Phase contrast images of mouse endometrial organoids
We established organoids from WT mouse endometrial epithelium, as described in the attached protocol (see diagram in Figure 1). Following enzymatic dissociation of the mouse endometrial epithelium, epithelial sheets were mechanically separated from the uterine stromal cells and further dissociated with collagenase to generate a single-cell suspension. If performed correctly, this method of epithelial and stromal cell separation should yield samples with contamination of no more than 10%-15% of the opposite cell type (see immunofluorescence images in Figure 2). The epithelial cell pellet was then resuspended in gel matrix, allowed to gel at room temperature, and plated into 25 µL domes on tissue culture plates. After the gel matrix domes solidified, they were overlaid with organoid medium and allowed to grow. We typically observed that endometrial epithelial cells assembled into organoids within 3-4 days, as pictured in Figure 3. The organoids were maintained in culture, with media changes occurring every 3 days, and passaging every 5-7 days.
Imaging of mouse endometrial organoids using histological and immunofluorescent staining
To analyze the morphology of the mouse epithelial organoids, we adapted methods used for the encapsulation of cellular fine needle aspirates using specimen processing gel20. This allows for the preservation of the endometrial organoid morphology and encapsulation into a matrix that can be subjected to processing in preparation for paraffin embedding. Specimen processing gel is a modified agar that is widely used in clinical pathology laboratories for the analysis of fine needle aspirates and has been used to encapsulate vaginal organoids21,22. After the specimen processing gel/organoid mixture solidifies, it can be processed, stained, and imaged with techniques typically used to analyze tissues. In Figure 4A,B, we show that sectioned formalin-fixed paraffin-embedded endometrial organoids can be visualized by hematoxylin and eosin stains, which display the single layer of epithelium and hollow centers-the lumens-of the organoids. Certain organoids also contain secretions in the lumen, indicating that the organoids acquire the functional properties of glandular epithelial cells. We also describe techniques to visualize the organoids using immunostaining (Figure 4C,D); sectioned endometrial organoids were incubated with a Cytokeratin 8 primary antibody (TROMA-1) and a fluorophore-conjugated secondary antibody (secondary anti-rat-594). Nuclei were then visualized with DAPI, and the slides were imaged using a fluorescence microscope. We observed that all the cells in the organoids were Cytokeratin 8-positive and contained DAPI, indicating that fixation, embedding, and processing of the endometrial organoids is compatible with antibody-based immunostaining.
RNA extraction and gene expression analysis
To determine the gene expression response of endometrial organoids, we allowed endometrial organoids from WT mice to grow for 4 days after the first passage. On the evening before treatment, the endometrial organoid medium was changed to starvation medium. The organoids were then treated in triplicate wells (each well containing three domes) with vehicle (ethanol; equal volume in solution as used for estradiol) or 10 nM estradiol (E2) for a total of 48 h. After 48 h, the medium was removed, and the organoids were processed for RNA extraction. Approximately 4 µg of RNA can be obtained from a total of two domes from each well, and 1 µg of RNA was used to reverse transcribe cDNA. Amplification of the genes encoding lipocalin 2 (Lcn2), lactoferrin (Ltf), and the progesterone receptor (Pgr) was performed using real-time quantitative PCR (Figure 5 and Table 1)23,24,25. As expected, we observed that the expression of Lcn2, Ltf, and Pgr increased in the epithelial organoids following stimulation with 10 nM E2 (Figure 5). Thus, these results show that endometrial epithelial organoids can be successfully used to measure the gene expression changes in response to E2.

Figure 1: Procedures used to establish endometrial organoids. Diagram outlines the key steps that were followed to (A) establish and (B) passage epithelial organoids from the mouse endometrium. (A) Methods include the enzymatic and mechanical dissociation of endometrial epithelium from the mouse uterus, followed by single-cell dispersion and encapsulation of the epithelium into a gel matrix. To obtain the uterine tissues for enzymatic dissociation, the ovaries and oviducts are removed from the uterine horns with fine scissors, cut laterally into 4-5 mm fragments, and then placed in the enzyme solution. After enzymatic incubation, the endometrial epithelium is mechanically separated from the underlying stroma under a microscope using forceps and a pipette to "squeeze" out the epithelium from the uterine tube. The epithelium is further digested into epithelial cells using a short incubation in collagenase, followed by encapsulation into a gel matrix. (B) Passaging the endometrial organoids requires mechanical dissociation in cold medium and centrifugation to release the organoids from the matrix and to generate smaller organoid fragments. Once the organoids have been released from the matrix and mechanically dissociated into smaller fragments, they are encapsulated into matrix and replated. Please click here to view a larger version of this figure.
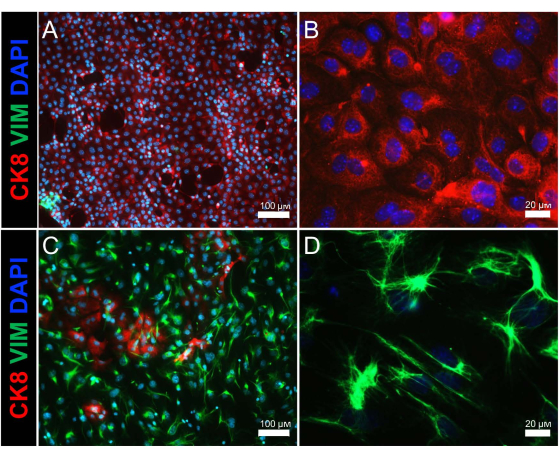
Figure 2: Immunostaining of isolated endometrial epithelium and stromal cell populations. Immunofluorescence images show epithelial and stromal cell populations of the (A,B) epithelial cell and (C,D) stromal cell fractions following enzymatic and mechanical separation of the uterus. Cytokeratin 8 (an epithelial cell marker) is shown in red, vimentin (a stromal cell marker) is shown in green, and DAPI (a nuclear marker) is shown in blue. Scale bars = 100 µm (A,C), 20 µm (B,D). Abbreviations: CK8 = cytokeratin 8; VIM = vimentin; DAPI = 4',6-diamidino-2-phenylindole. Please click here to view a larger version of this figure.

Figure 3: Formation of endometrial organoids from WT mice over the course of 4 days. Endometrial epithelium was isolated from a WT mouse and used to generate organoids. (A) Phase contrast image of the epithelial cells encapsulated in the gel matrix immediately after digestion (day 0). (B,C) A few small organoids can be observed on day 1 (B) and day 2 (C) of culture. (D) Larger and mature epithelial organoids can be observed on day 3 of culture. Images were captured with a 5x objective; scale bars = 200 µm. Abbreviation: WT = wild type. Please click here to view a larger version of this figure.

Figure 4: Histological analysis of endometrial organoids. (A,B) FFPE endometrial epithelial organoids were sectioned and stained with H&E. (C,D) FFPE epithelial organoids were immunostained with the epithelial cell marker cytokeratin 8 (red). Cell nuclei were stained with DAPI (blue). Scale bars = 200 µm (A), 100 µm (C), 50 µm (B,D). Abbreviations: FFPE = formalin-fixed paraffin embedded; H&E = hematoxylin & eosin; CK8 = cytokeratin 8; DAPI = 4',6-diamidino-2-phenylindole. Please click here to view a larger version of this figure.

Figure 5: Gene expression analysis of mouse endometrial organoids. Mouse endometrial epithelial organoids from WT mice were cultured for 4 days in organoid growth medium, followed by treatment with vehicle or 10 nM E2 for 48 h in DMEM/F12 supplemented with 2% charcoal-stripped FBS. (A,B) Phase contrast images of the WT mouse organoids after treatment with vehicle (A) or 10 nM E2 (B). (C–E) Real time qPCR analysis of epithelial endometrial organoids treated with vehicle or 10 nM E2. Expression of E2-regulated genes, lipocalin 2 (Lnc2), lactoferrin (Ltf), and progesterone receptor (Pgr). Bars represent mean ± SEM, analyzed by a two-tailed t-test; *p < 0.033, **p < 0.002, ***p < 0.001. Repeated with organoids derived from n = 3 WT mice per condition. Scale bars = 200 µm (A,B). Abbreviations: WT = wild type; E2 = estradiol; qPCR = quantitative PCR. Please click here to view a larger version of this figure.
| Primer Sequence | Forward (5'-3') | Reverse (5'-3') |
| Lipocalin 2 (Lcn2) | GCAGGTGGTACGTTGTGGG | CTCTTGTAGCTCATAGATGGTGC |
| Lactoferrin (Ltf) | TGAGGCCCTTGGACTCTGT | ACCCACTTTTCTCATCTCGTTC |
| Progesterone (Pgr) | CCCACAGGAGTTTGTCAAGCTC | TAACTTCAGACATCATTTCCGG |
| Glyceraldehyde 3 phosphate dehydrogenase (Gapdh) | CAATGTGTCCGTCGTGGATCT | GCCTGCTTCACCACCTTCTT |
| PCR mixture contains: |
| 2x SYBR Green |
| 0.5 µM Fwd Primer |
| 0.5 µM Rev Primer |
| cDNA |
| RNAse/DNAse-Free H2O |
| Cycler Conditions: | |||
| Temperature | Time | Cycles | |
| Hold | 95 °C | 10 min | 1 |
| Denature | 95 °C | 15 s | 40 |
| Extend | 60 °C | 60 s |
Table 1: Primer sequences and PCR conditions.
