

Summary
Aqui apresentamos um protocolo para uma única célula, técnica baseada em microscopia de epifluorescência para quantificar as taxas de pastejo em eucariontes predatórios aquáticos com alta precisão e resolução taxonômica.
Abstract
Elucidar interações tróficas, como predação e seus efeitos, é uma tarefa frequente para muitos pesquisadores em ecologia. O estudo das comunidades microbianas tem muitas limitações, e a determinação de um predador, presa e taxas predatórias é muitas vezes difícil. Aqui apresentamos um método otimizado com base na adição de presas rotuladas fluorescentamente como traçador, o que permite quantificar de forma confiável as taxas de pastejo em eucariontes predatórios aquáticos e estimar a transferência de nutrientes para níveis tróficos mais elevados.
Introduction
Os procariontes heterotróficos são um componente biológico fundamental nos sistemas aquáticos e respondem por uma fração significativa da biomassa plâncton1,2,3. Fatores que controlam sua abundância, diversidade e atividade são cruciais para a compreensão de seu papel no ciclismo biogeoquímico (ou seja, o destino de carbono orgânico e outros nutrientes e fluxo de energia de procariontes a níveis tróficos mais elevados). O pastoreio de protozoan é um desses fatores importantes. Bacterivory de nanoflagelados e ciliados heterotróficas impõe um controle top-down forte sobre a abundância procarióticas, a função da Comunidade, a estrutura, a diversidade, e mesmo a morfologia celular e a taxa de crescimento de grupos bacterianos particulares4, 5,6. Em alguns sistemas, os protistas servem como a principal causa de mortalidade bacteriana6,7.
A aproximação padrão usada para avaliar o bacterivory do protozoário, que tem sido usada por algum tempo agora, envolve o uso de bactérias cDNAs etiquetadas (FLB) como análogos da rapina e microscopia do epifluorescência. As taxas de captação específicas de células podem ser determinadas quantificando o número de partículas de presas rotuladas em vacúolos de alimentos protistan ao longo de um curso de tempo selecionado8. Existem várias vantagens para esta abordagem. O Tracer é adicionado às amostras naturais com as assemblages naturais do predador e da rapina. Há uma manipulação mínima da amostra antes da incubação, alteração mínima da amostra pelo traçador FLB adicionado, e os tempos de incubação são curtos para garantir resultados sonoros obtidos em condições próximas a in situ. Alternativamente, em ambientes com baixo número de protistas bacterivorosos ou zooplâncton (por exemplo, sistemas marinhos offshore), as taxas de desaparecimento de FLB adicionadas a amostras em baixas quantidades (2%-3% traçador) podem ser detectadas via citometria de fluxo em longo prazo (12-24 h) experimentos de incubação. Em seguida, os números de FLB nos pontos inicial e final (integrando o impacto de todos os bacterivores) são quantificados por citometria de fluxo (para detalhes, ver publicação anterior9). No entanto, tal parâmetro representa apenas as taxas totais de bactérias agregadas que não podem ser diretamente atribuídas a qualquer grupo ou espécie de Grazer protistan e zooplâncton.
Em geral, a quantificação da espécie protistan-ou taxas de mortalidade bacteriana específicas do morphotype no ambiente aquático com precisão e com significado ecológico pode ser desafiador. Alguns protistas são pastosos seletivos, e o tamanho e a forma celular do traçador de FLB adicionado podem distorcer as taxas naturais de ingestão de presas10,11. Além disso, a atividade e o metabolismo do protistan são altamente temperatura-sensível12; Conseqüentemente, a quantidade de Tracer adicionado de FLB precisa de ser manipulada com cuidado para cada tipo individual da amostra (não somente baseado na abundância, no tamanho, e na morfologia naturais de bactérias e em tipos prevalecentes de bacterivores, mas igualmente na temperatura). A maioria dos estudos centra-se na atividade de pastoreio protistan granel; no entanto, o bactermarfim de espécies específicas protistan muitas vezes detém um valor de informação muito maior e pode ser preferível. Neste caso, é necessário o conhecimento taxonômico das espécies de protistas presentes em uma amostra e compreensão de seu comportamento. Portanto, quantidades consideráveis de tempo e trabalho são necessárias para obter resultados sonoros sobre as taxas específicas de bactérias de bacterivoria atribuíveis a um determinado grupo ou espécie protistan.
Apesar destas dificuldades, esta aproximação permanece a ferramenta a mais apropriada atualmente disponível para avaliar o bacterivoria do protistan em ajustes naturais. Aqui apresentamos um método abrangente e fácil de seguir para o uso de FLB como traçador em estudos de ecologia microbiana aquática. Todos os aspectos problemáticos mencionados da abordagem são contabilizados e um fluxo de trabalho melhorado é descrito, com dois experimentos de ambientes contrastantes, bem como contrastantes espécies ciliadas como exemplos.
O primeiro estudo de caso foi conduzido em um ambiente epilimnetic do reservatório de água mesotrófico de římov na República Checa, que mostra o Grazer e as abundâncias bacterianas comparáveis à maioria de corpos de água doce de superfície (cf.5,7). O segundo estudo de caso foi conduzido no ambiente altamente específico dentro das armadilhas da planta carnívoros aquática Utricularia reflexa, que hospeda números extremamente elevados de ambos os ciliados mixotróficas de pastoreio (utriculariae de Tetrahymena) e células bacterianas. Cálculos de taxas de pastejo específicas de células e estoques de pé bacteriano em ambos os tipos de amostra são mostrados. Uma escala de interpretações ecological dos resultados é discutida então, e os exemplos de estudos possíveis da continuação são sugeridos finalmente.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. coleta de amostra
-
Coleta de amostra de água do reservatório: o primeiro estudo de caso (exp I; menor predador natural in situ e sistema de abundância de presas)
- Colete amostras de água do local desejado em uma profundidade adequada. Mantenha as amostras em um refrigerador temperatura-controlado enchido na temperatura in situ (evitando o choque da temperatura; deve notar-se que as taxas de captação de protistas são temperatura-dependentes) durante o transporte para o laboratório.
Nota: nossa amostragem foi realizada no reservatório de Římov em forma de cânion meso-eutrófico (Boémia do Sul, volume 34,5 x 106 m3, profundidade máxima de 43 m, tempo médio de retenção de 100 dias, dimicótico). O local de amostragem foi localizado a uma profundidade de 30 m, perto da barragem. Uma amostra misturada em uma profundidade de 0,5 m foi coletada por um amostrador de 2-l Friedinger. - Continue até a seção 1,2 o mais rápido possível.
- Colete amostras de água do local desejado em uma profundidade adequada. Mantenha as amostras em um refrigerador temperatura-controlado enchido na temperatura in situ (evitando o choque da temperatura; deve notar-se que as taxas de captação de protistas são temperatura-dependentes) durante o transporte para o laboratório.
-
Coleta de fluido das armadilhas de plantas carnívoras de Utricularia reflexa: o segundo estudo de caso (exp II; um sistema com abundância de predadores e presas elevadas, volume de amostra extremamente pequeno)
- Agitar suavemente as plantas debaixo d' água, retire do recipiente de cultivo, e colocá-los em material absorvente para absorver o excesso de água. Dependendo da robustez das plantas e do tamanho das armadilhas, escolha 8-10 indivíduos da planta.
- Divida cada disparo em partes aproximadamente iguais contando os nós de folha que carregam órgãos de aprisionamento. Cada segmento de tiro servirá para coletar amostras mistas representando jovens, de meia-idade, e armadilhas antigas.
- Prenda um capilar de vidro fino e um frasco de Eppendorf para a coleção da amostra a uma bomba peristáltica. Proceder para inserir a ponta capilar na abertura da armadilha. Usando a bomba de vácuo, Sugar para fora todo o líquido de cada armadilha até 900 ± 100 μL do líquido da armadilha é coletado para cada categoria da idade da armadilha.
- Use subamostras de triplicado de 200 μl do líquido de armadilha agrupado para experimentos de pastoreio protistan. Processe estes imediatamente conforme detalhado na seção 3. Preservar o restante ± 300 μL da amostra para todas as outras análises de componentes microbianos que vivem no fluido, conforme detalhado abaixo (seção 2).
- Prossiga imediatamente para a seção 2.
2. fixação de amostras coletadas
- Exp I e II: correção de subamostras de fluido de água/armadilha para enumeração bacteriana (seção 4; aproximadamente 20 mL e 0,3 mL, respectivamente) com formaldeído por pelo menos 1 h para obter um volume final de 2%: concentração de volume em cada amostra.
Nota: Manuseie o formaldeído exclusivamente na capa das emanações e use luvas em todos os momentos durante a manipulação das amostras.
3. filtração da amostra
- Diluição da amostra (exp I): nenhuma diluição é necessária para as amostras de água do reservatório. (Exp II): diluir a amostra de fluido de armadilha 10x-100x com água MQ sem partículas para alcançar uma distribuição adequada dos micróbios alvo em superfícies filtrantes antes da contagem através de microscopia de epifluorescência.
- Filtrar 1-2 mL (exp I) da água do reservatório ou 10-30 μL (exp II) da subamostra do fluido de armadilha para contagem bacteriana em filtros pretos de tamanho de poros de 0,2 μm, usando um funil de filtração (25 mm de diâmetro).
- Manchar os filtros com DAPI (4 ', 6-diamidino-2 '-phenylindole dihydrochloride, 0,2% concentração final) por 4 min.
Nota: Evite a contaminação da pele e da superfície de trabalho e use luvas. - Coloque o filtro com micróbios concentrados sobre uma gota de óleo de imersão (para microscopia fluorescente) em uma lâmina de microscópio. Coloque outra gota de óleo sobre o centro do filtro e cubra com uma lamínula, certificando-se de que o óleo é distribuído uniformemente.
- Neste ponto, de preferência processar amostras imediatamente, ou alternativamente, armazená-las no congelador (-20 ° c) por várias semanas a meses até uma análise mais aprofundada.
4. enumeração de números bacterianos nos filtros
- Coloque o slide o microscópio de epifluorescência (com o conjunto de filtros correspondente ao fluorocromo DAPI). Coloque uma grade de contagem de 10 x 10 em um dos oculars. Mova o slide para uma posição aleatória.
- Quantificar as células bacterianas (fluorescência azul) na área da grelha de contagem (sob a ampliação 1000x). Nas contagens, inclua as células cruzando as bordas esquerda e superior da grade de contagem, excluindo as localizadas nas bordas direita e inferior.
- Mova para outra posição aleatória e repita a enumeração em pelo menos 10-15 grades de contagem, totalizando 500 células contabilizadas no total.
- Estabelecer um fator de conversão para um determinado microscópio e ampliação, com base no conhecimento da razão da área de uma grade para a área de filtração efetiva total do filtro. Em seguida, divida o número total de células contados pelo número de grades contados, produzindo um número médio de bactérias por grade.
- Multiplique o último parâmetro pelo fator de conversão estabelecido e normalizar o número resultante por ml de uma amostra (dependendo do volume da amostra filtrada) para obter a abundância bacteriana total por ml.
5. determinação da abundância protistan
- Fixar a amostra de água (exp I) ou as subamostras de fluido de armadilha (exp II) com glutaraldeído (1% de concentração final, mais adequada para amostras com a presença de partículas contendo clorofila a serem processadas dias a semanas após a fixação) ou utilizando o técnica de descoloração de formol-tiossulfato especificada abaixo.
Nota: ambas as técnicas de preservação impedem a egestion do material ingerido de vacúolos alimentares de protistas8. - Em relação à técnica de descoloração de formol-tiossulfato, adicionar 100 μL/1 μL de solução de Lugol em 20 mL/200 μL da subamostra de fluido de água/armadilha (exp I/exp II, respectivamente).
- Seguir imediatamente com a adição de 0,5 mL/50 μL de formalina tamponada em borato e, em seguida, 20 μL/2 μL de tiossulfato de sódio a 3% (exp I/exp II, respetivamente).
Nota: o tiossulfato de sódio descoloração a cor amarela de Lugol para possibilitar a observação de células o microscópio de epifluorescência8. - Filtre um volume conhecido de amostra (dependendo do número de protistas de destino) em filtros de policarbonato preto de tamanho de poros de 1 μm.
- Estimar o número de espécies de protistas é contando pelo menos 200 células ampliação 600x usando a grade de contagem (ver acima).
- Reduza o volume da amostra um vácuo no funil da filtração pelo baixo vácuo a aproximadamente 2 mL. Em seguida, solte a subpressão e adicione o DAPI fluorocromo (4 ', 6-diamidino-2-phenylindole dicloridrato de13, 0,2% concentração final) por 2 min.
6. determinação da estrutura comunitária dos ciliados em amostras de plâncton
Nota: comunidades ciliate em habitats de água doce são altamente diversificadas14,15,16, 18, e sua determinação microscópica é desafiador. A classificação dos grupos ciliados em guildas funcionais10,14,16,17permite uma análise mais detalhada dos diferentes grupos ciliados como bacterivores pelágicos.
- Avalie a estrutura da Comunidade ciliate combinando o seguinte:
- Amostras de DAPI-manchadas na microscopia do epifluorescência (para localizar as pilhas ciliate com fluorescência brilhante de macro-e micronúcleos de tamanhos e de morfologia diferentes) combinadas com a tomada de bactérias cDNAs marcadas (FLB8; para detalhes, Veja abaixo), controlando a capacidade dos ciliados de se alimentar de bactérias.
- Observação da amostra ao vivo em casos selecionados17,18. Para obter mais detalhes sobre as abordagens e os critérios acima utilizados para o agrupamento de ciliados em diferentes categorias taxonômicas, ver publicações anteriores16,17.
Nota: estudos citados indicaram que entre os ciliados, espécies onívoras de Stichotrichia (gêneros halteria e Pelagohalteria) e oligotrichia (ou seja, rimostrombidium spp.) são os consumidores pelágicos mais importantes da bacterioplâncton na grande maioria dos habitats de água doce10,17,18.
7. estimativa das taxas de pastejo ciliado
- Calcule as taxas de captação de ciliados em bactérias com base em mudanças no número médio de traçador [i.e., FLB8 por Ciliato relacionado ao tempo de incubação (5-15 min)] e quantidade traçada de FLB adicionado, contabilizando no máximo 5%-15% do total de bactérias.
- Para comparar as taxas de captação entre as diferentes espécies de protistan, normalizar as taxas de captação como o número de bactérias por Ciliato por hora, com cálculos baseados no tempo real de incubação e proporção do traçador FLB adicionado.
Nota: o esquema geral da aplicação do método FLB para estimar as taxas de bactérias específicas de células ou espécies e a granel em amostras naturais é representado na Figura 1. - Preparação de FLB de cepas bacterianas nativas de um ambiente de água doce8
- Selecione um tamanho adequado (volume celular médio = MCV) e morfologia das bactérias para que eles efetivamente imitam os tamanhos típicos de bacterioplâncton/células bacterianas no sistema aquático que está sendo investigado.
Nota: para exp I, utilizou-se uma mistura de cepas isoladas do local de estudo do gênero Limnohabitans e Polynucleobacter (isto é, bacterioplâncton típico, altamente abundante em lagos e lagoas)20. Para detalhes sobre morfologia e tamanhos das cepas, ver publicações anteriores3,17,18,21. - Colher células bacterianas da cultura por centrifugação (5.000 x g) por 15 min na fase estacionária precoce e misturá-las em uma proporção numérica que produz MCV ± DP das células na mistura correspondente ao MCV típico de bactérias no local escolhido.
- Suspender os pellets em 10 mL de solução salina tamponada com fosfato (PBS; pH = 9).
- Adicionar 2 mg do corante fluorescente amarelo-verde 5-(4, 6-dichlorotriazin-2-yl) aminofluoresceína (DTAF, liga-se às proteínas) à suspensão celular no tampão fosfato-salino, e incubar em um banho de água de 60 ° c por 2 h.
- Após a incubação, Centrifugue as células para baixo, decantar a solução DTAF, e lave e centrifugue 3x com PBS.
- Após a lavagem final, re-suspender as células em 20 mL do tampão PPi-Saline.
- Vórtice da suspensão FLB e pipeta 1,5 mL alíquotas em 2 mL crio-frascos, em seguida, manter congelado (em-20 ° c) em PPi-tampão salina até o uso.
- Pré-filtro PPi-tampão Saline através de um filtro de policarbonato de 0,2 μm para uso na próxima etapa.
- Para determinar a concentração de FLB, transfira uma alíquota pequena (geralmente 20-40 μL) a 2 ml do amortecedori-Saline partícula-livre do PP, proceda em 30 W para diversas explosões de 2 s, e filtre no filtro do preto do policarbonato do μm 0,2 para a enumeração através da epifluorescência microscopia (1, 000X ampliação) configurações de filtro óptico para DTAF (448 nm/520-540 nm).
- Selecione um tamanho adequado (volume celular médio = MCV) e morfologia das bactérias para que eles efetivamente imitam os tamanhos típicos de bacterioplâncton/células bacterianas no sistema aquático que está sendo investigado.
- Técnica de traçador para estimativa de bactermarfim ciliado
- Para as experiências de pastoreio em habitats planctônicos naturais, dispense 300 ml de amostras em frascos de 1 L bem enxaguados e incubar na temperatura in situ durante 15 min (para permitir que os protistas se recuperem do choque de manuseamento).
- Adicionar traçadores FLB para constituir 5%-15% do total de bactérias, com os montantes adicionados dependendo da estação e temperatura da água.
Nota: há um espectro muito largo, dependente da época em taxas de captação específicas de espécies ciliate abrangendo várias ordens de magnitude (ou seja, de 101-104 bactérias ciliate-1 por hora)10,16, 17,18,22,23,24. - Em períodos de maior ocorrência de ciliados com altas taxas de captação (geralmente durante o verão), também executam uma incubação paralela com adições de FLB muito baixas, constituindo apenas 2%-4% das bactérias totais para evitar o carregamento excessivo de vacúolos ciliados por traçador FLB (ver exemplos na Figura 2).
- Incubar amostras de ciliados/plâncton com FLB por 5-15 min.
- Há duas possibilidades para a fixação da amostra para impedir a egestion do material ingerido dos vacúolos do alimento dos protistas8. Encerre as incubações pela adição de 1% de glutaraldeído (concentração final que é mais adequada para amostras com partículas contendo clorofila, como algas). Alternativamente, use 100 μl/10 μL de solução de Lugol em 20 mL/200 μL de subamostra de fluido de água/armadilha, seguido imediatamente pela adição de 0,5 mL/10 μL de formalina tamponada em borato e, em seguida, 200 μL/2 μL de tiossulfato de sódio a 3% (exp I/exp II, respectivamente).
- Após a adição do fixador, deixe as amostras descansar por pelo menos 1 h no escuro a 4 ° c para garantir a preservação completa das células ciliadas.
- Tome subamostras de plâncton natural de 4-30 mL/10-30 μL (exp I/exp II, respectivamente; o volume depende da abundância de ciliate) e mancha com DAPI (concentração final 0,2% WT/vol; para detalhes, veja a etapa 3,2 acima).
- Passe através de filtros pretos de 1 μm e inspecione através da microscopia de epifluorescência para contar ciliados (ampliação 600x) e enumerar o número de traçadores de FLB ingeridos (na maior parte na ampliação 1000x) como detalhado em publicações precedentes2,17 . Inspecione amostras no prazo de 7 dias após a preservação.
- Para estimar o pastejo total/espécies específicas, multiplicar as taxas médias de captação de todos os ciliados, ou apenas as espécies ciliadas detectadas pela abundância in situ.
- O exemplo do cálculo das taxas de captação por célula de dados in situ do reservatório de água de Římov é descrito a seguir:
- Suponha que a concentração bacteriana é 3,55 x 106 bactérias/ml e traçador FLB adicionado é 0,25 x 106 FLB/ml, que produz uma soma de 3,8 x 106 bactérias/ml de partículas bacterianas totais (bactérias naturais + FLB = 100% de partículas de presa) disponíveis para protistas fagotróficos na amostra natural.
Nota: os traçadores de FLB adicionaram assim representam 6,58% (um projeto de 0.25/0.038) de partículas bacterianas totais. O número médio de FLB por Halteria SP. é 6,2 FLB em incubações de 5 min. - Para normalizar a captação por hora, use o seguinte cálculo: (6,2 x 12)/(6.58/100) = 1131 bactérias por célula Halteria /h.
Nota: para obter mais exemplos de distribuições de taxas de captação individuais de Halteria SP. detectadas temperaturas de água variáveis, quantidades (como percentuais) de traçador FLB adicionadas e diferentes épocas de incubação com FLB, ver Figura 3.
- Suponha que a concentração bacteriana é 3,55 x 106 bactérias/ml e traçador FLB adicionado é 0,25 x 106 FLB/ml, que produz uma soma de 3,8 x 106 bactérias/ml de partículas bacterianas totais (bactérias naturais + FLB = 100% de partículas de presa) disponíveis para protistas fagotróficos na amostra natural.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Experimento de exemplo fui executado no reservatório de água de římov (Boémia do Sul, CZ), que é um local natural com menor predador natural in situ e abundância de presas. Os dados representativos são relatados para a espécie onívora do ciliate halteria grandinella, que é um Grazer abundante e eficiente do picoplâncton (< 2 μm) partículas10,16,17,18 ,22. A Figura 3 mostra parcelas de caixa-e-Whisker de números de FLB por célula de halteria SP. a partir do reservatório de římov (Figura 3a), que foi recalculado para taxas de captação bacteriana por hora (Figura 3B) detectado em quatro experimentos individuais conduzidos em abril, maio, agosto e setembro. Houve alta variabilidade nas taxas de captação de ciliados, em grande parte causadas pelas diferenças temporais na temperatura da água.
Deve-se notar que o parâmetro Q10 reflete o fato de que os processos microbianos executam aproximadamente 2,5 x mais rápido com um aumento de temperatura de 10 ° c12, que também detém para as taxas de captação de ciliados em bactérias. Com esta regra fisiológica em mente, proporções consideravelmente diferentes de FLB e tempos de incubação foram utilizadas para diferentes épocas (para detalhes, ver Figura 3A). Assim, o efeito esperado da temperatura foi compensado, e o ajuste experimental rendeu valores médios e medianos otimizados de taxas da captação aproximadamente entre 5-10 FLB por a pilha ciliate. Geralmente, essas quantidades de FLB ingerida são facilmente contáveis (ver exemplos na Figura 2, duas fotografias esquerdas), gerando estimativas precisas do traçador (principalmente entre 1-15 FLB por Ciliato). Entretanto, devido ao traçador modificado de FLB adicionado (%) e diferentes épocas de incubação da amostra os valores absolutos (expressos em número de bactérias ciliadas por hora) diferiram significativamente (p < 0, 1, teste de Kruskal-Wallis; seguido pelo teste de múltipla comparação de Dunn, p < 0, 5; Ver exemplos na Figura 3B) entre os experimentos. Os dados também ilustram a variabilidade natural típica nas taxas absolutas de bacterina nas populações planctônicas de Halteria grandinella, com uma correspondência estreita de seus valores médios e médios (Figura 3).
Na presença de ciliados bacterivorous altamente eficientes nas amostras, tais como ciliados peritríquios, podem tornar-se pesadamente "excesso-etiquetado" por FLB em quantidades típicas do Tracer de 5%-10% das bactérias totais (veja a fotografia do lado direito em Figura 2). Isto pode limitar fortemente a quantificação exata de FLB ingerido. Nesses casos, sugere-se a execução de incubações paralelas adicionais com apenas baixas quantidades de FLB contabilizando apenas 1,5%-3% do total de bactérias. No entanto, geralmente tanto os valores do traçador quanto os tempos de incubação podem ser manipulados para otimizar o número de FLB por célula (Figura 2).
Exemplo de experimento II: exibido são os dados de um sistema com grandes abundâncias de predadores e presas, onde apenas um volume de amostra extremamente pequeno está disponível para estimar experimentalmente as taxas de bacterivoria do ciliado Tetrahymena utriculariae 25. é um Grazer bacteriano moderado que vive na abundância elevada exclusivamente nas armadilhas de plantas carnívoras de Utricularia reflexa 26,27. A Figura 4 mostra parcelas de caixa-e-Whisker do número de FLB por célula de T. utriculariae diferentes configurações experimentais (Figura 4A, B) que é recalculada em taxas de captação bacteriana por hora (Figura 4 C, D) detectado em armadilhas jovens, maduras e velhas. Curiosamente, em armadilhas, foram detectadas populações de cloroplastos do ciliado t. utriculariae , enquanto as populações apoclómicas de t. utriculariae foram isoladas de armadilhas e mantidas em suspensão bacteriana mista crescendo em grãos de trigo no escuro (para detalhes, ver Figura 1 em uma publicação anterior26).
As populações que carregam cloroplastos vivem em armadilhas iluminadas por luz; assim, os cloroplastos podem fornecer uma fonte de carbono orgânico adicional e oxigênio para o hospedeiro ciliado. Uma das hipóteses testadas foi que as populações de ciliados apoclóicos pastavam significativamente mais rapidamente, pois as bactérias representam a única fonte particulada de carbono orgânico disponível para subpopulações isoladas de origem escura do ciliado.
Na verdade, embora não houvesse diferenças significativas nas taxas de bacterina dos ciliados que viviam em armadilhas jovens, maduras e velhas de Utricularia reflexa (Figura 4a, C), as populações apoclóicas de T. utriculariae pastavam bactérias significativamente (p < 0, 1, teste de Kruskal-Wallis; seguido pelo teste de múltipla comparação de Dunn, p < 0, 5), aproximadamente 3x mais rápido do que os ciliados portadores de cloroplastos vivendo em armadilhas jovens, maduras e velhas (Figura 4C, D). Note-se que, novamente, tanto os valores do traçador quanto os tempos de incubação (Figura 4a, B, superior) foram modificados para otimizar o número de FLB por célula (geralmente entre 1-15), com média e mediana em torno de 5 FLB/ciliate. Estes números são distinguíveis em vacúolos ciliate do alimento e permitiram a contagem exata do Tracer. No entanto, expressa em números absolutos de bactérias pastadas por hora, as populações de cloroplastos e apocloricos pastaram aproximadamente 350 e 1.000 bactérias ciliadas por hora, respectivamente. Esta set-up experimental trouxe novos insights sobre os traços metabólicos e fisiológicos de duas subpopulações distintas da mesma espécie ciliada vivendo constrangimentos ambientais surpreendentemente diferentes25,26, a 27.
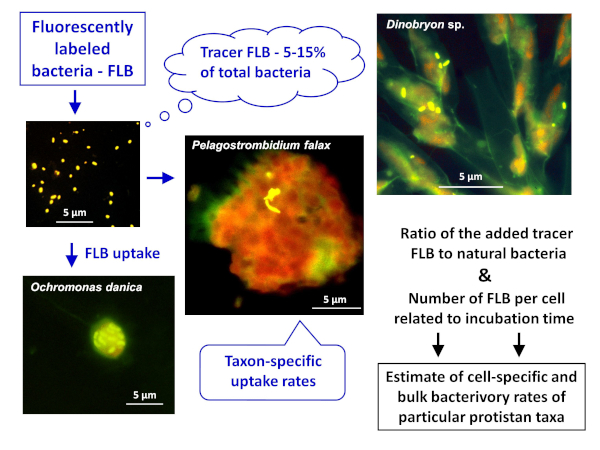
Figura 1 : Fluxo de trabalho de uso de bactérias rotuladas cDNAs (FLB) para estimar as taxas de pastejo de células e espécies específicas a partir da proporção de FLB traçador ingerido para o número total de bactérias naturais na amostra. Para obter mais detalhes, consulte a seção 7 do protocolo. Por favor clique aqui para ver uma versão maior desta figura.

Figura 2 : Exemplos de pilhas ciliate do plâncton de um fishpond eutrófico. Os exemplos são mostrados da lagoa com FLB contáveis em pilhas ciliate (geralmente 1-10 traçador FLB por a pilha, os dois microphotograps esquerdos) comparados a um ciliate peritríquios natans do pelagovorticella (a microfotografia do lado direito). Mesmo durante um curto, período de incubação de 5 minutos, tornou-se "over-rotulado" pelo traçador FLB, fazendo a quantificação do FLB ingerido impreciso ou quase impossível. Neste caso, sugere-se diminuir o valor do traçador para 1,5%-3% do total de bactérias. No entanto, geralmente os valores de traçador e os tempos de incubação podem ser manipulados para otimizar o número de FLB ingerido por célula. Para obter mais detalhes, consulte a seção 7 do protocolo. Por favor clique aqui para ver uma versão maior desta figura.

Figura 3 : Parcelas caixa-e-Whisker de números de FLB por célula de Halteria SP. do reservatório de Římov (exp I) (a), recalculado para taxas de captação bacteriana por hora (B). Os dados foram detectados em diferentes configurações sazonais, representados por quatro exemplos de abril a setembro. A parte superior do painel A mostra informações sobre a temperatura da água, diferentes traçadores FLB adicionados (%) e diferentes épocas de incubação da amostra. Deve-se notar que os dois últimos parâmetros podem ser modificados para otimizar o número de FLB por células, com valores médios (linha completa) e mediana (linha tracejada) aproximadamente entre 5-10 FLB por células ciliadas (A). As barras mostram os percentis 25 e 75 de todos os dados (50-180 células inspecionadas) e os bigodes estão para os percentis 1 e 99. (B) letras pequenas diferentes indicam diferenças significativas nas taxas de bacterocos específicos de células de halteria SP. durante o período estudado. Por favor clique aqui para ver uma versão maior desta figura.

Figura 4 : Parcelas de caixa-e-Whisker dos números de FLB por a pilha. As parcelas são mostradas dos Tetrahymena utriculariae do Chloroplast-rolamento dos tratamentos triplicados de armadilhas novas, maduras, e velhas de Utricularia reflexa (exp II) (a), recalculado às taxas de captação bacteriana por a hora (C). Os dados foram comparados com as taxas de captação bacteriana das populações apoclorímicas duplicadas de T. utriculariae (B, D) isoladas de armadilhas, mas mantidas em suspensão bacteriana mista crescendo em grãos de trigo no escuro. Na parte superior dos painéis A e B, diferentes traçadores FLB adicionados (%) e os tempos diferentes da incubação da amostra são mostrados. Deve-se notar que os dois últimos parâmetros foram modificados para otimizar o número de FLB por células, com valores médios (linha completa) e mediana (linha tracejada) aproximadamente entre 5-10 FLB por células ciliadas (A, B). As barras mostram os percentis 5 e 95 de todos os dados (50-100 células inspecionadas), e os bigodes estão para os percentis 1 e 99. Por favor clique aqui para ver uma versão maior desta figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Decifrar a interação trófica em sistemas aquáticos é sempre desafiador28, especialmente nas escalas de nano-plâncton envolvendo protistas e suas presas, bactérias. Quando se trata de vias de captação de nutrientes e quantificação, a aplicação de métodos utilizados com sucesso em níveis tróficos mais elevados é menos possível, devido à alta complexidade das interações bióticas. Estes incluem, por exemplo, abordagens de rotulagem de isótopos estáveis. Este protocolo mostra as vantagens de usar a microscopia do epifluorescência e as bactérias cDNAs etiquetadas como um Tracer para controlar e semiquantificar/estimar o fluxo de carbono (rapina bacteriana: vários pasters do protistan que incluem os pasters mixotróficas29) caminhos através da base de teias alimentares microbianas. Uma dessas vantagens é a alta precisão da abordagem de célula única, e a outra é a resolução sem precedentes sobre a estrutura da comunidade de Grazer e distinguir entre diferentes guildas funcionais, espécies (exp I) e até mesmo subpopulações do mesmo espécie (exp II).
Etapas críticas no protocolo
Existem várias etapas críticas no protocolo, que podem garantir que as vantagens da metodologia sejam utilizadas para seu pleno potencial. Primeiro, uma compreensão básica do ambiente estudado antes do início do experimento é sempre benéfica. Isso inclui a triagem microscópica da diversidade e abundância de potenciais herbívoros presentes, tamanhos de presas bacterianas e distribuição de presas tanto 1) na coluna de água (por exemplo, um perfil vertical do epílimnion para o hipolimnion) e 2) no caso de reservatórios em forma de cânion, no transect da barragem-Inflow. Em segundo lugar, a manipulação cuidadosa com amostras coletadas assegurará resultados representativos. A temperatura é um fator extremamente importante que afeta a maioria dos processosmicrobianos 12, incluindo as taxas de pastejo Protista (Figura 3).
Em terceiro lugar, manipular a quantidade de traçador adicionado com base na quantificação de células bacterianas ou tipo de Grazer na amostra irá garantir que os problemas com excesso de rotulagem (Figura 2) são eliminados. Deve-se notar que há um espectro muito amplo em taxas de captação específicas de espécies ciliadas (para detalhes, ver etapa 7,2); assim, para aplicar o protocolo apropriadamente, o conhecimento prévio de espécies ciliate principais com suas taxas de captação do tempo-curso é essencial. Recomenda-se fortemente para executar experimentos preliminares com diferentes quantidades de traçador para evitar possível ciliate Under-rotulagem (nenhuns ou demasiado poucos FLB são tomados acima por a pilha ciliate, produzindo dados estatisticamente doentio) ou over-rotulagem (aparece como um grande número de FLB formando "condensar as nuvens de FLB" ou rebanhos em vacúolos de alimentos ciliados embalados pelos traçadores, limitando assim severamente sua quantificação precisa; Veja o exemplo superior direito na Figura 2). Deve-se também notar que os tempos de incubação com FLB são geralmente mais curtos do que 30 min, uma vez que o tempo médio de digestão de picoplâncton por ciliados é de cerca de 1,5 h, e começa a digestão (células de picoplâncton ingeridos perde sua forma típica e cor) aproximadamente após 45-60 min30. Similarmente, a diluição e a distribuição óptimas das amostras no filtro antes da Visão microscópica precisam de ser conseguidas para resultados exatos.
Modificações e solução de problemas
As etapas principais, as possíveis modificações e a solução de problemas de modificações da técnica são ilustradas na Figura 1 e na Figura 2. Além disso, deve-se notar que, em casos de altas concentrações de partículas detritais, células fitoplanctônicas, ou suas colônias em plâncton, tais amostras 1) deve ser correspondentemente diluído para atingir um estágio em que as células de Grazer individuais podem ser distinto na superfície do filtro e 2) ser submetido à quantificação de conteúdo de vacuol de alimentos.
Limitações
A principal limitação para a aplicação bem-sucedida deste método reside na presença de vários detritos orgânicos ou partículas inorgânicas/orgânicas abundantes com bactérias anexadas ou agregados em quantidades que impedem a visualização clara da amostra a epifluorescência microscópio e estimativa precisa de um valor de traçador adicionado. Deve-se notar que a técnica de traçador apresentada funciona principalmente com bactérias livres (i.e., suspensas) que não estão ligadas a partículas. No entanto, com base em nossa própria experiência e referências bibliográficas (ver publicações anteriores2,4,8,10,16,18,21 ,26), a metodologia apresentada é adequada para a maioria dos ambientes aquáticos. São fornecidos exemplos de dois sistemas naturais contrastantes que diferem no estado trófico, no conteúdo de detritos e na diversidade e números de Grazer (Figura 3 e Figura 4).
Importância da abordagem em relação aos métodos existentes
Importante, do conhecimento da abundância de um táxon/táxons de bacteriófagos e suas taxas de bacterivoria espécies específicas, a taxa de bacterivoria a granel do táxon protistan (ou assemblage ciliate total) pode ser calculada. Se esta abordagem é aplicada a ambientes plâncton natural concomitantemente para ambos os flagelados e ciliados heterotróficos (representando os principais pastadores de bacterioplâncton2,6,7), oprotistan o tempo de giro induzido por pastejo de populações bacterianas em determinadoambiente pode ser estimado16,17,18,19. Tais dados têm importância fundamental para a estimativa da dinâmica do fluxo de carbono em teias de alimentos microbianos.
Aplicações futuras
Há outros ambientes específicos em que esse método, com algumas modificações, pode ser usado com êxito. Estes incluem sistemas de lodo ativados, ecossistemas de rúmen, sedimentos aquáticos e Fishponds hipertróficos17. No entanto, a aplicação nestes ambientes ricos em nutrientes e micróbios requer testes preliminares para otimizar o protocolo sobre o tamanho adequado, morfologia e números de FLB traçador que pode imitar a distribuição de tamanho típico e outras características de bactérias presas inerentes ao meio ambiente.
Atualmente, há interesse crescente em combinar essa abordagem com a hibridação in situ de fluorescência de deposição de repórter catalisada (cartão-peixe), na qual a identidade da célula de Grazer (por exemplo, flagelo heterotrófico) é detectada com um específico A sonda de peixe e a taxa de captação baseiam-se no teor de FLB em vacúolos alimentares da célula flagelada no mesmo slide microscópico31. Uma abordagem sofisticada e nova chamada hibridação dupla32 é uma combinação de sondas Fish nos níveis da célula predadora e das bactérias presas (que também são rotuladas especificamente por uma cepa filogenética, uma sonda de peixe específica de linhagem bacteriana). A abordagem é elegante, mas também demorada e requer habilidades específicas e experiência31,32, enquanto a aplicação de várias modificações de abordagem de captação de FLB pode ser mais facilmente adotada para uso rotineiro em laboratórios.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores não têm nada a revelar.
Acknowledgments
Este estudo foi apoiado pela Czech Science Foundation a subvenção de pesquisa 13-00243S e 19-16554S concedida a K. Š. e D. S., respectivamente. Este artigo também foi apoiado pelo projeto "Biomanipulação como uma ferramenta para melhorar a qualidade da água dos reservatórios de barragem" (no CZ. 02.1.01/0.0/0.0/16_025/0007417), financiado pelo Fundo Europeu de desenvolvimento regional, em pesquisa de programas operacionais, desenvolvimento e educação.
Materials
| Name | Company | Catalog Number | Comments |
| 0.2-µm pore-size filters | SPI supplies, https://www.2spi.com/ | B0225-MB | Black, polycarbonate track etch membrane filters, diameter approprite for filtering apparatus used |
| 5-(4,6-dichlorotriazin-2-yl) aminofluorescein (DTAF) | Any brand | ||
| Automatic pipettes with adjustable volumes | Any brand, various sizes | ||
| Centrifuge | 22 000 x g | ||
| Cryovials | Any brand, 2 mL size | ||
| DAPI (4´,6-Diamidino-2´-phenylindole dihydrochloride) | Any brand | 1 mg ml-1 | |
| Epiflorescence microscope | Magnification from 400 x up to 1000 x | ||
| Filters appropriate for viewing in the DAPI and DTAF range | |||
| Counting grid in one of the oculars | |||
| Filtering apparatus | Usually with a diameter of 25 mm | ||
| Formaldehyde | A brand for microscopy | ||
| Glutaraldehyde | A brand for microscopy | ||
| Immersion oil for microscopy | Specific oil with low fluorescence | ||
| Lugol´s solution | Any brand or see comment | Make an alkaline Lugol' solution as follows: Solution 1 - dissolve 10 g of potassium iodide in 20 ml in MQ water, then add 5 g of iodine. Solution 2 - add 5 g of sodium acetate to 50 ml of MQ water. Add the solution 2 to the solution 1 and thoroughly mix | |
| Methanol stabilized formalin | Any brand available for microscopy purposes | ||
| Microscope slides and cover slips | Any brand produced for microscopy purposes | ||
| MQ water for diluting samples | Any brand |
||
| Phosphate-buffered saline (PBS; pH = 9) | Any brand | 0.05 M Na2HPO4-NaCl solution, adjusted to pH 9 | |
| PPi-saline buffer | Any brand | 0.02 M Na4P2O7-NaCl solution. Add 0.53 g Na4P2O7 to 100 ml of MQ water plus 0.85 g NaCl | |
| Sampling device | Appropriate for obtaining representative sample | e.g. Friedinger sampler for lake plankton | |
| Sodium thiosulfate solution | Any brand | 3% solution is used in the protocol | |
| Sonicator | Any brand | 30 W | |
| Vortex | Any brand allowing thorough mixing of the solutes and samples | ||
| Water bath | Any brand allowing temperature to be maintained at 60 °C |
References
- Azam, F., et al. The ecological role of water-column microbes in the sea. Marine Ecology Progress Series. 10, 257-263 (1983).
- Šimek, K., et al. A finely tuned symphony of factors modulates the microbial food web of a freshwater reservoir in spring. Limnology & Oceanography. 59, 1477-1492 (2014).
- Šimek, K., et al. Bacterial prey food characteristics modulate community growth response of freshwater bacterivorous flagellates. Limnology & Oceanography. 63, 484-502 (2018).
- Šimek, K., et al. Changes in bacterial community composition, dynamics and viral mortality rates associated with enhanced flagellate grazing in a meso-eutrophic reservoir. Applied & Environmental Microbiology. 67, 2723-2733 (2001).
- Jürgens, K., Matz, C. Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Antonie Van Leeuwenhoek. 81, 413-434 (2002).
- Pernthaler, J. Predation on prokaryotes in the water column and its ecological implications. Nature Reviews Microbiology. 3, 537-546 (2005).
- Berninger, U. B., Finlay, J., Kuuppo-Leinikki, P. Protozoan control of bacterial abundances in freshwaters. Limnology and Oceanography. 36, 139-147 (1991).
- Sherr, E. B., Sherr, B. F. Protistan grazing rates via uptake of fluorescently labeled prey. Handbook of Methods in Aquatic Microbial Ecology. Kemp, P. F., Sherr, B. F., Sherr, E. B., Cole, J. J. , Lewis Publishers. Boca Raton, Florida. 695-701 (1993).
- Vazquez-Dominguez, E., Peters, F., Gasol, J. M., Vaqué, D. Measuring the grazing losses of picoplankton: methodological improvements in the use of fluorescently tracers combined with flow cytometry. Aquatic Microbial Ecology. 20, 119-128 (1999).
- Šimek, K., et al. Ecological role and bacterial grazing of Halteria spp.: Small oligotrichs as dominant pelagic ciliate bacterivores. Aquatic Microbial Ecology. 22, 43-56 (2000).
- Montagnes, D. J. S., et al. Selective feeding behaviour of key free-living protists: avenues for continued study. Aquatic Microbial Ecology. 53, 83-98 (2008).
- Kirchman, D. L. Processes in Microbial Ecology. 2nd Edition. , Oxford University Press. Oxford, UK. (2018).
- Porter, K. G., Feig, Y. S. The use of DAPI for identifying and counting aquatic microflora. Limnology and Oceanography. 25, 943-948 (1980).
- Foissner, W., Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshwater Biology. 35, 375-482 (1996).
- Foissner, W., Berger, H., Schaumburg, J. Identification and ecology of limnetic plankton ciliates. Informationsberichte des Bayer Landesamtes für Wasserwirtschaft Heft. , 3-99 (1999).
- Šimek, K., et al. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: a study at the species and community level. Limnology & Oceanography. 40, 1077-1090 (1995).
- Skibbe, O. An improved quantitative protargol stain for ciliates and other planktonic protists. Archiv für. Hydrobiolgie. 130, 339-347 (1994).
- Macek, M., et al. Growth rates of dominant planktonic ciliates in two freshwater bodies of different trophic degree. Journal of Plankton Research. 18, 463-481 (1996).
- Šimek, K., et al. Microbial food webs in hypertrophic fishponds: omnivorous ciliate taxa are major protistan bacterivores. Limnology & Oceanography. , in press (2019).
- Jezbera, J., et al. Major freshwater bacterioplankton groups: Contrasting trends in distribution of Limnohabitans and Polynucleobacter lineages along a pH gradient of 72 habitats. FEMS Microbiology Ecology. 81, 467-479 (2012).
- Kasalický, V., et al. The diversity of the Limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS One. 8, 58209 (2013).
- Stabell, T. Ciliate bacterivory in epilimnetic waters. Aquatic Microbial Ecology. 10, 265-272 (1996).
- Zingel, P., et al. Ciliates are the dominant grazers on pico- and nanoplankton in a shallow, naturally highly eutrophic lake. Microbial Ecology. 53, 134-142 (2007).
- Bickel, S. L., Tang, K. W., Grossart, H. P. Ciliate epibionts associated with crustacean zooplankton in german lakes: distribution, motility, and bacterivory. Frontiers in Microbiology. 3 (243), (2012).
- Sirová, D., et al. Hunters or gardeners? Linking community structure and function of trap-associated microbes to the nutrient acquisition strategy of a carnivorous plant. Microbiome. 6, 225 (2018).
- Šimek, K., et al. Ecological traits of a zoochlorellae-bearing Tetrahymena sp. (Ciliophora) living in traps of the carnivorous aquatic plant Utricularia reflexa. Journal of Eukaryotic Microbiology. 64, 336-348 (2017).
- Pitsch, G., et al. The green Tetrahymena utriculariae n. sp. (Ciliophora, Oligohymenophorea) with its endosymbiotic algae (Micractinium sp.), living in the feeding traps of a carnivorous aquatic plant. Journal of Eukaryotic Microbiology. 64, 322-335 (2017).
- Nielsen, J. M., Clare, E. L., Hayden, B., Brett, M. T., Kratina, P. Diet tracing in ecology: Method comparison and selection. Methods in Ecology and Evaluation. 9, 278-291 (2018).
- Beisner, B. E., Grossart, H. P., Gasol, J. M. A guide to methods for estimating phago-mixotrophy in nanophytoplankton. Journal of Plankton Research. , 1-13 (2019).
- Dolan, J. D., Šimek, K. Processing of ingested matter in Strombidium sulcatum, a marine ciliate (Oligotrichida). Limnology and Oceanography. 42, 393-397 (1997).
- Massana, R., et al. Grazing rates and functional diversity of uncultured heterotrophic flagellates. The ISME Journal. 3, 588-596 (2009).
- Grujčić, V., et al. Cryptophyta as major freshwater bacterivores in experiments with manipulated bacterial prey. The ISME Journal. 12, 1668-1681 (2018).
