

Summary
Xenopus эмбриональной эктодермы стала привлекательной моделью для исследования клеточной полярности. Анализа описывается, в котором субклеточных распределение флуоресцентных белков оценивается в эктодермы клеток. Этот протокол поможет решить вопросы, связанные с пространственным управления сигнализацией.
Abstract
Сотовые полярность фундаментальное свойство клеток эукариот, который динамически регулируется как внутренних и внешних факторов во время эмбрионального развития 1, 2. Один из сигнальных путей, участвующих в этом регулирование Wnt путь, который используется во много раз во время эмбриогенеза и критические для человека болезнь 3, 4, 5. Множественные молекулярные компоненты этого пути согласованно регулировать сигнализации в пространственно-ограниченной форме, но основные механизмы, полностью не поняты. Xenopus эмбриональных эпителиальных клеток является отличной системой для изучения субклеточных локализации различных сигнальных белков. Флуоресцентные белки слияния выражены у эмбрионов Xenopus путем микроинъекции РНК, эктодермального эксплантов готовятся и локализации белка оценивается epifluorescence. В этом протоколе исследования мы описываем, как субклеточные локализации Diversin, цитоплазматических белков, которые были причастны к сигнализации и клеточной полярности определения 6, 7 визуализируется в Xenopus эктодермальных клеток для изучения передачи сигналов Wnt 8. Коэкспрессия Wnt лиганда или Frizzled рецепторов изменяет распределение Diversin сливается с красного флуоресцентного белка, ППП, и новобранцы его клеточную мембрану в поляризованной моды 8, 9. Это бывший протокол естественных условиях должно быть полезным дополнением к лабораторные исследования в культивируемых клетках млекопитающих, в котором пространственные контроль сигнализации отличается от неповрежденной ткани и является гораздо более трудной для анализа.
Protocol
1. Экстракорпоральное оплодотворение яйца из Xenopus
- Получить яйца самка лягушки, которые вводили хорионический гонадотропин человека (400 ед / лягушка) 12 часов до эксперимента.
- Место яйца в небольшом количестве (0,5-1 мл) на 1 изменение х Марка раствор Рингера (MMR) 10 к яйцам и оплодотворяют их в пробирке с небольшой фрагмент расчлененного яичка. Через 2-3 мин, добавить 0,1 х MMR, чтобы покрыть всю поверхность яйца. Через 20 минут, яйца желе слой удаляется на 3% цистеина - HCL (доводят до рН 8 с гидроксидом натрия). Яйца моют с 0,1 х MMR в три раза и оставили в холодном инкубатор (13 ° С) для инъекций.
- Оплодотворенные яйца позволили разработать до 2-4 клеточной стадии. Для инъекций, эмбрионы переносят в раствор, содержащий 3% Ficoll, 0,5 х MMR.
2. РНК Микроинъекция
- РНК синтезируются из линеаризованной ДНК с использованием шаблонов mMessage mMachine комплект (Амбион) и разбавляют РНКазы свободной воды на фондовом концентрации 0,1-1 мкг / мкл. Оптимальные дозы РНК для инъекций определяются в пилотных экспериментах. РНК для Diversin-RFP, мембранные маркера GFP-CAAX и Frizzled 8 используются в 0,1-1 нг на инъекцию.
- Инъекция иглы готовы с иглой съемник из капилляра, а затем с помощью иглы мясорубки. Перед инъекцией, каждая игла откалиброван с водой, чтобы извлечь 10 п жидкого на инъекцию.
- Для инъекций, эмбрионы помещаются в пластиковый блюдо в большую каплю 3% Ficoll, 0,5 х MMR. Один микролитр решение РНК всасывается в инъекционной иглой с Narishige microinjector. 10 п РНК вводят животным бластомеров от 8 клетка эмбриона 2-3 раза. Вводят эмбрионы переносятся в скважине 12-луночного планшета.
3. Подготовка эктодермальная эксплантов
- Когда вводят эмбрионы достигают ранней стадии гаструлы, они переводятся в 0,6 х MMR решение в 3 см пластиковая тарелка с покрытием 1% агарозы. Желточный мембрана удалены вручную с парой щипцов. Эктодермальная эксплантов которые вырезали из эмбрионов с помощью иглы вольфрама и hairloop.
- Эктодермальная эксплантов переносятся в стеклянный пузырек и фиксировали 3,7% формальдегида в фосфатном буферном растворе (PBS) в течение 30 минут. Исправлена эксплантов промывают PBS в три раза (10 минут каждый). DAPI входит в третью мыть для окрашивания ядер.
- Эксплантов установлены на предметное стекло. Две полосы скотчем прикреплены к слайд-шоу и эксплантов помещаются между двух полос. С наружной поверхности эксплантатов это пигментированные, внутренняя сторона должна быть обращена эксплантов объектива микроскопа. Добавить два-три 20 мкл капли решение для монтажа (70% глицерина в PBS в том числе 25 мг / мл DABCO, анти-выцветания реагента) 11. Положите покровное на вершине.
4. Изображений эксплантов Под Флуоресцентный микроскоп
- Образцы смотреть под Zeiss Axioplan флуоресцентный микроскоп с соответствующими фильтрами.
- Изображения взяты с Apotome привязанность к себе конкретный самолет из нескольких независимых эксплантов.
5. Cryosectioning
Cryosectioning это альтернативный способ визуализировать распределение флуоресцентных белков в клетке и больше подходит для иммунной окраски. На этапе 10, эмбрионы фиксированной в течение 1-2 часов с фиксатором Дента (20% ДМСО, 80% метанола), промывали PBS, и встроенных в 15% рыбы gelatin/15% раствором сахарозы 11. Встроенный эмбрионы быстро замороженных в сухом льду и cryosections генерируются на Leica криостат. Сечения будет включать эктодермального клетки, которые наследуют вводили РНК и их перевод белковых продуктов. Разделы сохраняют флуоресценции и может быть immunostained со специфическими антителами, а затем помечены вторичных антител, конъюгированных с флуоресценции. Ядра окрашивали DAPI. Монтаж средств массовой информации такие же, как описано выше. Изображений может быть выполнена как описано выше.
6. Представитель Результаты:
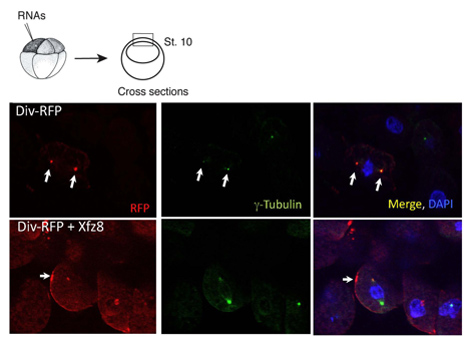
Рисунок 1. Frizzled рецепторов новобранцев Diversin к клеточной мембране. Эктодермы клеток, экспрессирующих Frizzled 8 (Xfz8) и Div-RFP РНК выявить Div-RFP на клеточную мембрану, а не центросомы (как показал г-тубулина совместного окрашивания). Схема эксперимента находится на самом верху; типичный сечение показано на рисунке.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Мы использовали выше протокол, чтобы охарактеризовать субклеточные локализации Diversin. В эксплантатах животных полюса, Diversin-ППП был обнаружен рядом с ядром и colocalized с г-тубулина, centrosomal маркер, в cryosections (рис. 1). После субклеточные локализации белка определяется, удаление конструкции могут быть сгенерированы установить, какие белковых доменов являются необходимыми и достаточными для субклеточные локализации. Используя этот подход, centrosomal области локализации Diversin будут переведены в середине и в карбокси-концевых участков белка, каждая из которых содержит биспиральных мотив 8.
Же протокол может быть использован в исследованиях, в которых белок локализации изменяется в ответ на сигнализацию. Мы обнаружили, что Wnt секретируемых белков действовать, чтобы переместить Div-RFP для точечной структуры, прилегающей к клеточной мембране, тогда как FZ8 новобранцев Div-RFP к клеточной мембране патчи (рис. 1). Кроме того, мы обнаружили, что карбокси-концевой домен не является необходимым для вербовки мембраны как таковой, но необходимые для поляризованных набор мембран.
Таким образом, выше экспериментальный протокол поможет в различных исследованиях белок-белковых взаимодействий и белковых локализации на различных клеточных отсеках после стимуляции клеток со специфическими факторами роста или сигнальных белков.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Нет конфликта интересов объявлены.
Acknowledgments
Исследования в лаборатории Сокол спонсируется Национальным Institues здравоохранения.
References
- Gurdon, J. B. Embryonic induction --- molecular aspects. Development. 99, 285-306 (1987).
- Principles of Developmental Genetics. Moody, S. A. , Academic Press. (2007).
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell. 127, 469-480 (2006).
- Klaus, A., Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat Rev Cancer. 8, 387-398 (2008).
- Gordon, M. D., Nusse, R. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem. 281, 22429-22433 (2006).
- Schwarz-Romond, T., Asbrand, C., Bakkers, J., Kuhl, M., Schaeffer, H. J., Huelsken, J., Behrens, J., Hammerschmidt, M., Birchmeier, W. The ankyrin repeat protein Diversin recruits Casein kinase Iepsilon to the beta-catenin degradation complex and acts in both canonical Wnt and Wnt/JNK signaling. Genes Dev. 16, 2073-2084 (2002).
- Moeller, H., Jenny, A., Schaeffer, H. J., Schwarz-Romond, T., Mlodzik, M., Hammerschmidt, M., Birchmeier, W. Diversin regulates heart formation and gastrulation movements in development. Proc Natl Acad Sci U S A. 103, 15900-15905 (2006).
- Itoh, K., Jenny, A., Mlodzik, M., Sokol, S. Y. Centrosomal localization of Diversin and its relevance to Wnt signaling. J. Cell Sci. 122, 3791-3798 (2009).
- Itoh, K., Jacob, J., Sokol, Y. S. A role for Xenopus Frizzled 8 in dorsal development. Mech Dev. 74, 145-157 (1998).
- Newport, J., Kirschner, M. A major developmental transition in early Xenopus embryos: I. characterization and timing of cellular changes at the midblastula stage. Cell. 30, 675-686 (1982).
- Itoh, K., Brott, B. K., Bae, G. U., Ratcliffe, M. J., Sokol, S. Y. Nuclear localization is required for Dishevelled function in Wnt/beta-catenin signaling. J Biol. 4, 3-3 (2005).
