

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Bu protokoller, kullanıcıların Denizatı hücre dışı akı analizini kullanarak 3D kanser hücre hattı kaynaklı sferoidlerde mitokondriyal enerji metabolizmasını araştırmalarına yardımcı olacaktır.
Abstract
Sferoidler olarak adlandırılan üç boyutlu (3D) hücresel agregalar, son yıllarda in vitro hücre kültürünün ön saflarında yer almaktadır. Hücreleri iki boyutlu, tek hücreli tek katmanlı (2D kültür) olarak kültürlemenin aksine, küresel hücre kültürü, hücre dışı matris proteinlerinin ekspresyonu, hücre sinyalizasyonu, gen ekspresyonu, protein üretimi, farklılaşma ve proliferasyon dahil olmak üzere in vivo olarak var olan fizyolojik hücresel mimariyi ve özellikleri teşvik eder, düzenler ve destekler. 3D kültürün önemi, onkoloji, diyabet, kök hücre biyolojisi ve doku mühendisliği dahil olmak üzere birçok araştırma alanında kabul edilmiştir. Son on yılda, sferoidler üretmek ve metabolik işlevlerini ve kaderlerini değerlendirmek için geliştirilmiş yöntemler geliştirilmiştir.
Hücre dışı akı (XF) analizörleri, bir XF24 adacık yakalama plakası veya bir XFe96 küresel mikroplaka kullanarak sferoidler gibi 3D mikro dokularda mitokondriyal fonksiyonu keşfetmek için kullanılmıştır. Bununla birlikte, farklı protokoller ve XF teknolojisini kullanarak sferoidlerde mitokondriyal enerji metabolizmasının araştırılmasının optimizasyonu ayrıntılı olarak tanımlanmamıştır. Bu yazıda, XFe96 XF analizörü ile küresel mikroplakalar kullanılarak tek bir 3D sferoidde mitokondriyal enerji metabolizmasının araştırılması için ayrıntılı protokoller sunulmaktadır. Farklı kanser hücre hatları kullanarak, XF teknolojisinin sadece farklı boyutlarda değil, aynı zamanda farklı hacimlerde, hücre sayılarında, DNA içeriğinde ve tipinde 3D sferoidlerde hücresel solunum arasında ayrım yapabildiği gösterilmiştir.
Oligomisin, BAM15, rotenon ve antimisin A'nın optimal mitokondriyal efektör bileşik konsantrasyonları, 3D sferoidlerde mitokondriyal enerji metabolizmasının spesifik parametrelerini araştırmak için kullanılır. Bu yazıda ayrıca sferoidlerden elde edilen verileri normalleştirme yöntemleri tartışılmakta ve XF teknolojisini kullanarak küresel metabolizmayı keşfederken göz önünde bulundurulması gereken birçok husus ele alınmaktadır. Bu protokol, gelişmiş in vitro sferoid modellerde araştırma yapılmasına yardımcı olacaktır.
Introduction
Biyolojik araştırmalarda in vitro modellerdeki ilerlemeler son 20 yılda hızla ilerlemiştir. Bu tür modeller artık çip üzerinde organ modalitelerini, organoidleri ve 3D mikrodoku sferoidlerini içeriyor ve bunların hepsi in vitro ve in vivo çalışmalar arasındaki çeviriyi geliştirmek için ortak bir odak noktası haline geldi. Gelişmiş in vitro modellerin, özellikle de sferoidlerin kullanımı, doku mühendisliği, kök hücre araştırması, kanser ve hastalık biyolojisi 1,2,3,4,5,6,7 ve genetik toksikoloji 8,9,10, nanomalzemeler toksikolojisi 11 dahil olmak üzere güvenlik testleri dahil olmak üzere çeşitli araştırma alanlarını kapsamaktadır. 12,13,14 ve ilaç güvenliği ve etkinliği testi8,15,16,17,18,19.
Normal hücre morfolojisi biyolojik fenotip ve aktivite için kritik öneme sahiptir. Hücreleri 3D mikrodoku sferoidlerine kültürlemek, hücrelerin in vivo olarak gözlemlenene daha çok benzeyen ancak klasik tek katmanlı hücre kültürü teknikleriyle yakalanması zor olan bir morfoloji, fenotipik fonksiyon ve mimari benimsemelerini sağlar. Hem in vivo hem de in vitro , hücresel fonksiyon, hücresel iletişim ve programlama ile sınırlı olmayan hücresel mikro çevreden doğrudan etkilenir (örneğin, hücre-hücre birleşme oluşumları, hücre nişleri oluşturma fırsatları); yakın çevrelerde hormonlara ve büyüme faktörlerine hücre maruziyeti (örneğin, enflamatuar yanıtın bir parçası olarak hücresel sitokin maruziyeti); fiziksel ve kimyasal matrislerin bileşimi (örneğin, hücrelerin sert doku kültürü plastiğinde mi yoksa elastik bir doku ortamında mı yetiştirildiği); ve en önemlisi, hücresel metabolizmanın beslenme ve oksijene erişimin yanı sıra laktik asit gibi metabolik atık ürünlerin işlenmesinden nasıl etkilendiği.
Metabolik akı analizi, tanımlanmış in vitro sistemlerdeki hücresel metabolizmayı incelemenin güçlü bir yoludur. Spesifik olarak, XF teknolojisi, bozulmamış hücrelerin ve dokuların hücresel biyoenerjetik özelliklerindeki canlı, gerçek zamanlı değişikliklerin analizine izin verir. Birçok hücre içi metabolik olayın saniyeler ila dakikalar arasında meydana geldiği göz önüne alındığında, gerçek zamanlı fonksiyonel yaklaşımlar, bozulmamış hücrelerde ve dokularda in vitro hücresel metabolik akıdaki gerçek zamanlı değişiklikleri anlamak için çok önemlidir.
Bu yazıda kanserden türetilmiş hücre hatları A549 (akciğer adenokarsinomu), HepG2/C3A (hepatosellüler karsinom), MCF-7 (meme adenokarsinomu) ve SK-OV-3 (over adenokarsinomu) hücre hatlarının in vitro 3D sferoid modeller kullanılarak zorla agregasyon yaklaşımları kullanılarak geliştirilmesi için protokoller sunulmaktadır (Şekil 1). Ayrıca, (i) Agilent XFe96 XF analizörü kullanılarak tek 3D sferoidlerin mitokondriyal enerji metabolizmasının nasıl araştırılacağını ayrıntılı olarak açıklar, (ii) tek 3D sferoidleri kullanarak XF analizlerini optimize etmenin yollarını vurgular ve (iii) bu yaklaşımı kullanarak 3D sferoid metabolizmasını araştırmanın önemli hususlarını ve sınırlamalarını tartışır. En önemlisi, bu makalede, hücresel sferoidlerde oksidatif fosforilasyonu ve dolayısıyla mitokondriyal fonksiyonu belirlemek için oksijen tüketim hızının (OCR) hesaplanmasına izin veren veri kümelerinin nasıl toplandığı açıklanmaktadır. Bu protokol için analiz edilmemiş olsa da, hücre dışı asitleşme hızı (ECAR), XF deneylerinde OCR verileriyle birlikte ölçülen başka bir parametredir. Bununla birlikte, ECAR genellikle XF veri kümelerinden kötü veya yanlış yorumlanır. Teknoloji üreticisinin temel yaklaşımlarını izleyerek ECAR'ı hesaplamanın sınırlamaları hakkında bir yorum sunuyoruz.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Şekil 1: Hücresel sferoidlerin oluşturulması, hücre dışı akı analizi ve aşağı akış tahlilleri için grafiksel iş akışı. Dört kanser hücresi hattı, doku kültürü şişelerinden ayrılan tek katmanlı (A) olarak seçici olarak kültürlendi ve sferoidler (B) oluşturmak için ultra düşük bağlanmalı 96 kuyucuklu mikroplakalara tohumlandı. A549 akciğer karsinomu, HepG2 / C3A karaciğer karsinomu, SK-OV-3 yumurtalık adenokarsinomu ve MCF-7 meme karsinomu hücreleri 1 × 10 3-8 × 103 hücre / kuyuda tohumlandı ve tek sferoidler oluşturmak ve sürekli gözlem ve planimetrik ölçümlerle sferoid tohumlama yoğunluğunu ve yetiştirme süresini optimize etmek için 7 güne kadar büyütüldü. Oluştuktan sonra, tek sferoidler serumsuz bir XF ortamına yıkandı ve poli-D-lizin (C) ile önceden kaplanmış sferoid tahlil mikroplakalarına dikkatlice tohumlandı. Sferoidler, XFe96 analizörü kullanılarak hücre dışı akı analizine tabi tutuldu: (1) bazal mitokondriyal solunum yanıtı için optimal küresel boyut; (2) mitokondriyal solunum inhibitörlerinin optimize edilmiş titrasyonları; (3) mikroplaka kuyuları içinde küresel yerleşimin optimizasyonu. (D) XF sonrası analizler, faz kontrast mikroskobu ve sferoid DNA nicelleştirmesi, veri normalizasyonu ve diğer aşağı akış in vitro tahlilleri için kullanılmıştır. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
1. Kanser hücre hatlarının 3D in vitro sferoidler olarak yetiştirilmesi
| Hücre hattı | Tarif | Kültür ortamı | Kaynak |
| A549 · | Akciğer karsinomu hücre hattı | RPMI 1640 Devir sayısı | Avrupa Doğrulanmış Hücre Kültürleri Koleksiyonu (ECACC) |
| Sodyum piruvat (1 mM) | |||
| Penisilin- Streptomisin - (100 U/mL – 100 mg/mL) | |||
| % 10 (v/v) FBS | |||
| HepG2/C3A | Hepatik karsinom hücre hattı, ana HepG2 hücre hattının klonal bir türevi | cesaret | Amerikan Doku Kültürü Koleksiyonu (ATCC) |
| Penisilin- Streptomisin - (100 U/mL – 100 mg/mL) | |||
| % 10 (v/v) FBS | |||
| MCF7 Serisi | Meme adenokarsinomu hücre hattı | RPMI 1640 Devir sayısı | Avrupa Doğrulanmış Hücre Kültürleri Koleksiyonu (ECACC) |
| Sodyum piruvat (1 mM) | |||
| Penisilin- Streptomisin - (100 U/mL – 100 mg/mL) | |||
| % 10 (v/v) FBS | |||
| SK-OV-3 | Yumurtalık adenokarsinomu hücre hattı | RPMI 1640 Devir sayısı | Avrupa Doğrulanmış Hücre Kültürleri Koleksiyonu (ECACC) |
| Sodyum piruvat (1 mM) | |||
| Penisilin- Streptomisin - (100 U/mL – 100 mg/mL) | |||
| % 10 (v/v) FBS | |||
| Parça | RPMI tahlil ortamı (50 mL son hacim) | ||
| Baz Orta | Çevik Denizatı XF RPMI, pH 7,4 | ||
| Glikoz (1 M steril stok) | 11 mM (0,55 mL stok çözümü) | ||
| L-glutamin (200 mM steril stok) | 2 mM (0,5 mL stok çözeltisi) | ||
| Sodyum piruvat (100 mM steril stok) | 1 mM (0,5 mL stok çözeltisi) |
Tablo 1: Kanser hücre hattı ortamı ve XF ortam bileşimleri.
- Standart aseptik doku kültürü tekniğini kullanarak tüm hücre hatlarını kültürleyin ve uygun bir tahlil kiti kullanarak mikoplazma içermediklerini onaylayın.
- Önerilen ortamı kullanarak T75 doku kültürü şişelerinde veya eşdeğerindeki hücre hatlarını kültürleyin (Tablo 1). Hücreyi% 65-80 oranında akıcılığa kadar kültürleyin ve bunları düzenli olarak en fazla 25 pasaja kadar geçirin.
- Hücre kültürü şişelerini Dulbecco'nun modifiye fosfat tamponlu salininde (DBPS) iki kez durulayın.
- Hücreleri, 37 ° C'de 5 dakika boyunca 3 mL hücre ayrışma reaktifi ile şişelerden ayırın ( Malzeme Tablosuna bakınız) ve mikroskopi ile ayrılmayı onaylayın.
- Tek hücreli bir süspansiyon sağlamak için ayrılmış hücre süspansiyonunu nazikçe aspire edin ve hücre ayrışma reaktifini 7 mL tam doku kültürü ortamı ile devre dışı bırakın.
- Hücreleri 5 dakika boyunca 300 × g'da santrifüjleme yoluyla toplayın, süpernatanı atın ve hücreleri tam ortamda yeniden askıya alın.
- Bir hemositometre veya otomatik bir hücre sayacı kullanarak hücreleri sayın ve tohumlama için gereken istenen hücre yoğunluğuna titre edin.
NOT: 96 delikli bir plakanın tamamını 4 × 103 hücre/kuyuda 100 μL/kuyucukta tohumlamak için, hücreler önerilen 12 mL'lik bir hacimde 4 × 104 hücre/mL'ye titre edilmelidir. - Hücre süspansiyonunu steril bir rezervuara boşaltın ve çok kanallı bir pipetör kullanarak hücre süspansiyonunun 100 μL'sini hücre itici bir mikro plakanın her bir kuyucuğuna dağıtın.
NOT: Bir mikroplakanın sadece iç 60 kuyucuğu tohumlanmalı ve geri kalanı DPBS ile doldurulmalıdır. Bu bir buharlaşma bariyeri oluşturacak, plaka boyunca küresel homojenliği sağlayacak ve plaka kenar etkilerini en aza indirecektir. - Hücreleri gevşek agregalara zorlamak için 300 × g'da küresel mikro plakaları 15 dakika boyunca santrifüj yapın.
- Küresel oluşumu sağlamak için plakaları en az 3 gün boyunca 37 °C'de,% 5 CO2'de inkübe edin.
- Sferoidlerin büyümesini izlemek için standartlaştırılmış laboratuvar uygulamalarını kullanarak faz kontrastlı mikroskopi gerçekleştirin. Hücre kültürü ortamını her 3 günde bir veya haftada iki kez, yarım hacimli bir ortam değişimi gerçekleştirerek doldurun.
2. Hücre Dışı Akı (XF) Teknolojisi kullanarak tek sferoidlerin mitokondriyal enerji metabolizmasının araştırılması
- Tahlil hazırlığı (bir gün önce)
- Bozulmamış küresel yapı, morfoloji ve numuneler arasında genel homojenlik sağlamak için 4x büyütmede faz kontrastına sahip ters çevrilmiş bir ışık mikroskobu kullanarak küresel canlılığı kontrol edin.
- Sensör kartuşunu nemlendirin.
- Kalibrantın ~ 20 mL'sini konik bir tüp haline getirir.
- Kalibranı içeren konik tüpü gece boyunca CO2 37 °C olmayan bir inkübatöre yerleştirin.
- İçeriği tahlil kitinden çıkarın.
- Sensör kartuşunu yardımcı plakadan çıkarın ve yardımcı plakanın yanındaki tezgahın üzerine baş aşağı yerleştirin.
- Çok kanallı P300 pipet kullanarak sensör kartuşu yardımcı plakasının her bir kuyucuğuna200μL steril ddH 2 O pipetin.
- Sensör kartuşunu yardımcı plakanın üzerine yerleştirin.
- Her bir kuyucuktaki su seviyesinin, sensör problarını suya batırmak için yeterince yüksek olup olmadığını kontrol edin.
- Takılan sensör kartuşunu CO2 olmayan 37 °C inkübatöre aktarın ve gece boyunca bırakın.
NOT: Bu adım, tahlil başlamadan 12-72 saat önce gerçekleştirilebilir.
- Ceket sferoid tahlil mikroplakası
- Aseptik teknikler kullanarak, sferoid mikroplakaya 30 μL / kuyucuk steril Poli-D-Lizin (0.1 mg / mL) çözeltisi ekleyin ve oda sıcaklığında 30 dakika boyunca inkübe edin.
- Çözeltiyi küresel mikroplakanın her bir oluğundan aspire edin, plakayı ters çevirin ve herhangi bir artık çözeltiyi çıkarmak için doku kağıdına sıkıca dokunun.
- Plakayı 200 μL/kuyucuk steril ddH2O ile iki kez yıkayın.
- Son yıkamadan sonra, mikro plakayı ters çevirin ve kalan suyu çıkarmak için kağıt mendile sıkıca dokunun.
- Kullanmadan veya ileride kullanmak üzere 4 °C'de saklamadan önce plakanın 30 dakika boyunca hava ile kurumasını bekleyin.
NOT: Sferoid tahlil mikroplakası, sferoidlerin mikroplakanın dibine sabitlenmesini sağlamak için moleküler bir yapıştırıcı ile kaplanmalıdır. Moleküler bir yapıştırıcı olmadan, sferoidler yerinden çıkabilir ve tahlil sonuçlarına müdahale edebilir. Diğer moleküler yapıştırıcılar, plakaları ön kaplamak için Poli-D-Lizin'e alternatif olarak da kullanılabilir. Önceden kaplanmış plakalar 4 ° C'de saklanabilir, ancak tahlil başlamadan önce oda sıcaklığına dengelenmelidir.
- XF Testi ortamını hazırlama
- Tablo 1'de ayrıntılı olarak açıklandığı gibi XF RPMI ortamını hazırlayın ve 0,22 μm şırınga filtreli steril filtre uygulayın
- Tahlil hazırlığı (tahlilden 1 saat önce)
- Takviyeli XF RPMI tahlil ortamını 37 ° C'ye kadar önceden ısıtın.
- Kaplanmış sferoid tahlil mikroplakasını CO2 37 °C olmayan bir inkübatörde veya kuru banyoda önceden ısıtın.
- Sensör kartuşunu hazırlayın.
- Kalibrant ve sensör kartuşunu içeren konik tüpü hava inkübatöründen çıkarın.
- Sensör kartuşunu yardımcı plakadan çıkarın ve çalışma yüzeyine baş aşağı yerleştirin.
- P300 çok kanallı pipet kullanarak, suyu kullanım plakasından aspire edin ve atın.
- Kalibrant çözeltisini steril bir reaktif haznesine dökün ve P300 çok kanallı pipet kullanarak ön ısıtılmış kalibrantın 200 μL/kuyucuğuna yardımcı plakaya ekleyin.
- Sensör kartuşunu alın ve tekrar yardımcı plakanın üzerine yerleştirin, böylece sensörlerin kalipranta iyice batırıldığından emin olun.
- Montajlı sensör kartuşunu, port enjeksiyon çözeltilerini yüklemeye hazır olana kadar CO2 37 °C olmayan inkübatöre geri aktarın.
- Sferoidleri tahlil ortamı ile yıkayın.
- Küresel kültür plakasını 37 °C,% 5 CO2 inkübatöründen çıkarın ve sferoid transfer adımlarından önce bütünlüklerini sağlamak için sferoidleri mikroskop altında gözlemleyin.
- Küresel plakanın tüm kuyularını, herhangi bir arka plan düzeltme kuyusu da dahil olmak üzere 180 μL / önceden ısıtılmış tahlil ortamı kuyusu ile yükleyin.
- 7 cm'lik bir Petri kabını 3 mL tahlil ortamı ile kısmen doldurun.
- Geniş delikli pipet uçlarıyla yüklü çok kanallı bir pipet kullanarak, pipetleyiciyi 10-50 μL'lik bir aspirasyon hacmine ayarlayarak sferoidleri 96 delikli kültür plakasından 7 cm Petri kaplarına aktarın.
- Önceden kaplanmış sferoid tahlil mikroplakasına tohum sferoidleri.
- Bir diseksiyon mikroskobu ve bir ışık kutusu aparatı kullanarak, sferoidleri Petri kabından aşağıda ayrıntılı olarak açıklandığı gibi sferoid tahlil mikroplakasına aktarın.
- Geniş delikli pipet ucuyla donatılmış tek kanallı pipetörün ses seviyesini 20 μL'ye ayarlayın ve tek bir sferoidi dikkatlice aspire edin. Ucu doğrudan sferoid tahlil mikroplakasının her bir kuyucuğunun ortasına yerleştirin ve yerçekiminin her bir kuyucuğun merkezine tek bir sferoid salınmasına izin verin, yani pipet ucundan herhangi bir ortamı dışarı atmayın ve kılcal hareketin sferoidi pipet ucundan çekmesine izin verin. Elüsyonu doğrulamak için, pipetleyicinin içeriği mikroskop altında 7 cm'lik Petri kabına geri pipetlenebilir.
NOT: Tek bir sferoidin yerçekimi elüsyonu, küresel boyuta / yoğunluğa bağlı olarak tipik olarak 15-30 s sürer. Bu süre zarfında pipetleyici çıkarılmamalıdır. Herhangi bir arka plan düzeltme kuyusu sferoidlerden arındırılmış olmalı ve sadece tahlil ortamı içermelidir. Mikroskop altında, her sferoidin konumunu onaylayın. Her küre ideal olarak her bir kuyucuğun ortasına yerleştirilmelidir. - Tüm sferoidler sferoid tahlil mikroplakasına aktarıldıktan sonra, plakayı tahlilden en az 1 saat önce 37 ° C'de CO2 olmayan bir inkübatöre aktarın.
- Geniş delikli pipet ucuyla donatılmış tek kanallı pipetörün ses seviyesini 20 μL'ye ayarlayın ve tek bir sferoidi dikkatlice aspire edin. Ucu doğrudan sferoid tahlil mikroplakasının her bir kuyucuğunun ortasına yerleştirin ve yerçekiminin her bir kuyucuğun merkezine tek bir sferoid salınmasına izin verin, yani pipet ucundan herhangi bir ortamı dışarı atmayın ve kılcal hareketin sferoidi pipet ucundan çekmesine izin verin. Elüsyonu doğrulamak için, pipetleyicinin içeriği mikroskop altında 7 cm'lik Petri kabına geri pipetlenebilir.
- Bir diseksiyon mikroskobu ve bir ışık kutusu aparatı kullanarak, sferoidleri Petri kabından aşağıda ayrıntılı olarak açıklandığı gibi sferoid tahlil mikroplakasına aktarın.
3. XF testleri için bileşiklerin hazırlanması ve sensör kartuşuna yüklenmesi
| Enjeksiyon Stratejisi | Bileşik (Bağlantı Noktası) | XFe96 mikro kuyucuk başlangıç hacmi (μL) | İstenilen nihai kuyu konsantrasyonu | Bağlantı Noktası Hacmi (μL) | Son XFe96 mikrowell hacimli enjeksiyon sonrası (μL) | Çalışma stoğu konsantrasyonu |
| 1 | Oligomisin (A) | 180 | 3 ug/mL | 20 | 200 | 30 μg/mL |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimisin A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimisin A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tablo 2: XFe96 Analizörü kullanılarak tek 3D sferoidlerin mitokondriyal enerji metabolizmasını araştırmak için mitokondriyal bileşik konsantrasyonları.
- Tamamen takviyeli, önceden ısıtılmış XF RPMI tahlil ortamı kullanarak Tablo 2'de belirtildiği gibi her bileşiğin çalışma stok konsantrasyonlarını hazırlayın.
- Kartuş plakasını (yardımcı plakaya bağlanmış) sütun yönünde, soldan sağa 1-12 olacak şekilde yönlendirin.
- Yükleme kılavuzu kullanıyorsanız, iyi yükleme prosedürüne göre kartuş plakasının üzerine yerleştirin, örneğin önce A bağlantı noktası yüklenecekse, kılavuzun sol üst köşesinde A'nın görünür olduğundan emin olun.
- Her bileşiğin çalışma çözeltisini uygun bir hazneye aktarın ve kalibre edilmiş bir P100 çok kanallı pipet kullanarak 20 μL'yi ilgili tüm portlara dağıtın. Her bileşik için kalan bağlantı noktalarına tekrarlayın.
NOT: Sensör kartuşu plakasında herhangi bir bağlantı noktası kullanılmazsa, bunlar boş bırakılabilir veya tahlil ortamıyla doldurulabilir. Yalnızca belirli bir port harfinin seçimi kullanılıyorsa, bu mektuba karşılık gelen diğer portların tahlil ortamı ile yüklendiğinden emin olun; aksi takdirde, kuyuya hava enjekte edilecek ve bu kuyulardaki sonuçlardan ödün verilecektir. - Bağlantı noktası yüklemesinden sonra, plaka yükleme kılavuzlarını (kullanılıyorsa) çıkarın ve analizörü sensör kartuşunu yüklemek üzere hazırlayın.
NOT: Bağlantı noktaları yüklendikten hemen sonra tahlil yapılmıyorsa, kapağı tekrar sensör kartuşuna yerleştirin ve makineye yüklenmeye hazır olana kadar plakayı 37 °C hava inkübatörüne geri koyun.
4. Tahlil tasarımı, enjeksiyon stratejileri ve veri toplama
- Tahlilin yürütülmesi
- Analizörü açın ve denetleyiciye (bilgisayar) bağlayın.
NOT: Bu, Wave Controller yazılımının widget panelindeki cihaz bağlantı durumu ile doğrulanabilir. - WAVE yazılımında şablonlar sayfasına gidin, denemenin tahlil şablonu dosyasını bulun ve çift tıklayarak açın.
NOT: Tahlil şablonu Şablonlar görünümünde görünmüyorsa, şablon dosyasını paylaşılan ağ sürücüsünden veya USB flash sürücüden şablon klasörüne alın. - Tahlili başlatmak için, Tahlil Çalıştır sekmesini tıklatın.
NOT: Grup tanımları plaka haritası içinde doğru şekilde ayrılmışsa, tahlil sayfanın sağ tarafındaki yeşil onay işaretiyle belirtildiği gibi çalışmaya hazır olacaktır. Bu aşamada, herhangi bir ek bilgi tahlil özeti sayfasına veya boş bırakılan sayfaya girilebilir; bir sonraki adıma geçin. 3D mikrodoku sferoidlerinde mitokondriyal modülatörlerin gecikmiş penetrasyonu nedeniyle (Şekil 2), Tablo 3'te açıklanan ölçüm protokolü bilgilerini kullanın.
- Analizörü açın ve denetleyiciye (bilgisayar) bağlayın.
| Ölçüm Süresi | Enjeksiyon Numarası ve Portu | Ölçüm Detayları | Süre (h:min:s) |
| Kalibrasyon | Uygulanamaz | XF analizörleri, ölçümlerin doğru olduğundan emin olmak için her zaman bu kalibrasyonu gerçekleştirir | 00:20:00 (bu ortalamadır ve makineler arasında değişebilir) |
| Equillibration | Uygulanamaz | Dengeleme Kalibrasyondan sonra gerçekleşir ve tavsiye edilir. | 00:10:00 |
| Bazal | Uygulanamaz | Döngüler = 5 | 00:30:00 |
| Karışım = 3:00 | |||
| Bekle = 0:00 | |||
| Ölçü = 3:00 | |||
| Oligomisin / BAM15 | Enjeksiyon 1 (Port A) | Döngüler = 10 | 01:00:00 |
| Karışım = 3:00 | |||
| Bekle = 0:00 | |||
| Ölçü = 3:00 | |||
| Rotenon + antimisin A | Enjeksiyon 2 (Port B) | Döngüler = 10 | 01:00:00 |
| Karışım = 3:00 | |||
| Bekle = 0:00 | |||
| Ölçü = 3:00 | |||
| Toplam Süre: | 03:00:00 | ||
Tablo 3: XFe96 Analizörü kullanılarak tek 3D sferoidlerin mitokondriyal enerji metabolizmasını araştırmak için protokol kurulumu.
- Konumu kaydet iletişim kutusunu açmak için çalıştırmayı başlat'ı tıklatın.
- Sonuç dosyasının kaydetme konumunu girin ve monte edilmiş sensör kartuşunu, analizörün yan tarafındaki kapıdan görünen termal tepsiye yerleştirin. Termal tepsinin otomatik olarak açılmasını ve ekranın Kalibrant Yükle Yardımcı Plakası mesajını görüntülemesini bekleyin. Ekrandaki talimatları izlemeden önce, i) sensör kartuşunun Yardımcı plakaya düzgün oturduğundan, ii) kapağın sensör kartuşundan çıkarıldığından ve iii) yardımcı plakadaki sensör kartuşunun doğru yönlendirildiğinden emin olun.
- Sensör kartuşu kalibrasyonunu başlatmak için ekrandaki komutları izleyin.
NOT: Kalibrasyonu tamamlamak için geçen süre yaklaşık 10-20 dakikadır (37 °C'de tahliller için). - Sensör kartuşu kalibrasyonundan sonra, 12 dakikalık dengeleme adımını başlatmak için Dalga Kontrol Cihazındaki ekrandaki talimatları izleyerek küresel mikro plakayı analizöre yükleyin.
NOT: Beyaz onay işaretli yeşil kutular, bu kuyu için 'iyi' bir kalibrasyonu gösterir. Herhangi bir kuyucuk 'iyi' bir kalibrasyon sağlayamazsa, kırmızı bir kutu ve beyaz haç ile belirtilecektir. Bu tür kuyular, değişiklik testi sekmesi kullanılarak tahlil tamamlandıktan sonra not edilmeli ve herhangi bir analizden çıkarılmalıdır. - Makine dengeleme adımını tamamladıktan sonra analizörün otomatik olarak taban çizgisi ölçümlerini almaya başlamasını bekleyin (cihaz protokolünde belirtildiği gibi).
- Denemeyi tamamlamak için WAVE denetleyicisindeki ekrandaki komutları izleyin.
NOT: Küresel mikro plaka analizörden çıkarıldıktan sonra, sensör kartuşunu atın ve gerekirse daha fazla analiz için küresel plakayı bir kenara koyun (örneğin, çift sarmallı (ds) DNA nicelemesi). Mikro plaka daha fazla analiz için gerekli değilse, sensör kartuşu ile birlikte atılabilir. - Tahlil iletişim kutusunun görünmesini bekleyin ve sonuçları görüntüleyin veya şablonlar görünümüne dönün.
5. Veri normalleştirme ve analiz stratejileri - tahlil sonrası normalizasyon ve aşağı akış tahlilleri (isteğe bağlı adımlar)
- Veri normalleştirme
- Küresel verileri normalleştirmek için, küresel boyut ve hacmi hesaplamak ve sferoid tahlillerde dsDNA'yı ölçmek için veri normalleştirme stratejileriyle ilgili protokoller dizisine bakın. Bunlar ek dosyalar olarak eklenmiştir; Ek dosya 1 ve Ek dosya 2'ye bakın.
- Veri analizi
- Verileri otomatik analiz üreteçlerinden birine aktarmak için, WAVE denetleyicisindeki veri dışa aktarma komutlarını izleyin ve tahlil türüyle eşleşen dışa aktarma oluşturucuyu seçin. Alternatif olarak, veri dosyasını dışa aktarın ve Seahorse analitiğine yükleyin.
NOT: Rapor üreteçlerinin ve Seahorse analitiğinin dezavantajı, veri analizinin XF testinin nasıl tasarlandığıyla sınırlı olması ve ölçüm döngüleri boyunca ortalamaların alınmasına izin vermemesidir. Veri kümelerinin cihaz yazılımından manuel olarak dışa aktarılması, bu konuda kullanıcı tercihine izin verir. 3D sferoidlerin mitokondriyal solunumunu değerlendirmek için enjeksiyon stratejisinin tipik bir 'MitoStress' testinden farklı olacağı göz önüne alındığında, 3D hücre kültürlerine özgü bu veri kümelerini analiz etmeye yardımcı olmak için bir dizi elektronik tablo şablonu geliştirilmiştir ve talep üzerine sağlanacaktır. Bu veri şablonu dosyaları, Şekil 2'de ayrıntılı ve açıklanan temel mitokondriyal solunum parametreleri hakkında veri sağlayacaktır. - Verileri analiz etmek için, verileri WAVE denetleyici yazılımından elektronik tablo raporu olarak dışa aktarın ve analiz için bağımsız bir elektronik tablo şablonu kullanın.
- Verileri otomatik analiz üreteçlerinden birine aktarmak için, WAVE denetleyicisindeki veri dışa aktarma komutlarını izleyin ve tahlil türüyle eşleşen dışa aktarma oluşturucuyu seçin. Alternatif olarak, veri dosyasını dışa aktarın ve Seahorse analitiğine yükleyin.

Şekil 2: Hücre dışı akı veri analizlerinden türetilen parametreler için şematik tanımlayıcılar. Kısaltma: OCR = oksijen tüketim oranı. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
İyi biçimlendirilmiş, kompakt sferoidler elde etmek için, her hücre hattı tohumlama yoğunluğu ve ekim süresi için ayrı ayrı optimize edilmiştir (Şekil 3). A549, HepG2 / C3A ve SK-OV-3 hücre hatları başlangıçta, kültürde 7 gün sonrasına kadar açıkça tanımlanmış çevrelere sahip yuvarlak sferoidlere ilerlemeyen gevşek agregalar oluşturdu. Tersine, MCF-7 hücreleri 3 gün içinde sferoidler oluşturabilir. Tüm sferoid modeller için kültür periyodundan sonra ilk hücre tohumlama yoğunluğu ile küresel hacim arasında açık bir korelasyon vardı. Küresel boyut ve morfoloji, tohumlama yoğunluğuna göre optimize edildi. Morfoloji ve döngüsellik, tüm modellerde artan küresel boyutla azalmaya başladı. Hücre hatları için tohumlama stratejileri, A549 ve SK-OV-3 hücreleri için 4 × 103 hücre / kuyuda optimize edilmiştir; HepG2 / C3A hücreleri daha önce başka bir yerde 1 × 10 3 hücre /kuyuya optimize edilmiş ve MCF-7 hücreleri tüm tahlillerde 4 × 103 hücre / kuyuda kullanılmıştır. Optimize edilmiş tohumlama stratejilerinde, küresel hacim 5.46 × 107 μm3 (SK-OV-3) ile 1.45 × 108 μm3 (A549) arasındaydı (Şekil 3B). Tüm sferoid tiplerin ilk tohumlama yoğunluğu ile sferoid hacim arasında doğrusal bir korelasyonu vardı, burada A549 ve HepG2 / C3A sırasıyla 0.957 ve 0.947 R2 değerlerine sahipti. MCF-7 ve SK-OV-3 sferoid hacimlerinin her ikisinin de ilk tohumlama yoğunluğu, R2 = 0.977 ile daha büyük bir korelasyona sahip olduğu bulunmuştur (Şekil 3A).
Küresel döngüsellik, FIJI analiz yazılımında uzun ve kısa küresel çaplar kullanılarak görüntü planimetrisi kullanılarak hesaplandı. Mükemmel küresel simetri daireselliğe sahipti = 1.0; 1.0'dan sapma dairesellik kaybına işaret ediyordu (Şekil 3C). MCF-7 sferoidlerinde dairesellik, daireselliğin tüm tohumlama yoğunluklarında 0.83 ila 0.9 arasında tutulduğu diğer modellerden daha fazlaydı. Buna karşılık, SK-OV-3 sferoidlerinin dış çevresi açıkça tanımlanmamıştır ve küresel hacim, kültürde 7 gün sonra bile önemli ölçüde daha küçüktü ve 4 × 103 / well tohumlama yoğunluğunda maksimum daireselliğe sahip sferoidler verdi. HepG2 / C3A hücrelerinin ayrıca, tüm sferoidlerin yüzey alanı boyunca eşit bir morfolojiye sahip sıkı, iyi biçimlendirilmiş sferoidler oluşturduğu ve daireselliğin 1 × 103 hücre / kuyuda tohumlanan hücreler için 0.79'da tutulduğu bulunmuştur. A549 hücrelerinin, küresel dairesellik ve morfolojinin tohumlama yoğunluğu ile arttırıldığı bir eğilimi takip ettiği görülmüştür; Bununla birlikte, döngüsellik bu deneylerde kullanılan yoğunlukta 0.63'ten büyük değildi.
Bazal mitokondriyal solunum, ultra düşük bağlanmalı küresel kültür mikroplakalarında 1 × 10 3, 2 × 10 3, 4 × 10 3 veya 8 × 10 3 hücre/kuyuda tohumlanan sferoidlerden ölçülen OCR olarak hesaplandı (Şekil 3D). Tüm sferoid tipleri için OCR, sferoid boyutu ile artmış ve 0.988'de MCF-7 sferoidlerinde R2 en yüksek ve 0.744'te SK-OV-3 sferoidlerinde en düşük olan sferoid hacimle doğrusal olarak korelasyon göstermiştir (Şekil 3E). Ölçülen OCR, tüm deney grupları arasında istatistiksel olarak farklıydı. A549 en düşük OCR'ye sahipti ve en büyük küresel boyutta sadece 18 pmol / dak / kuyu elde etti (Şekil 3D). Tersine, MCF-7 sferoidleri, kültürde sadece 3 gün sonra en küçük sferoid boyutunda benzer bir OCR verdi ve en büyük küresel boyut için maksimum 53 pmol / dak / kuyu başlangıç OCR'sine ulaştı (Şekil 3D). HepG2 / C3A, küresel boyut ve morfoloji ile oldukça tutarlı OCR verileri verdi. 1 × 10 3 hücre/kuyudan tohumlanan HepG2/C3A sferoidlerinde, başlangıç OCR ortalama 15 pmol/dak/kuyuya ulaşarak en büyük sferoidlerde maksimum 52 pmol/dk/kuyuya yükselmiştir (Şekil 3D). SK-OV-3 sferoidlerindeki OCR, sadece 4 × 10 3 hücreli / kuyu ve 8 × 10 3 hücreli / kuyudan yetiştirilen sferoidler arasında anlamlı idi ve 1 ila 103, 2 × 10 3 veya 4 × 10 3 hücre arasında yetiştirilen sferoidlerde çok az fark ×. Boyut farklılıklarına rağmen, OCR verileri tüm boyut noktalarında HepG2 / C3A ve MCF-7 sferoidleri arasında oldukça benzerdi. Sferoid boyuta (μm3) göre, MCF-7 sferoidleri tarafından bazal OCR, kuyu başına 1.000 hücreden 7 gün boyunca yetiştirilen HepG2 / C3A sferoidlerininkiyle karşılaştırılabilirdi.

Şekil 3: Küresel büyüme parametreleri başlangıç mitokondriyal solunumu belirler. (A) A549, HepG2 / C3A, MCF-7 ve SK-OV-3 sferoidleri tohumlama yoğunluğu için optimize edildi ve büyümeleri sırasıyla 1 × 10 3, 2 × 10 3, 4 × 10 3 ve 8 × fotomikrografların her birinde, sırasıyla sol üstten sağ alta doğru 10 3 hücre / kuyuda izlendi; ölçek çubukları = 500 μm. (B) Küresel boyut, toplanan fotomikrograflardan elde edilen planimetrik veriler kullanılarak hesaplandı ve Pearson'ın korelasyon istatistiği kullanılarak karşılaştırıldı; noktalı çizgiler %95 güven aralığının dağılımını temsil eder. (C) Küresel morfoloji, daireselliğin hesaplanmasıyla karşılaştırıldı. (D) OCR 5x ölçüldü, daha sonra Agilent Seahorse XFe96 analizörü kullanılarak mitokondriyal olmayan solunum hızını hesaba katmak için rotenon-antimisin A eklendi. OCR - OCRr/a olarak ölçülen OCRbazal, tohumlama yoğunluğu (D) ve küresel hacim (E) arasında karşılaştırıldı. Veriler, küresel tip ve hücre tohumlama yoğunluğu başına 5-8 kuyu replikasyonundan SEM'± ortalamalarıdır. Kısaltmalar: OCR = oksijen tüketim oranı; OCRbazal = Bazal mitokondriyal solunum; OCRr / a = rotenon-antimisin A ilavesinden sonra OCR. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
XF analizinde solunum modülatörlerine maruz kalma konsantrasyonu ve zaman seyri, tahlil optimizasyonunda kritik bir adımdır. Solunum modülatörü bileşikleri-oligomisin, BAM15, bir rotenon-antimisin A-veya DMSO araç kontrolü karışımı, sensör kartuşu enjeksiyon portlarından MCF-7 sferoidlerini içeren mikroplaka kuyucuklarına sırayla enjekte edildi (Şekil 4A). Tüm numune grupları için ortalama bazal OCR'nin 30-40 pmol/dak/kuyu olduğunu belirlemek için dört ölçüm döngüsü tamamlandı. Tahlil süresinin geri kalanı boyunca, enjeksiyon 1'den sonra 0.5 μM (0.5 μg / mL oligomisin) nihai kuyu konsantrasyonlarına ulaşmak için solunum modülatörleri her 5 ölçüm döngüsünde sırayla eklendi; Enjeksiyon 2'den sonra 2.0 μM (2 μg / mL oligomisin); Enjeksiyondan sonra 5 μM (5 μg / mL oligomisin) 3; ve son olarak, dördüncü sıralı eklemeden sonra maksimum 11 μM (11 μg / mL oligomisin) kuyu konsantrasyonu. MCF-7 sferoidleri deney boyunca araç kontrolüne cevap vermedi (Şekil 4A). Bazal OCR, her bir bileşiğin ilk enjeksiyonundan hemen sonra, en düşük konsantrasyonda 0.5 μM veya 0.5 μg / mL oligomisin (Şekil 4B) değişmiştir. MCF-7 sferoidlerindeki OCR, 0.5 μg / mL'lik ilk enjeksiyonu takiben 5 ölçüm döngüsünden sonra oligomisin ile 41 pmol / dak / kuyudan 23 pmol / dak / kuyuya düşürüldü (Şekil 4B).
0.5 μm BAM15'e yanıt olarak, OCR ikinci enjeksiyondan önce 33'ten 41 pmol / dak / kuyuya yükseltildi (Şekil 4C). Karşılaştırmalı olarak, rotenon artı antimisin A kombinasyonu, OCR'yi ikinci enjeksiyondan önce 37'den 13 pmol / dak / kuyuya düşürdü (Şekil 4D). Kinetik izler ayrıca OCR'de sabit bir doğrusal azalma (oligomisin ve rotenon-antimisin A) veya artış (BAM15) ortaya koydu. Tüm bileşik dozlama rejimleri için, 2 μM BAM15, 2 μM rotenon, 2 μM antimisin A ve 2 μg / mL oligomisin toplam kuyu konsantrasyonunda 10-12 tam ölçüm döngüsü (60-72 dakika) içinde kararlı durum OCR elde edildi (Şekil 4A). Oksijen tüketim hızı ~19 pmol/dak/kuyu (oligomisin), 52 pmol/dak/kuyu (BAM15) ve 10 pmol/dak/kuyu (rotenon-antimisin A) (Şekil 4A) sabit hal platosuna ulaşmıştır. Oligomisin, BAM15 veya rotenon artı antimisin A'nın bileşik konsantrasyonunun arttırılması, tahlilin geri kalanı boyunca sabit kalan OCR üzerinde belirgin bir etkiye sahip değildi. Bu veriler, 3D sferoidler kullanılırken hem bileşik konsantrasyonunun hem de solunum modülatörü bileşiklerine maruz kalma süresinin analiz optimizasyonu için dikkate alınması gerektiğini göstermektedir.

Şekil 4: Hücre dışı akı analizini optimize etmek için önemli bir adım olarak solunum modülatörü bileşiklerinin titrasyonlanması. (A) MCF-7 sferoidleri, XF RPMI içeren bir sferoid tahlil mikroplakasının kuyularına yerleştirilmeden ve XFe96 analizörü kullanılarak OCR × mitokondriyal modülatörler için problamadan önce 4 ± 103 hücre/kuyuda tohumlandı ve 3 gün boyunca kültürlendi. OCR 5x olarak ölçüldü, bundan sonra mitokondriyal ATP sentazı inhibe etmek, maksimum solunum kapasitesini belirlemek veya mitokondriyal olmayan solunum hızını belirlemek için sırasıyla araç kontrolü, oligomisin (B), BAM15 (C) veya rotenon-antimisin A (D) titrasyonları eklendi. Her mitokondriyal modülatörün konsantrasyonu, optimum bileşik konsantrasyonuna yanıt olarak maksimum kararlı durum OCR'sini belirlemek için 4 ayrı titrasyon enjeksiyon stratejisi (0,5 μM, 1,5 μM, 3 μM ve 6 μM; oligomisin için birimler μg/mL'dir) üzerinde artırılmıştır. OCR, her enjeksiyon arasında 5 ölçüm döngüsü için ölçüldü. Veriler, SEM'± 5-8 bireysel kuyudan çoğaltıldığı ortalamalardır. Kısaltma: OCR = oksijen tüketim oranı. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
XF teknolojisinin temel faydalarından biri, sağlam hücrelerde ve dokularda mitokondriyal fonksiyonu araştırabilme yeteneğidir. Hücrelerdeki ve dokulardaki mitokondriyal fonksiyonun belirli yönlerini incelemek için, mitokondriyal modülatörler, sensör kartuşu üzerindeki mevcut 4 enjeksiyon portu aracılığıyla numune mikroplakasının kuyucuklarına sırayla eklenir. XF tahlillerinde mitokondriyal parametreleri araştırmak için kullanılan tipik modülatör dizisi, oligomisin, bir protonofor (örneğin, FCCP veya BAM15) ve mitokondriyal ATP sentazı inhibe etmek, maksimum solunum kapasitesini belirlemek ve mitokondriyal olmayan solunum hızını düzeltmek için sırayla eklenen rotenon artı Antimisin A'nın bir kombinasyonudur. Bu tipik modülatör ilaveleri dizisi, tahlil teknolojisi üreticisi tarafından MitoStress testi olarak adlandırılır. Oligomisinin bazı hücre monokatmanları20'de uncoupler ile uyarılmış solunumu inhibe edebileceği göz önüne alındığında, bunu (tek) ve sonra (sıralı) oligomisin enjeksiyonundan önce (Şekil 5A-D) bağlanmamış uyarılmamış OCR'yi (OCRmax) ölçerek kanser kaynaklı 3D sferoidlerle inceledik. OCRmax, HEPG2 / C3A veya SK-OV-3'ten oluşan sferoidlere oligomisin eklenmesiyle anlamlı olarak sınırlı değildi (Şekil 5E ve Şekil 5G). Bununla birlikte, OCR max, oligomisin sonrası sıralı bir BAM15 enjeksiyonunu takiben A549 ve MCF-7 sferoidlerinde, tek bir BAM15 enjeksiyonundan elde edilen OCRmax'e kıyasla anlamlı derecede düşürülmüştür (Şekil 5F ve Şekil 5H). Bu nedenle, aksi bilinmedikçe, 3D sferoidlerin mitokondriyal enerji metabolizmasını araştırırken son bir rotenon ve antimisin A ilavesi ile oligomisin ve uncoupler ile tedavi etmek için ayrı kuyucukların kullanılması önerilir. Bu yaklaşım hala, bileşiklerin sırayla eklendiği tipik bir MitoStress testinde olduğu gibi tüm mitokondriyal parametrelerin hesaplanmasına izin vermektedir.

Şekil 5: Mitokondriyal solunum bileşiklerinin tek veya sıralı enjeksiyonu. MCF-7, HEPG2 / C3A, SK-OV-3 ve A549'un kanser hücresi kaynaklı sferoidleri, XF RPMI'daki bir XFe96 sferoid mikroplakasının kuyularına yerleştirildi ve Agilent Seahorse XFe96 analizörü kullanılarak OCR için araştırıldı. OCR, mitokondriyal ATP sentazı inhibe etmek ve maksimum solunum kapasitesini belirlemek için sırasıyla 2 μg / mL oligomisin (enjeksiyon Port A: yeşil iz) veya 5 μM BAM15 (enjeksiyon Port A: mavi iz veya enjeksiyon portu B: yeşil iz) ölçüldü. Kinetik OCR verileri % bazal (A-D) olarak ifade edilir. Maksimum solunum kapasitesi (OCR max), bazal OCR'nin bir faktörü olarak denklemle hesaplandı: OCRmax = OCRBAM15 / OCRbazal. OCRmax, (yeşil çubuklar) ve (mavi çubuklar) oligomisin içermeyen (yeşil çubuklar) enjeksiyon sonrası 8-10 ölçüm döngüleri boyunca OCR ortalamalarından elde edildi. Veriler, 3-8 bireysel kuyudan SEM'± küresel tahlil mikroplakası boyunca çoğaldığı ortalamalardır. Kısaltmalar: OCR = oksijen tüketim oranı. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Bu optimizasyon deneylerinde belirlenen optimal hücre tohumlama yoğunluklarını, bileşik konsantrasyonlarını, enjeksiyon stratejisini ve ölçüm döngüsü süresini kullanarak (Tablo 3), bazal mitokondriyal solunumu doğru bir şekilde araştırmak için ayrıntılı bir protokol geliştirdik: OCRbazal (Şekil 6A), ADP fosforilasyon solunum: OCRADP (Şekil 6B), sızıntı solunumu: OCRomi (Şekil 6C), bağlantı verimliliği (Şekil 6D ), maksimum solunum kapasitesi: OCRmax (Şekil 6E) ve yedek solunum kapasitesi: Kanser kaynaklı 3D sferoidleri kullanarak OCRyedeği (Şekil 6F).

Şekil 6: Kanser kaynaklı sferoidlerin mitokondriyal enerji metabolizmasını oluşturmak için OCR'yi XF teknolojisi ile araştırmak. MCF-7, HEPG2 / C3A, SK-OV-3 ve A549'un kanser hücresi kaynaklı sferoidleri, XF RPMI'daki bir sferoid tahlil mikroplakasının kuyularına yerleştirildi ve Agilent Seahorse XFe96 analizörü kullanılarak OCR için araştırıldı. OCR 5x ölçüldü, bundan sonra 2 μg / mL oligomisin veya 5 μM BAM15 ve mitokondriyal ATP sentazı inhibe etmek, maksimum solunum kapasitesini belirlemek ve mitokondriyal olmayan solunum hızını hesaplamak için RA eklendi. (A) Bazal mitokondriyal solunum (OCRbazal), port A enjeksiyonundan önceki 3 ölçüm döngüsünden OCR'nin ortalaması olarak hesaplanmıştır. (B) Oksidatif fosforilasyonun bağlanma etkinliği, OCRbazalinin bir yüzdesi olarak OCRADP (OCR bazal-OCRsızıntısı) eksprese edilerek yaklaştırılmıştır. (C) ADP fosforilasyon solunumu (OCRADP), BAM15 enjeksiyonundan önce 11-13 ölçüm döngüleri boyunca ortalama OCR'den hesaplanan oligomisin-duyarlı OCR olarak ölçülmüştür. (D) Sızıntıomi solunumu (OCRsızıntısı), oligomisin'e duyarsız OCR olarak ölçüldü ve ölçüm döngüleri 11-13 arasındaki ortalama OCR'den hesaplandı. (E) Maksimum solunum kapasitesi (OCRmax), BAM15 enjeksiyonunu takiben ölçülen ortalama maksimum OCR olarak ölçüldü. (F) Yedek solunum kapasitesi, OCRbazalinin bir yüzdesi olarak OCRyedeği (OCRmax - OCR bazal) eksprese edilerek hesaplandı. Rotenon-antimisin A enjeksiyonundan sonraki OCR (OCRr / a), mitokondriyal olmayan OCR'yi düzeltmek için tüm oranlardan çıkarıldı. Veriler, XFe96 küresel plaka boyunca 3-8 bireysel kuyudan SEM'± ortalamalarıdır. Kısaltmalar: OCR = oksijen tüketim oranı; RA = 2 μM rotenon-2 μM antimisin A. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
4 × 10 3 hücreden /3 günden fazla bir sürede yetiştirilen MCF-7 sferoidleri, sferoid tahlil mikroplakaları içinde optimum transfer, yerleştirme ve analizi belirlemek için bir model olarak kullanılmıştır. Üreticiden sferoid mikroplaka için sağlanan boyutlar kullanılarak, kuyu yüzeyi optimum küresel yerleştirme için üç bölge alanına bölünmüştür (Şekil 7A), burada bölge 1, kuyunun merkezinde en uygun bölge olarak vurgulanmıştır. Geniş delikli pipet uçları kullanılarak dikkatli pipetleme ile sferoidler sferoid plakalara aktarıldı ve yerçekimi elüsyonu ile kuyu yüzeylerine rastgele dağıtıldı (Şekil 7B). Sferoidlerin yerçekimi elüsyonu kullanılarak dikkatlice aktarıldığı yerlerde, çoğu sferoid, üreticiden önerilen transfer teknikleri kullanılarak, tipik olarak mikroplakanın 1-2 bölgelerinde bulunabilir. Sferoidlerin aspirasyonla pipet ucundan dışarı çıkmaya zorlandığı yerlerde, sferoidler genellikle bu bölgelerin ötesine yerleştirildi ve mikroskopi kullanılarak görülemedi.
Sferoid yerleştirme pozisyonlarını karşılaştırmak için, MCF-7 sferoidleri, belirlenmiş bölge 1-3 veya bölge dışında sferoid tahlil mikroplakalarına aktarıldı (Şekil 7A). Bu 4 kuyu, başlangıçta ve oligomisin, BAM15 veya rotenon-antimisin A ilavesinden sonra kinetik bir deney OCR ile izlendi (Şekil 7C). OCR, her enjeksiyondan önceki üç döngü okumasının ortalamasından hesaplandı (Şekil 7B). OCR, seçilen 4 kuyuda (Şekil 7C) 200 dakika boyunca kinetik olarak ölçüldü ve taban çizgisi düzeltildi (Şekil 7D). Sferoidlerin bölge 3'e veya bölge dışına yerleştirildiği yerlerde, taban çizgisi OCR'leri bölge 1 ve 2'ye yerleştirilen sferoidlerden anlamlı derecede düşüktü (Şekil 7C). Solunum bileşikleri oligomisin, BAM15 ve rotenon-antimisin A'nın etkileri, bölge 3 ve bölge dışı bölgelere kıyasla bölge 1 ve 2'ye yerleştirilen sferoidler arasında çarpıcı bir şekilde farklılık göstermiştir. OCR'de bir artış, bölge 3'e veya bölge dışına yerleştirilen sferoidlerde oligomisin ile görüldü (Şekil 7E). Ayrıca, bölge 3'e veya bölge dışına yerleştirilen sferoidler, rotenon-antimisin A enjeksiyonunu takiben OCR'nin taban çizgisinden daha yüksek olduğu BAM15'e aşırı yüksek bir yanıt vermiştir (Şekil 7E). Bazal OCR'de (Şekil 7C) bölge 2'ye karşı bölge 1'e yerleştirilen sferoidlerle neredeyse iki kat artışa rağmen, tüm solunum bileşiklerine yanıt olarak katlanma değişiklikleri çok benzerdi (Şekil 7E), bu da bölge 1 veya 2'ye yerleştirilen sferoidler arasındaki bazal OCR'deki farklılıkların kuyu içine yerleştirmenin sonucu olma ihtimalinin düşük olduğunu düşündürmektedir.

Şekil 7: Sferoidlerin sferoid tahlil mikroplakası içine yerleştirilmesi, XF teknolojisini kullanarak bazal OCR ve mitokondriyal modülatör etkilerini belirler. MCF-7 sferoidleri, 4 × 10 3 hücre/kuyuda tohumlandı ve XF RPMI içeren küresel mikroplakanın kuyularına yerleştirilmeden önce3 gün boyunca kültürlendi ve Agilent Seahorse XFe96 analizörü kullanılarak OCR ± mitokondriyal modülatörler için araştırıldı. (A) Tahlil süresinden sonra sferoid tahlil mikroplakalarındaki sferoid bölge konumlarının fotomikrografları; ölçek çubuğu = 500 μm ve zaman içinde karşılık gelen kuyulardan yakalanan OCR, pmol/min-1/kuyu-1 (B) veya % bazal (C) olarak ifade edilir. (D) Sferoid tahlil mikroplakası içindeki farklı bölgelere yerleştirilen MCF-7 sferoidlerinin mitokondriyal modülatör etkileri; bazaldan katlama değişimi olarak ifade edilen veriler. (E) E'de sunulan veriler için her mitokondriyal modülatörün tepkisini hesaplamak için hangi OCR veri ölçümlerinin (kırmızı daireler) kullanıldığını vurgulayan örnek kinetik iz. Gösterilen veriler bireysel kuyu yanıtlarından alınmıştır. Kısaltma: OCR = oksijen tüketim oranı. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Arka plan için seçim kriterleri çok önemlidir; Arka plan düzeltmesi için en dıştaki kuyucukların kullanılması, tüm mikro plaka kuyularını temsil etmez, bu da yanlış veri varsayımlarının çizilmesine ve küresel mikroplaka boyunca kenar etkileri nedeniyle hatalı veri sonuçlarına yol açabilir. Bu gözlemi değerlendirmek için, MCF-7 sferoidleri, bir araç kontrolü, oligomisin, BAM15 veya rotenon-antimisin A ilavesine yanıt olarak OCR değerlerini türetmek için tahlil düzeltme prosedürlerini karşılaştırmak için kullanıldı (Şekil 8). Tüm solunum bileşikleri, seçilen bileşikler için beklenen kinetik OCR profillerini verdi ve ortalama 20-30 pmol / dak / kuyu sabit bazal solunum hızı ortaya çıkardı (Şekil 8A). Bununla birlikte, tahlil verilerinin arka plan sıcaklık düzeltmesi için en dıştaki kuyucuklar (A1, A12, H1 ve H12) kullanılarak analiz edildiği yerlerde, solunum bileşiklerinin eklenmesinden sonra OCR için ortaya çıkan değerler özellikle düşüktü; OCR, rotenon-antimisin A için negatif değerler verdi. Bu gözlemlere yanıt olarak, arka plan sıcaklık düzeltme kuyuları olarak küresel mikroplaka boyunca rastgele dağılmış bir dizi boş kuyu kullanılarak alternatif analiz yapılmıştır (Şekil 8B). Alternatif arka plan düzeltmesinin uygulandığı yerlerde, OCR üzerindeki tüm göreceli bileşik etkiler her iki analiz setinde de aynıydı; ancak mutlak OCR değerleri yaklaşık 10 pmol/dk/kuyu artmıştır (Şekil 8). Bu veriler, küresel tahlil mikroplakaları üzerinde arka plan sıcaklık düzeltmesinin gücünü ve önemini vurgulamakta ve XF analizi için kullanıcı optimizasyonunun önemini vurgulamaktadır.

Şekil 8: Küresel tahlil mikroplakası boyunca sıcaklık gradyanlarının kontrolünü iyileştirmek için arka plan düzeltmesi için kuyuların rastgele seçimi. OCR verileri, arka plan düzeltmesi için önerilen kuyucuklar (A) ve arka plan düzeltmesi için rastgele atanmış kuyular (B) kullanılarak Şekil 2A'dan tahmin edilmiştir. Kısaltma: OCR = oksijen tüketim oranı. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Hücre monokatmanlarının aksine, sferoidler 3B bir alanda hücrelerin heterojen bir şekilde toplanmasını temsil eder ve bu nedenle, özellikle bu verileri normalleştirirken, analizle ilgili olarak kapsamlı bir şekilde düşünülmesi gerekir. Bu yazıda MCF-7 sferoidlerinden elde edilen XF verilerinin normalleştirilmesi için üç yaklaşım sunulmaktadır (Şekil 9). Normalleştirilmediğinde, OCR, Pearson korelasyon katsayısı, P = 0.0057 ile istatistiksel olarak karşılaştırıldığında, sferoid boyut (ilk hücre tohumlama yoğunluğu ile belirlenen) ile pozitif olarak (R 2 = 0.98) ilişkili (Şekil 9A). OCR ilk hücre tohumlama yoğunluğuna (R2 = 0.78) normalleştirildiğinde ve artık küresel boyutla anlamlı bir şekilde ilişkili olmadığında (P = 0.117, Şekil 9B) bu doğrusal ilişki azalır. Bu, küresel hacme normalleştirildiğinde de geçerlidir (R2 = 0.77; Pearson korelasyon katsayısı P = 0.120, Şekil 9C) ve nükleer dsDNA içeriği (R2 = 0.58; Pearson korelasyon katsayısı P= 0.233, Şekil 9D). Bu veriler, özellikle farklı boyutlardaysa, sferoidlerin mitokondriyal metabolizmasını araştırırken XF verilerinin normalleştirilmesinin önemini vurgulamaktadır.

Şekil 9: Hücresel sferoidlerden elde edilen hücre dışı akı verilerinin normalleştirilmesi. (A) Ham OCR verileri, 3 gün boyunca kültürlenmiş MCF-7'den elde edildi ve sferoid tohumlama yoğunluğu ile OCR arasında bir korelasyon katsayısı elde etmek için Pearson modeli kullanılarak çizildi; P değeri 0,05 olarak ayarlanmıştır. (B) Ham OCR verileri başlangıçtaki küresel tohumlama yoğunluğuna karşı normalleştirildi; (C) Mikroskopi planimetrisinden elde edilen MCF-7 sferoid hacim; ve (D) nükleer ds DNA içeriği Pearson'un korelasyon katsayısı kullanılarak karşılaştırılmıştır. Kısaltmalar: OCR = oksijen tüketim oranı; ds DNA = çift sarmallı DNA. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
| Hücre hattı | Tohumlama yoğunluğu (iyi) | Küresel büyüme (gün) | Son sferoid hacim (μM3) | Bazal OCR (pmolO2/dak/kuyu) | Bazal OCR met duyarlılığı (EVET/NO) |
| cesaret | 1000 | 5 | 9,52E+06 | 28 ± 3,5 | Evet |
| cesaret | 2000 | 5 | 2,38E+07 | 26 ± 1,4 | Evet |
| cesaret | 4000 | 5 | 4,92E+07 | 36 ± 3,1 | Evet |
| cesaret | 8000 | 5 | 1,11E+08 | 50 ± 7,9 | Evet |
| HepG2 | 1000 | 5 | 1,11E+07 | 15 ± 0.7 | Hayır |
| HepG2 | 2000 | 5 | 2,88E+07 | 23 ± 1,8 | Evet |
| HepG2 | 4000 | 5 | 5,46E+07 | 31 ± 1,7 | Evet |
| HepG2 | 8000 | 5 | 1,21E+08 | 52 ± 2,8 | Evet |
| A549 · | 1000 | 5 | 2,11E+07 | 30 ± 2,5 | Evet |
| A549 · | 2000 | 5 | 3,57E+07 | 41 ± 1,6 | Evet |
| A549 · | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Evet |
| A549 · | 8000 | 5 | 1,44E+08 | 65 ± 8,4 | Evet |
| MCF-7 Serisi | 1000 | 3 | 1,60E+07 | 29 ± 0,8 | Evet |
| MCF-7 Serisi | 2000 | 3 | 2,52E+07 | 37 ± 1,7 | Evet |
| MCF-7 Serisi | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Evet |
| MCF-7 Serisi | 8000 | 3 | 1,06E+08 | 66 ± 2,9 | Evet |
Tablo 4: Tek 3D sferoidlerde bazal OCR ölçümlerini belirlemek için optimize edilmiş parametreler. Kısaltma: OCR = oksijen tüketim oranı.
Ek Dosya 1: Küresel boyut ve hacmin analizi. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Dosya 2: Sferoid mikroplakadaki sferoidlerden çift sarmallı DNA'nın miktarı. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Dosya 3: Güvenilir XF tahlil veri kümeleri elde etmek için gereken çoğaltma sayısına ilişkin öneriler. Bu Dosyayı indirmek için lütfen tıklayınız.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Ana bulgular ve çıktılar
Bu makale, XFe96 XF Analyzer ile bir dizi kanser kaynaklı hücre hattı kullanarak tek 3D sferoidlerin mitokondriyal enerji metabolizmasını araştırmak için ayrıntılı bir protokol sunmaktadır. Zorla toplama için hücre kovucu teknolojiler kullanılarak A549, HepG2 / C3A, MCF7 ve SK-OV-3 hücresel sferoidlerin hızlı bir şekilde yetiştirilmesi için bir yöntem geliştirilmiş ve tanımlanmıştır. Bu protokol, (1) küresel kültür protokollerinin optimizasyonu ve sferoidlerin orijinal kültürleme kaplarından teknoloji üreticisinden spesifik sferoid tahlil mikroplakalarına taşınması ve aktarılması; (2) Kullanılacak solunum bileşiklerinin konsantrasyonu ve bileşik penetrasyonunun zamana bağımlılığı; (3) Kullanılacak enjeksiyon stratejileri; ve (4) deney grupları arasındaki verileri normalleştirmenin yolları. Tüm bu hususlar mevcut makalede incelenmiş ve aşağıda daha ayrıntılı olarak tartışılmıştır. Bu yöntemler, XFe96 Flux analizörü ile tek 3D sferoidler kullanarak tutarlı metabolik oksijen akısı verileri üretmek için basitleştirilmiş yaklaşımlar olarak sunulmaktadır. Bu deneysel yaklaşım, temel bir laboratuvar ortamında kolayca uygulanabilen diğer küresel modellerde kullanılmak üzere bir başlangıç noktası ve değerlendirme listesi olarak kullanılabilir.
Husus -lar
XF teknolojisinin küresel büyümesi, boyutu ve hassasiyeti
XF teknolojisi ile tekrarlanabilir veriler oluşturmak için, belirli bir model için tahlilin karakterize edilmesi ve optimize edilmesi esastır. Bu yaklaşım, temel bir tek katmanlı hücrede nispeten basittir; Bununla birlikte, bu, hücreleri 3D sferoidler olarak yetiştirirken ek zorluklar ortaya koymaktadır. Burada sunulan deneyler sırasında, üreticiden RPMI ortamı kullanım üzerine desteklenmiştir. Bazı hücre hatlarının, yani HepG2 / C3A'nın DMEM büyüme ortamında kültürlendiği belirtilirken, bu nispeten kısa tahliller sırasında (~ 3-5 saat), RPMI-DMEM formülasyonları ile ikame edilmesinin XF analizi üzerinde sınırlı etkisi olmuştur. İki ortamın formülasyonu çok benzerdir ve kullanıcılar, Seahorse RPMI ortamını, hücre kültürü ortamlarının matrisine takviye yoluyla, örneğin artan glikoz, karbonhidrat kaynaklarının daha fazla eklenmesi yoluyla eşleşecek şekilde 'ayarlayabilir'. Tüm XF tamponlarının ve ortamlarının nihai formülasyonu için kritik olan, XF prob kartuş plakası içindeki floresan problara müdahale etmesi muhtemel fenol-kırmızı ve hücre kültürü inkübatörlerinde bulunan CO2 tamponlama eksikliğinden dolayı alkaliniteye yol açacak sodyum bikarbonatın olmamasıdır. Diğer medya ve tamponlar şirket içinde satın alınabilir ve / veya yapılabilir. Örneğin, Krebs Ringer HEPES tamponu, küresel modeller de dahil olmak üzere birçok farklı hücrede solunumu değerlendirmek için kullanılabilecek basit bir tampondur. Bununla birlikte, XF tahlillerinin kullanıcıları, orta / tampon ve takviyesindeki bir değişikliğin genel tamponlama kapasitesini değiştirebileceğini not etmelidir. Bu, kullanıcılar ECAR'ın proton efflux hızına (PER) dönüşümüne izin vermek için ortamın tampon faktörünün değerlendirilmesi gereken ECAR'ı ölçmekle ilgilendiklerinde özellikle endişe vericidir.
XF teknolojisi ile ölçülen hücresel OCR, kuyudaki hücre sayısı sistemin hassasiyeti içinde olduğunda hücre yoğunluğu ile orantılı olduğundan, bu ilişkiyi tek bir 3D sferoidler kullanarak araştırmak önemliydi. Kuyu başına 1.000, 2.000, 4.000 veya 8.000 hücre yoğunluklarında tohumlanmış 4 farklı kanser hücresi hattından kültürlenmiş tek 3D sferoidlerin OCR'sini araştırarak, XFe96 analizörünün farklı hücre tohumlama yoğunluklarından yetiştirilen 3D sferoidler arasındaki mitokondriyal solunum hızındaki değişiklikleri alacak kadar hassas olduğunu gösteriyoruz (Şekil 3). Optimal hücre tohumlama yoğunluğu aralığının ve dolayısıyla OCR'yi araştırmak için 3D sferoidler oluşturmak için sferoid hacmin hücre tipine bağlı olarak farklılık gösterdiğini gösteriyoruz. Bu, OCR ile tohumlama yoğunluğu veya küresel hacim arasındaki doğrusal ilişki ile gösterilir (Şekil 3). A549 ve HepG2 / C3A hücreleri için, OCR duyarlılığı için optimum tohumlama yoğunluğu 1.000 ila 8.000 hücre / kuyu arasındaydı; MCF-7 için 2.000-8.000 hücre / kuyu ve SK-OV-3 hücreleri için 4.000-8.000 hücre / kuyu idi. Bu veriler, XF teknolojisini kullanarak OCR'yi değerlendirirken küresel boyutun optimizasyonunun özellikle önemli olduğunu göstermektedir.
Minimal ve maksimal sferoid hacimler ve bazal OCR ile ilgili dikkat edilmesi gerekenler
Genel olarak, bu deneyler için imalatçı tarafından önerilen ölçülebilir OCR parametreleri için her zaman minimum ve maksimum eşikler olacaktır. XFe96 analizörü için, 20 pmol O 2 / dak / kuyu ve 200 pmol O2 / dak / kuyu arasındaki bazal OCR, sırasıyla alt ve üst sınırlardır. Bu, tek katmanlı hücreler ve sferoidler için geçerlidir ve deneysel modelin bu dinamik OCR aralığında oturduğu yer, mevcut biyolojik materyal miktarına, örneğin tek katmanlı olarak hücre sayısına veya sferoidlerin boyutuna bağlı olacaktır. OCR eşiklerinin burada kullanılan küresel modellerle nasıl elde edildiğine dair bir örnek için Tablo 4'e bakın. Bu verilerin bu ölçümlerden seviye verisi olarak da elde edilebildiği kuyu içindeki oksijen seviyesini kontrol etmek ihtiyatlı olabilir. Bu, kalite kontrol amacıyla her deneyden rutin olarak görülmelidir. Kuyuda oksijen tükenmesi varsa, bu veriler içinde açıkça görülecektir. Bu durumda, deney içindeki ölçüm döngülerinin ayarlanması gerekli olabilir; örneğin, karıştırma adımını, kuyudaki oksijen seviyesinin ölçüm döngüsü içindeki bir sonraki ölçüm periyodundan önce geri kazanılacak şekilde arttırılması. Mümkün olsa da, tarif edilen hücre çizgilerini kullanan tek sferoid deneyler için bunun çok olası olmadığını bulduk.
Hücre dışı akı tahlilleri için mitokondriyal uncouplers seçimi
Karbonil siyanür 4-(triflorometoksi) fenilhidrazon (FCCP)21, karbonil siyanür m-klorofenil hidrazon (CCCP)22 veya BAM1523 gibi proton iyonoforları, mitokondriyal membranlar boyunca elektrokimyasal proton gradyanını bozabilen, ATP üretimini inhibe edebilen ve nihayetinde mitokondriyal solunumu ayırabilen güçlü küçük moleküllü kimyasallardır 24 . Bu amaçlar için, özellikle metabolik hastalıkların tedavisinde yeni küçük moleküller geliştirilmeye devam etmektedir25,26,27; iki mükemmel incelemeye bakın28,29. Tersine, oksidatif solunumun ayrılması, istenmeyen hedef dışı toksisite30 ile ilişkilendirilmiştir. Bununla birlikte, in vitro hücresel tahliller içinde, FCCP molekülü mitokondriyal membran potansiyelini depolarize eder ve plazma membran depolarizasyonu, NA + iyon akısı31'i bozma gibi hedef dışı etkiler uygular; hücresel protein işleme32 ile girişim ve hatta hücresel yaşlanmayı indükleyen33. BAM15 ilk olarak 2013 yılında plazma membranları 23 üzerinde minimum etkiye sahip bir mitokondriyal uncoupler olarak tanıtıldı, protonoforik aktivite tüm hücrelerde mikromolar aralıkta ve izole mitokondri23,34'te nanomolar aralıkta.
FCCP'nin plazma membran depolarizasyonu üzerindeki gücü göz önüne alındığında, BAM15, hücre dışı akı tahlillerinde sağlam bütün hücrelerde solunumu ayırmak için daha güvenilir bir protonofordur. FCCP ve muadili CCCP, maksimum solunum kapasitelerini test etmek için 50 yılı aşkın bir süredir kullanılmasına ve XF çalışmalarında yaygın olarak kullanılmaya devam etmesine rağmen, bu küçük moleküllerin kullanımı genellikle mitokondriyal ve hücresel metabolik kapasiteyi hafife almaktadır. Bu kısmen, XF teknolojisini kullanan bu kadar çok yayının neden negatif yedek solunum kapasitelerini bildirme tuzağına düştüğü veya FCCP kullanıldığında gerçek mitokondriyal solunum kapasitelerini hafife aldığı ile bağlantılıdır. FCCP'nin sağlam hücrelerde ve dokularda ilave gücü genellikle mitokondriyal fonksiyonun tehlikeye girmesine neden olur ve hücreler, çok düşük konsantrasyonlarda bile, ilavelerini takiben çoklu ölçüm döngüleri boyunca maksimum solunum kapasitesini sürdürmek için uygun şekilde çalışmak için mücadele edebilir35. Bu nedenle, hücrelerin FCCP'ye tepkisi, ilk ölçüm döngüsü periyodunu takiben düşmek üzere birçok çalışmada bulunabilir. FCCP, XF analizi için rutin olarak kullanılırken, BAM15, 10 μM3'e kadar yüksek konsantrasyonlarda tamamen depolarize mitokondride maksimum solunum kapasitesini koruyabilmesi nedeniyle, tüm hücreleri veya küresel modelleri içeren durumlarda tercihen kullanılır. Dahası, BAM15, FCCP 3'ten daha büyük ölçüde HCO3- ve H + 'yı oluşturmak için CO2'nin hidrasyonu yoluyla besin oksidasyonu ile çakışan hücre dışı asitleşme üzerinde etkilere nedenolur. Bununla birlikte, izole mitokondri ve geçirgenleştirilmiş hücreler durumunda, bu uncoupler'lardan herhangi biri, doğru konsantrasyonda titre edilirse, mitokondriyal uncoupling için BAM15'in yanı sıra performans göstermelidir.
Bileşik penetrasyon ve tahlil döngüsü kinetiği
XF analizörünü kullanarak 3D sferoidlerle tipik bir MitoStress testi yapmak için kullanılan kimyasal bileşiklerin konsantrasyonları, penetrasyonu ve kinetik profillerinin ele alınması daha karmaşıktır. Sferoidlerin 3D yapı gösterdiği göz önüne alındığında, moleküllerin sferoidin çapı boyunca nüfuz etmesi, hücre tek katmanlarından sonsuz derecede daha karmaşık bir süreçtir. Örneğin, kinetik penetrasyon ve dolayısıyla kemoterapötik sorafenib'e duyarlılık, küresel yaşa ve dolayısıyla bir HepG2 sferoid model36'daki boyuta göre belirlendi. Küçük moleküllü kimyasalların (örneğin, ilaçlar, nanopartiküller) biyolojik bir hedefe ulaşma yeteneği, sistemin37,38 boyunca dinamik olarak nüfuz edilmesi ve yayılması da dahil olmak üzere çeşitli temel faktörlere bağlıdır. Bu özellikle tümör dokusunu hedef alan ilaçlar için geçerlidir39. Bir 3D sferoid, boyut, kompaktlık ve ilaç taşıyıcı proteinlerin ekspresyonu gibi diğer fenotipik tepkiler bağlamında tümör hedeflemesine benzer şekilde, biyolojik bir yanıt ortaya çıkarmak için gereken bir bileşiğin penetrasyon süresini ve konsantrasyonunu yönetebilir.
Bu protokolde, ATP sentaz inhibitörü oligomisin, protonofor ve mitokondriyal uncoupler BAM15 ve Kompleks I ve Kompleks III inhibitörleri rotenon ve antimisin A'nın kombinasyonuna yanıt olarak penetrasyon süresi ve küçük molekül konsantrasyonu ile ilgili sorunu ele aldık. Bu yaygın solunum bileşiklerinin çoklu titrasyonlarına maruz kalan tek MCF-7 sferoidlerinin OCR'sini araştırarak, kararlı hal solunum hızını indüklemek için gereken her bileşiğin optimal konsantrasyonunun, tek katmanlı hücrelerinkine benzer bir aralıkta olduğunu gösteriyoruz (Şekil 4). Önemli olarak ve tek katmanlı muadillerinden farklı olarak, enjeksiyonlar arasındaki ölçüm döngülerinin sayısının arttırılmasının, tek bir 3D sferoidlerde kararlı durum OCR'si elde etmenin anahtarı olduğu gösterilmiştir. Bu veriler, bu yaklaşımları kullanarak 3D sferoidlerin mitokondriyal solunum parametrelerini araştırırken bileşik penetrasyonun ve ilgili kinetik profillerinin önemini vurgulamaktadır. Şekil 3, Şekil 4 ve Şekil 5'te sunulan verilerle bilgilendirilen küresel optimizasyon özellikleri, bileşiklerin konsantrasyonları ve ölçüm döngüsü süreleri kullanılarak, bir dizi kanser kaynaklı 3D sferoidde mitokondriyal oksidatif metabolizmanın spesifik parametrelerini araştırmak için doğrulanmış bir MitoStress testi oluşturulmuştur (Şekil 6). Önemli olan ve bazı tek katmanlı kanser hücre hatları40 gibi, bazı kanser kaynaklı 3D sferoidlerin maksimum solunum kapasitesi (bağlanmamış-uyarılmış solunum hızı) oligomisin tarafından inhibe edildi (Şekil 5). Spesifik olarak, A549 veya MCF-7 hücrelerinden yetiştirilen 3D sferoidler, oligomisin enjeksiyonunu takiben BAM15 ile ayrıldığında, oligomisin içermeyen BAM15 ile ayrılmaya kıyasla önemli ölçüde daha düşük bir maksimum solunum hızı göstermiştir (Şekil 5F ve Şekil 5H). Bu etkinin diğer 3D sferoid kültürlerde mevcut olabileceği göz önüne alındığında, daha önce doğrulanmış bir protokol kullanılmadığı sürece, 3D sferoidlerdeki maksimum solunum kapasitesinin oligomisin olmadan tahmin edilmesi gerektiğini öneriyoruz.
Hücresel sferoidlerde glikolitik akının bir ölçüsü olarak ECAR verilerinin eşzamanlı toplanması
Literatürde veya teknoloji üreticisinden gelen bilgilerde tipik olarak görüldüğü gibi, ECAR olarak ölçülen sferoidlerin glikolitik oranı, OCR ile birlikte yakalanabilen ikincil bir parametredir. Tek başına ECAR'ın hesaplanması, XF tahlil tamponunun tamponlama kapasitesi veya CO2'nin HCO3- ve H + 'ya hidrasyonundan kaynaklanan mitokondriyal asitifikasyonun eklenmesi için düzeltilmediğinden, herhangi bir XF deneyinde yararlı veya anlamlı bir parametre değildir. ECAR ancak bu veri düzeltmeleri uygulandıktan sonra anlayışlıdır, bundan sonra glikolitik akı hakkında daha doğru sonuçlar vermek mümkün hale gelir. Arabelleğe alma kapasitesinin daha anlamlı PER verileri üretmesini düzeltmek için, küresel mikroplaka için mikro odanın hacmini bilmek gerekir. Üretici, küresel mikroplaka ile bunun için gerçek bir hacim sağlayamamıştır ve bu nedenle PER verileri kolayca belirlenememektedir. Gerçekten de, bu ölçümler ampirik olarak elde edilebilse de, bu bu makalenin kapsamı dışındaydı. Bununla birlikte, uygun düzeltmeler ve kuyuda bulunan belirli bir küresel boyut için mikro odanın hacmini bilerek (örneğin, bir küresel yoğunluk ölçüsü elde etmek), ECAR verileri anlamlı hale gelecek ve glikolitik PER hesaplamaları yapılabilecektir. Bu nedenle, XF verileri daha sonra sferoidlerde glikolitik ve oksidatif metabolizmayı araştırmak için daha bilgilendirici olabilir, ancak yalnızca bu parametreler derinlemesine düşünülürse.
Küresel oluşum, taşıma, transfer ve hareket
Bazı hücre hatları sferoidlerin oluşumu için diğerlerinden daha uygundur ve hiç sferoid oluşturmayabilir, örneğin, MCF-7 yumurtalık kanseri hücreleri41,42, diğer hücre hatlarına kıyasla oldukça dairesel sferoidler oluşturur (Şekil 3). Başka bir örnek olarak, Capan-1 pankreas kanseri hücrelerinin Panc-1 veya BxPC343'ten daha iyi sferoidler oluşturduğu gösterilmiştir. Benzer şekilde, hepatik karsinom hücre hatlarının, kompakt sferoidler 5,44 oluşturmak için değişken yeteneklere sahip olduğu bilinmektedir; HepG2'ye karşı HepG2 / C3A9,45,46 veya HepaRG sferoidleri17,47,48'e karşı olduğu gibi, gelişmiş ilaç metabolizması veya albümin üretimi gibi fenotipte gözlenen bir değişiklikle birlikte. . Bu nedenle, kullanıcılar küresel kültürleme tekniklerini buna göre optimize etmeli ve optimum tohumlama yoğunluğunu ve yetiştirme süresi seyrini belirlemek için titrasyon deneyleri yapmalıdır. Ek olarak, tahlil ortamının formülasyonu ve bileşiminin, matris viskozitesini arttırmak için sıklıkla ortama eklenen metilselüloz ilavesi de dahil olmak üzere sferoid formülasyonu etkilediği gösterilmiştir43,49,50. Bu nedenle, kullanılan tüm hücre hatları için optimal hücre ortamı bileşimi ampirik olarak belirlenmelidir.
Küresel kültür boyunca orta değişimlerin sayısı, kullanılan hücre hattı tarafından belirlenir. Bununla birlikte, tipik olarak, besinleri yenilemek için çoğu durumda her 2-3 günde bir yarım hacimli orta değişimler uygulanabilir. XF analiz çalışmalarında sferoid modellerin hızlı geliştirilmesi ve konuşlandırılması için ticari olarak temin edilebilen kaynaklardan hücre itici mikroplakalar kullanarak 3D sferoidler üretmek için zorla agrega yaklaşımını kullandık. Bununla birlikte, alternatif platformlar, diğer hücre tiplerinden sferoidler üretmek için daha uygun olabilir, örneğin, asılı-damla veya matrise gömülü yaklaşımlar. Kaynak sınırlı laboratuvarlarda, kullanıcılar ilk küresel yöntem geliştirme adımlarının ekonomik maliyetlerini önemli ölçüde azaltmak için hücre itici mikroplaka yüzeylerinin oluşumu için agaroz-sıvı kaplama tekniğine51,52 bakmak isteyebilirler. Sferoidlerin kültür damarları arasındaki hareketi, XF analizi ve diğer aşağı akış tahlillerini gerçekleştirmek için gereklidir. Transfer kolaylığı tipik olarak küresel boyut ve genel yoğunluk tarafından belirlenir. Küresel bütünlüğü korumak için P200 veya P1000 geniş delikli pipet ucu kullanmanızı öneririz; daha küçük delikli pipet uçları, ticari kaynaklı olarak satın alınabilen veya deliği artırmak için pipet ucunun ucunun kesilmesiyle özenle yapılabilen sferoidin mekanik olarak bozulması riskini taşır. Bununla birlikte, bu yaklaşım, ucun ucunun etrafındaki plastiğe tüy sıkma sokmakla yükümlü olabilir ve bu da taşıma sırasında mekanik bozulmaya neden olabilir. Bir arka ışık veya ışık kutusunun kullanılması, sferoidlerin küresel tahlil mikroplakasına başarılı bir şekilde aktarılmasını sağlamak için önemli bir adım olarak diseksiyon mikroskobu altında küresel taşıma ve gözlem için de yararlıdır. Dahası, bir sferoid tahlil mikroplakasının kuyusundaki sferoid pozisyon özel bir öneme sahiptir ve tipik bir MitoStress testi sırasında OCR ve bileşik etkisini doğrudan etkiler (Şekil 7), büyük olasılıkla sferoid pozisyon ve sensör probu floroforlarının ilişkisi nedeniyle.
Arka plan düzeltme ve sıcaklık kontrol kuyuları
Mikroplaka tabanlı tahlillerin kullanımı, çeşitli araştırma alanlarında yaygın olarak kullanılan bir yaklaşımdır; Bununla birlikte, kullanımları çeşitli pratik zorluklar ortaya koymaktadır. Diğer deneysel yaklaşımlarda, özellikle de 96 (veya daha büyük) dizi formatını kullananlarda olduğu gibi, mikroplaka geometrisi ve konumlandırması, genellikle 'kenar etkileri'53,54 olarak adlandırılan zaman içinde plaka boyunca sıcaklık ve gaz değişim gradyanlarını etkileyebilir. Aynı şeyin küresel tahlil mikroplakası için de geçerli olduğunu bulduk. Üreticinin yönergeleri ve protokolleri uyarınca, en dıştaki köşe kuyuları: A1, A12, H1 ve H12, XFe96 analizörü için her zaman arka plan düzeltme ve sıcaklık kontrol kuyuları olarak belirlenmiştir. Tersine, 24 kuyucuklu dizi formatıyla, A1 ve D6, B4 ve C3'te plakanın ortasına eşit olarak yayılmış diğer iki kuyunun yanı sıra kontrol kuyuları olarak da adlandırılır. XF sferoid analizi yaparken, başlangıçta üreticinin rehberliği kullanılarak toplanan verilerde önemli sapmalar bulduk. Bu, verilerin elde edilmesine başlamadan önce sıcaklık ve CO2 içeriğine tahlil ön dengesini sağlamak için gerekli adımların dahil edilmesine rağmen, genellikle bazı solunum inhibitörlerinin enjeksiyonunu takiben OCR için negatif değerler vermiştir (Şekil 8).
Bu gözlemlerin muhtemelen küresel tahlil mikroplakasındaki kenar etkilerinden kaynaklandığını bulduk. Şekil 8'de, arka plan kontrol kuyularının mikro plaka boyunca yeniden dağıtılmasıyla, XF verilerinin yaklaşık 2 kat ayarlandığını bulduk. En olası iki neden, (1) kenar kuyularındaki buharlaşma etkilerinden kaynaklanan, XFe96 probunun numune alması için daha küçük bir toplam hacim odasına neden olan ve (2) arka plan düzeltmesi için belirlenen kuyular ile numune kuyuları arasındaki yetersiz sıcaklık dengelerinden kaynaklanır ve OCR'yi maskeleyen veya aşırı şişiren veri kümeleriyle sonuçlanır. Bu nedenle, bu tür sonuçlardan kaçınmak için, özellikle küresel analiz bağlamında, kullanıcıların sferoid tahlil mikroplakasının tamamı boyunca arka plan düzeltmesi için belirlenmiş kuyuları yeniden dağıtmaları ve XF verilerini almadan önce tahlillerini önceden dengelemek için gerekli adımları atmaları önerilir.
Verilerin normalleştirilmesi
XF teknolojisi ile tek bir 3D sferoidin mitokondriyal enerji metabolizmasını araştırmak için ayrıntılı bir protokol sağlamanın yanı sıra, bu makale ayrıca 3D sferoidlerle elde edilen mitokondriyal solunum hızı verilerini normalleştirmenin olası yollarını da sunmaktadır. Farklı hücre tohumlama yoğunluklarında kültürlenmiş MCF-7 sferoidleri ile elde edilen solunum hızı verilerini kullanarak (Şekil 3), MCF-7 sferoidlerinden ilk hücre tohumlama yoğunluğuna, sferoid hacme ve dsDNA içeriğine normalleştirildiğinde artan boyut ve çaptaki bazal mitokondriyal solunum hızlarını sunuyoruz (Şekil 9). Uygun normalleştirme yöntemi, özellikle in vitro 3D sferoid modelleri ve farklı hücre tiplerini karşılaştırırken, XF veri kümelerinin doğru yorumlanması için çok önemlidir. Kötü normalleştirme, veri kümeleri arasında karşılaştırılamayan hatalı sonuçlara yol açabilir. Küresel XF verilerinin normalleştirilmesi için protein içeriği tercih edilmez, çünkü ön işlemler solunum hızı üzerinde önemli bir etkisi olmadan protein sentezi oranlarını etkileyebilir. Dahası, önemli, tutarsız miktarlarda protein, hücre lizisi üzerine küresel mikroplakalara bağlanabilir ve kuyucuklar arasındaki protein içeriğinde varyasyona neden olabilir. Bu, protein içerebilen, bağlanmak için biyomoleküler yapıştırıcılar gerektiren sferoidler veya yapışkan olmayan hücreler kullanılarak XF analizlerinde daha da karmaşık olabilir.
Hücre içi protein içeriğinin aksine, nükleer DNA içeriği hücre tipinden bağımsızdır ve hücre sayısıyla orantılıdır (Şekil 9D) - hücre sayısı niceliği için sferoidlerin ayrıştırılmasından daha doğru ve daha az zaman alıcı bir yaklaşım. Tersine, fibroblast hücrelerinin tek katmanlarında XF analizleri yapan Yepéz ve ark.55, XF verilerinin hücre sayısına normalleştirilmesinin, normalleştirmeden önce olduğundan daha fazla veri dağılımı sağladığını bulmuşlardır. Nükleer DNA içeriği, farklılaşmış durumdan veya fenotipten bağımsızdır ve bu nedenle XF tahlillerinde küresel verilerin normalleştirilmesi için protein içeriğinden daha doğrudur. DNA içeriği ayrıca metabolizmaya bağlı diğer veri kümelerinin analizi için kanıtlanmış bir strateji olmuştur56. Bununla birlikte, nükleer DNA içeriğinin, sferoid içinde bulunan tüm hücrelerden ölçüldüğünü not etmek önemlidir; Bu nedenle, sferoidlerin hücre canlılığının önemli ölçüde kaybedilmesine neden olabilecek tedavilere tabi tutulduğu XF veri kümeleri için DNA içeriğine normalizasyon önerilmez. Bu tür veri kümeleri için, eğer mümkünse, hücre canlılığına normalleştirme tercih edilir veya veriler bazal solunuma taban çizgisi düzeltilebilir.
Veri normalizasyonunun önemi için yedek solunum kapasitesinin örnek olarak kullanılması
Yedek solunum kapasitesi, maksimal mitokondriyal solunum kapasitesi eksi bazal mitokondriyal solunum hızının bir ölçüsüdür (Şekil 6). Bununla birlikte, bu tür verilerin bir oran olarak raporlanmasıyla ilgili sorun, yani belirli deneylerde pmolO2 / dak / kuyu, verilerin normalleştirmeden geçersiz olmasıdır. Sferoid verileri hücre yoğunluğuna / DNA içeriğine normalleştirse bile, bu genellikle hücrelerdeki mitokondriyal yoğunluk için normalleştirilmesi gereken anahtar parametreyi dışlar. Mitokondriyal yoğunluktaki bir değişikliğin bazal ve maksimum solunumda orantılı bir değişikliğe yol açacağı göz önüne alındığında, yedek kapasite de artacaktır. Örneğin, küresel OCRbazal 200 ve OCRmax 400 ise, yedek kapasite 200 olarak bildirilir; OCRbazal 100 ve OCRmax ise, yedek kapasite de 100'dür; Bununla birlikte, yüzde olarak, her ikisi de maksimumun% 50'sidir (veya bazalin% 100'ü). Bu nedenle, yedek kapasite, pmolsO 2 / dak / kuyu olarak hesaplandığında 200 ve 100 oranlarındaki farklılıklara rağmen, bu iki örnek arasında değiştirilmez. Dahili olarak normalleştirilmiş değerler, XF verilerini çalışmalar ve projeler arasında daha karşılaştırılabilir hale getirmek için daha güvenilir ve anlayışlıdır. Bunu yedek solunum kapasitesi için yapmak için, bunu mutlak bir oran yerine maksimum solunum yüzdesi olarak sunmayı seçtik. Bu aynı zamanda bazal solunumun bir yüzdesi olarak da sunulabilir. Hücrelerle veya sferoidlerle çalışırken durum böyle olurdu. Bununla birlikte, sferoidin mikrovel plakasındaki yerinin mutlak OCR'yi değiştirebileceği, ancak inhibitörler veya unkuplörlerle göreceli değişiklikleri değiştiremeyeceği göz önüne alındığında, sferoidlerde dahili olarak normalleştirilmiş yanıtlara kıvrım değişimi veya yüzdeler olarak bakmak daha önemlidir.
Burada üretilen küresel modeller, klasik 2B modellerde yakalanamayan bir dizi hücre tipi ve mimarisi sunar. Bunlar, hücrelerin üç boyutlu heterojen, uzamsal düzenini, gelişmiş hücre-hücre temaslarını (örneğin, boşluk bağlantılarının ve hücre dışı matrislerin oluşumunu) ve küresel çap boyunca biyokimyasal gradyanları (örneğin, pH gradyanları, besin maddelerine oksijen difüzyonuna erişim) içerir. İn vitro sferoid biyolojiyi incelemek için hücre dışı akının kullanılması, ilaç tedavileri için optimal hedeflerin metabolik pertürbasyon gözlemleri yoluyla tanımlanmasına izin verebilir. Bunlar in vitro sferoidlerden in vivo tümörlere ekstrapolasyon yapılabilir ve sferoid tümör metabolizmasını, örneğin sferoid büyüme sırasında karbonhidrat kullanımını hedefleyebilecek yolları tanımlayabilir. Terapötik modaliteler, erken büyüme evrelerinde sferoidlerin hedeflenmesinde etkili olabilir, ancak metabolik ağ karmaşıklığı olgunlaştıkça sferoid büyümenin sonraki aşamalarında daha az etkili olduğu kanıtlanmıştır. Sonuç olarak, biyolojik araştırmalarda 3D hücre kültürü modellerinin ve sofistike analiz teknolojilerinin geliştirilmesi, eşsiz potansiyele sahip dinamik ve hızla değişen bir alan olmaya devam edecektir. İn vitro hücre kültürü sferoidlerinin hücre dışı akı analizi, insanla ilgili biyolojiyi daha iyi anlamak, araştırmada hayvan modellerinin kullanımını azaltmak ve hasta merkezli araştırmaları geliştirmek için tahmin edilebilecek araştırma sonuçlarını ilerletmek için en yeni araştırma yöntemi olarak kullanılabilir.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarların beyan edecekleri herhangi bir çıkar çatışması yoktur.
Acknowledgments
N.J.C, Sygnature Discovery Ltd ile BBSRC MIBTP CASE Award tarafından desteklenmiştir (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Kanser Araştırmaları Sayı 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
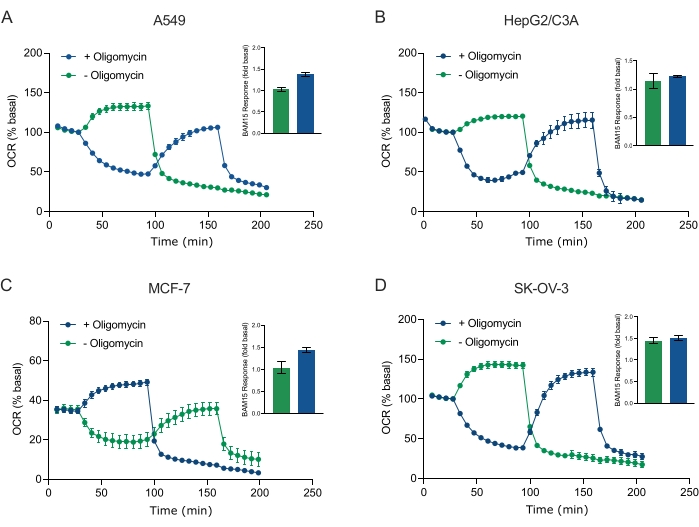
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
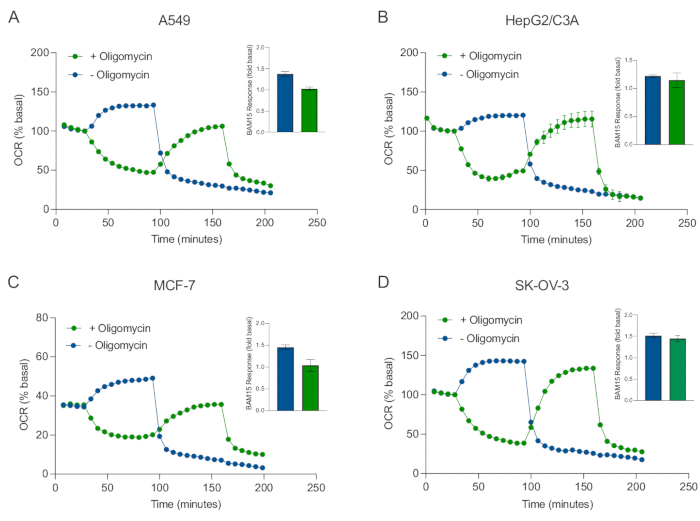
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
