Thyroid anlages (midline and UB, together with surrounding tissues), and thyroid lobes are dissected from mouse embryos at e12.5 and e13.5/e14.5, respectively (Figure 1). After one day in culture on filter, the midline anlage is visible (Figure 2A) as an elongated tissue spanning on top of the trachea. It progressively forms two lobes on each side of the trachea. If isolated thyroid lobes (e13.5 or e14.5) are cultured, they will expand, undergo morphogenesis and epithelial cells will organize into macroscopically visible follicular structures (Figure 2B).
Organization and polarization of epithelial cells, labeled with E-cadherin, can be visualized by ezrin immunolabeling (Figure 3A). Ezrin-positive intracellular structures progressively, and in a concerted way, fuse with one pole of the cell that will become the apical pole. During this process, endothelial cells positive for PECAM proliferate and organize around the developing follicles to form angio-follicular units (Figure 3B). To quantify differentiation of thyrocytes and C-cells, RNA can be isolated from the cultured thyroid lobes, and gene expression assayed by RT-qPCR (Figure 3C). While the expression of the thyroid transcription factor Nkx2.1 does not change during the culture, expression of thyrocyte-specific thyroglobulin and of C-cell-specific calcitonin dramatically increased, indicating functional differentiation.
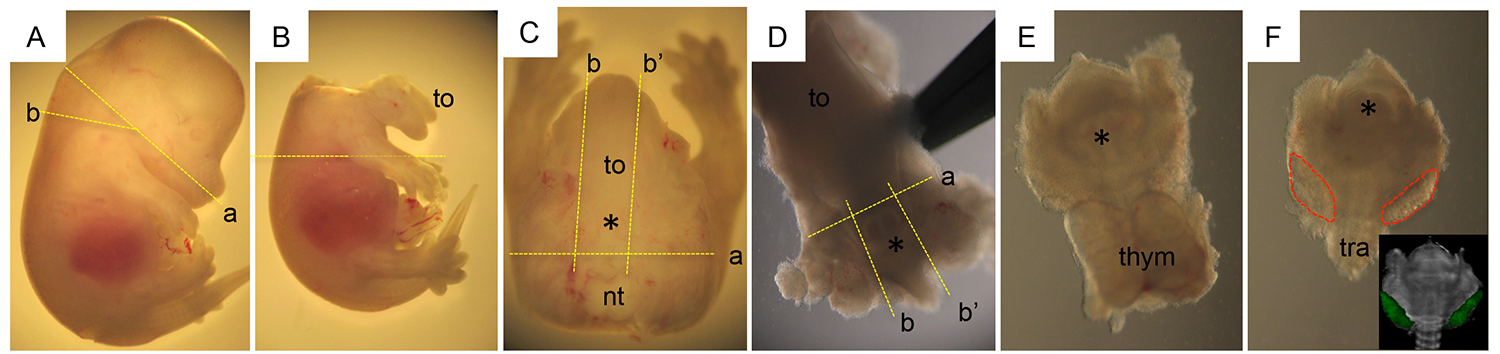
Figure 1. Dissection steps to isolate the thyroid from e14.5 mouse embryo. Incisions are depicted by yellow dotted lines. (A) The upper part of the head is removed by two incisions to expose the tongue. (B) The neck region is separated from the rest of the body by sectioning the embryo below the upper limbs and above the heart. (C) Keeping the tongue (to) as a guide, remove the neural tube (nt) and the lateral tissues and limbs. (D) The tongue, arytenoid swelling (*) and the esophagus/trachea are perfectly aligned. First remove the tongue and then tissues lateral to the esophagus/trachea region. (E) The thyroid lobes are partly hidden by the thymus (thym) which should be discarded. (F) Thyroid lobes are visible on each side of the trachea (red dotted ovoids), or better visualized using fluorescent reporter embryos (Pax8-Cre; Rosa-stop-YFP; inset). Abbreviations: to (tongue), thym (thymus), tra (trachea), * (arytenoid swelling). Please click here to view a larger version of this figure.
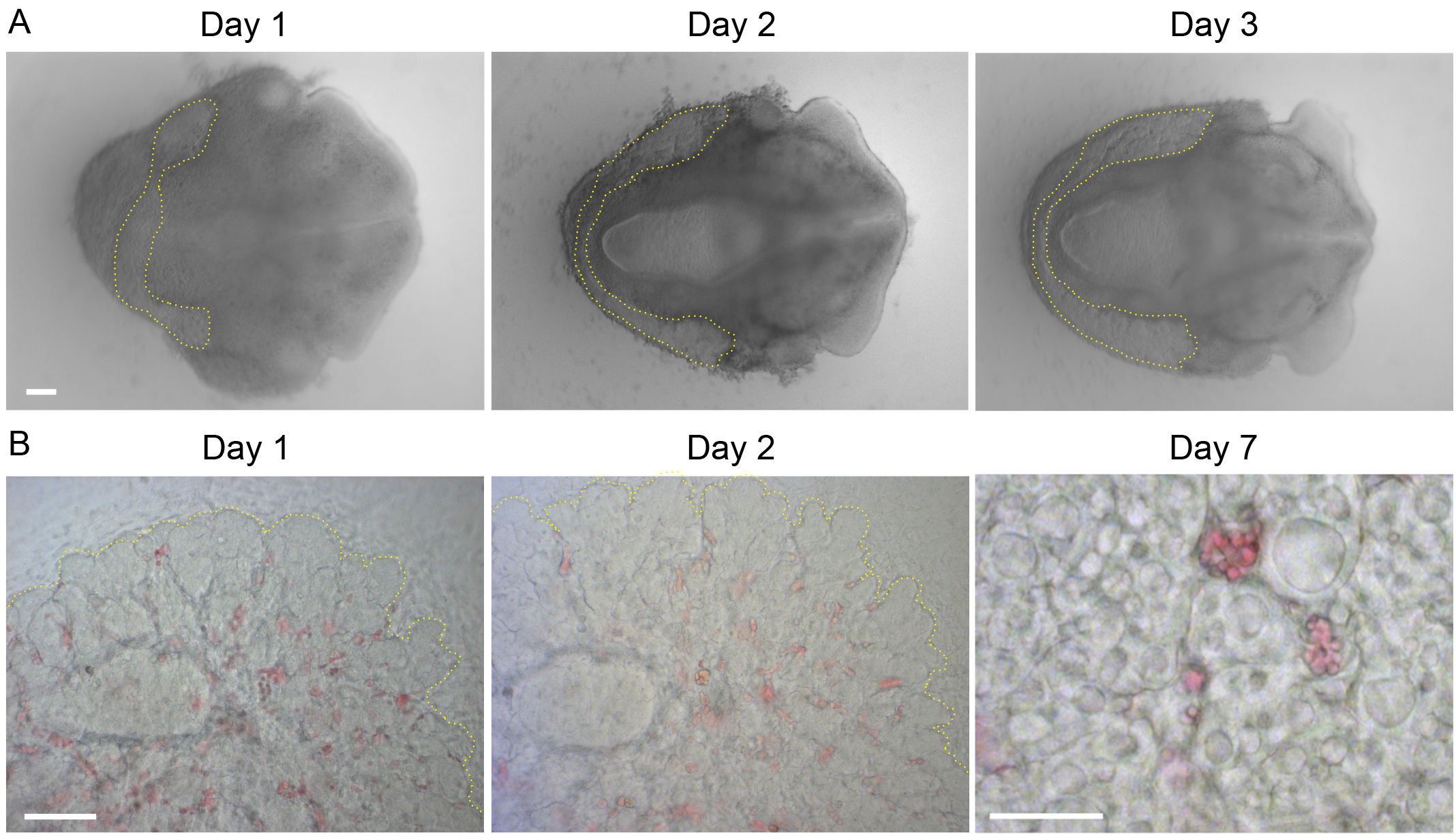
Figure 2. Dissected thyroid explants on filter and thyroid lobes on fibronectin nicely develop ex vivo. (A) Photographs of an e12.5 thyroid explant cultured for 1, 2 and 3 days on semiporous filter, at the air-medium interface. Thyroid epithelial cells migrate bilaterally on each side of the trachea and form two growing thyroid lobes. (B) Brightfield images on day 1 and day 2 of an e14.5 thyroid lobe cultured on microscopy plastic culture chambers. Note the expansion and remodeling of the epithelium that ultimately form large (20-100 μm) follicles around day 7. Yellow dotted line delineates the epithelial periphery of the thyroid. Scale bars, 100 μm. Please click here to view a larger version of this figure.

Figure 3. Folliculogenesis, angiogenesis and differentiation of thyroid in culture. Thyroid lobes were cultured for the indicated time. (A) Fixed tissues were whole-mount immunostained with antibodies against the basolateral marker E-cadherin, and the subapical membrane marker Ezrin. After one day in culture, epithelial cells of the thyroid parenchyma present small ezrin-positive structures that correspond to intracellular vesicles7. Concerted fusion of the vesicles from adjacent cells will form small lumen at day 2; their progressive growth, and organization of the surrounding cells around the lumen will delineate pre-follicular structures after 4 days. Scale bar, 20 μm. (B) Thyroid lobe immunostained with antibodies against the Platelet and Endothelial Cell Adhesion Molecule (PECAM) and Ezrin. Follicles are surrounded by endothelial structures. Scale bar, 50 μm. (C) RNA, extracted from thyroid lobes, was reverse transcribed before quantitative PCR. The expression of the thyroid transcription factor Nkx2.1, and of two differentiation genes, thyroglobulin and Calcitonin, was measured using specific primer pairs. To calculate the relative gene expression, the ΔΔ Ct method was used, using E-cadherin as the reference gene and the last time point of the culture as 100%. Images shown in A and B are single confocal optical sections of whole-mount immunostained thyroid lobes. Please click here to view a larger version of this figure.
