hydrobiolgie. 130, 339-347 (1994).
- Macek, M., et al. Growth rates of dominant planktonic ciliates in two freshwater bodies of different trophic degree. Journal of Plankton Research. 18, 463-481 (1996).
- Šimek, K., et al. Microbial food webs in hypertrophic fishponds: omnivorous ciliate taxa are major protistan bacterivores. Limnology & Oceanography. , in press (2019).
- Jezbera, J., et al. Major freshwater bacterioplankton groups: Contrasting trends in distribution of Limnohabitans and Polynucleobacter lineages along a pH gradient of 72 habitats. FEMS Microbiology Ecology. 81, 467-479 (2012).
- Kasalický, V., et al. The diversity of the Limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS One. 8, 58209 (2013).
- Stabell, T. Ciliate bacterivory in epilimnetic waters. Aquatic Microbial Ecology. 10, 265-272 (1996).
- Zingel, P., et al. Ciliates are the dominant grazers on pico- and nanoplankton in a shallow, naturally highly eutrophic lake. Microbial Ecology. 53, 134-142 (2007).
- Bickel, S. L., Tang, K. W., Grossart, H. P. Ciliate epibionts associated with crustacean zooplankton in german lakes: distribution, motility, and bacterivory. Frontiers in Microbiology. 3 (243), (2012).
- Sirová, D., et al. Hunters or gardeners? Linking community structure and function of trap-associated microbes to the nutrient acquisition strategy of a carnivorous plant. Microbiome. 6, 225 (2018).
- Šimek, K., et al. Ecological traits of a zoochlorellae-bearing Tetrahymena sp. (Ciliophora) living in traps of the carnivorous aquatic plant Utricularia reflexa. Journal of Eukaryotic Microbiology. 64, 336-348 (2017).
- Pitsch, G., et al. The green Tetrahymena utriculariae n. sp. (Ciliophora, Oligohymenophorea) with its endosymbiotic algae (Micractinium sp.), living in the feeding traps of a carnivorous aquatic plant. Journal of Eukaryotic Microbiology. 64, 322-335 (2017).
- Nielsen, J. M., Clare, E. L., Hayden, B., Brett, M. T., Kratina, P. Diet tracing in ecology: Method comparison and selection. Methods in Ecology and Evaluation. 9, 278-291 (2018).
- Beisner, B. E., Grossart, H. P., Gasol, J. M. A guide to methods for estimating phago-mixotrophy in nanophytoplankton. Journal of Plankton Research. , 1-13 (2019).
- Dolan, J. D., Šimek, K. Processing of ingested matter in Strombidium sulcatum, a marine ciliate (Oligotrichida). Limnology and Oceanography. 42, 393-397 (1997).
- Massana, R., et al. Grazing rates and functional diversity of uncultured heterotrophic flagellates. The ISME Journal. 3, 588-596 (2009).
- Grujčić, V., et al. Cryptophyta as major freshwater bacterivores in experiments with manipulated bacterial prey. The ISME Journal. 12, 1668-1681 (2018).
