1. hiPSC maintenance culture
- Coat a six-well culture plate with laminin (Table of Materials).
- Dilute laminin to 0.5 μg/mL in phosphate buffered saline (PBS).
- Add diluted laminin to a six-well plate at a volume of 2.0 mL/well.
- Incubate the plate at 37 °C for 30 min.
- Remove the laminin solution from the wells.
- Seed hiPSCs in 2 mL of iPSC maintenance medium (Table of Materials) onto the coated wells at a density of 3 × 104 cells/well without drying the surface.
- Subculture the hiPSCs.
- Prepare 0.5× cell dissociation enzymes (Table of Materials) solution.
- Mix 10 μL of 0.5 M Ethylenediaminetetraacetic (EDTA) and 10 mL of PBS to make 0.5 mM EDTA/PBS.
- Dilute 10 mL of 1× cell dissociation enzymes to 0.5× by adding 10 mL of 0.5 mM EDTA/PBS.
- Aspirate the spent medium from the wells.
- Add 2 mL/well of PBS to wash the cells, then discard the PBS.
- Add 800 μL of the 0.5× cell dissociation enzymes, and incubate the cells for 7 min at 37 °C in a humidified incubator containing 5% CO2.
NOTE: 0.05% trypsin-EDTA can be used to dissociate cells. - Gently remove the 0.5× cell dissociation enzymes solution.
- Wash the cells with 2 mL PBS, then discard the PBS.
NOTE: Be gentle because the cells easily detach after adding cell dissociation enzymes solution. - Add 1 mL of iPS maintenance medium (Table of Materials) containing 10 μM Y-27632.
NOTE: Adding Y-27632 increases the survival rate of hiPSCs. - Dislodge cells using a cell scraper, and collect them in a 15 mL centrifuge tube.
- Count the number of cells. Adjust the cell density to 1.5 × 104 cells/mL by adding iPS maintenance medium containing 10 μM Y-27632.
NOTE: Do not use centrifugation in order to prevent cell damage. - Seed 2 mL of cell mixture on the laminin-coated six-well plate (final density: 3 × 104 cells/well).
- Incubate the cells at 37 °C in a humidified incubator containing 5% CO2.
- Replace the culture medium with iPS maintenance medium without Y-27632 on days 1, 4, 5, and 6.
- Prepare 0.5× cell dissociation enzymes (Table of Materials) solution.
- Subculture the cells on day 7.
NOTE: Subculture the cells by day 7 in order to inhibit random differentiation of hiPSCs.
2. Induction of cardiac differentiation of hiPSCs
- Coat a 96-well culture plate with laminin (Table of Materials).
- Dilute laminin to 1.675 μg/mL with PBS.
- Add the diluted laminin solution to a 96-well plate at a volume of 0.1 mL/well.
- Incubate the plate at 37 °C for 30 min.
- Seed hiPSCs onto a 96-well plate at a density of 3 × 104 cells/well.
- Proliferate hiPSCs at 37 °C in a humidified incubator containing 5% CO2.
- One day later, replace the medium with 200 μL/well iPS growth medium (Table of Materials).
- Change the medium every day for an additional 2–3 days until the cells reach 70%–80% confluency.
- Apply differentiation media.
- Aspirate the spent medium and replace it slowly with 200 μL/well of pre-warmed differentiation medium A (Table of Materials).
- Place the plate at 37 °C in a humidified incubator containing 5% CO2.
- After 48 h, aspirate the medium and slowly replace it with 200 μL/well of pre-warmed differentiation medium B (Table of Materials).
- Place the plate at 37 °C in a humidified incubator containing 5% CO2.
NOTE: It is extremely important that the differentiation media are kept fresh. They will gradually lose their differentiation effect, typically within two weeks. If necessary, aliquot fresh medium and store at −20 °C.
- Apply cardiomyocyte maintenance medium.
- After 48 h, aspirate the medium and slowly replace it with 200 μL/well of pre-warmed cardiomyocyte maintenance medium (Table of Materials).
- Place the plate at 37 °C in a humidified incubator containing 5% CO2.
- Replace the cardiomyocyte differentiation medium every other day up to day 30.
NOTE: It is important that the duration of the application of differentiation media A and B is accurately set to 48 h to ensure the precise expression sequence of genes required for differentiation.
3. Exposure to ischemia
- Deprive the culture medium of nutrients and oxygen.
- Prepare Dulbecco's modified Eagle medium (DMEM) without glucose and serum.
- Aspirate culture medium from the wells of the 96-well plate containing hiPS-CMs.
- Add the nutrient-deprived medium to the wells at a volume of 200 μL/well.
- Place the culture plate in a hypoxic incubator (Table of Materials).
- Infuse nitrogen gas, and maintain the internal oxygen concentration at 2% and CO2 concentration at 5% for 24 h.
- Proceed to desired analyses: e.g., viability assay, assessment of contractility, or assessment of cellular damage.
4. Assessment of cellular viability using MTT assay
- Use the MTT assay kit (Table of Materials) to quantitatively assess the viability of cells.
NOTE: Exposure to 1 mM hydrogen peroxide in DMEM for 1 h can be used to damage cells (positive control). Exposure to 0 mM hydrogen peroxide in DMEM can be used for the negative control.- Add 10 μL of MTT reagent to the cells using a repeating pipettor.
- Mix gently for one minute on an orbital shaker.
- Incubate the cells for 3–4 h at 37 °C in a 5% CO2 incubator. After incubation, the formazan produced in the cells will appear as dark crystals in the bottom of the wells.
- After removing the supernatant, dissolve the insoluble formazan crystals in 100 μL of dimethyl sulfoxide solution (DMSO). This solution will dissolve the formazan crystals, producing a purple solution.
CAUTION: DMSO can irritate eyes, respiratory system, and skin. Wear suitable gloves and eye/face protection. - Measure the absorbance of each sample with a microplate reader at a wavelength of 570 nm.
5. Assessment of contractility of iPS-CMs
- If not installed, obtain Particle Image Velocimetry ImageJ plugin11 at https://sites.google.com/site/qingzongtseng/piv and install.
- Using a phase contrast microscope, record the video images of hiPS-CMs using the 4× objective lens at ~20 frames per second for ~10 s, and save as “analyze.avi”. For a comparison of contractility between before and after ischemia, confirm that the location of interest is recorded.
NOTE: A microscope with an automated stage is useful to target the location of interest. Movie file should be in .avi format for later analysis. If not, convert the movie to .avi. - Create folder structure as indicated in Figure 1. An example of “joblist.txt” is indicated in supplementary file.
- Analyze discrete two-dimensional vector fields of cellular displacement.
- Launch Fiji (ImageJ) software12, go to Plugins > Macros > Edit and open “vector_analysis.ijm” (Supplementary Coding File).
- Click Run. Analysis will be performed automatically.
NOTE: Displacement vectors D(x,y) are calculated for every 16 × 16 pixels between the reference frame (the first frame) and all subsequent frames (frames 1 vs. 2, 1 vs. 3, 1 vs. 4, etc.). The result is calculated for every frame and saved as “vec_x.txt” (x is a frame number). A vector of maximum displacement, M(x, y), is defined for every (x, y) pair as follows: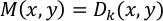
where k represents the frame number at which |Dk (x,y)| = max [|D2 (x,y)|, |D3 (x,y)|, …, |Dn (x,y)|] and n denotes the last frame. The result is saved as “Max_vector.txt”. |M(x, y)| represents the maximum displacement caused by cardiomyocyte contraction at the analysis point (x, y). Contractility value C in arbitrary units is calculated as follows: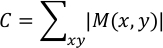
This value is saved at column 7, row 1 in the “Max_vector.txt”. C represents the sum of displacement for all (x, y). The larger the portion where the displacement due to myocardial contraction is large, the larger the value of C. The vector field of maximum displacement, M(x, y), is overlaid with phase contrast image of the first frame (“Phase_contrast.png”) and saved as “Overlaid.png”. As the magnitude of the displacement vector is calculated based on the first frame of the video, it is preferable for the hiPS-CMs to be at resting diastolic period for the first frame.
6. Assessment of cellular damage using flow cytometry
- Dilute a 1 mg/mL solution of propidium iodide to 1:1,000 in PBS.
- Stain the detached cells with propidium iodide.
- Aspirate the medium and place it in an appropriately sized centrifuge tube.
- Centrifuge the tube at 1,000 × g for 5 min, and carefully remove the supernatant so as not to lose sedimented cells.
- Incubate the cells with the diluted propidium iodide solution at room temperature for 15 min in the dark.
- Centrifuge the tube at 1,000 × g for 5 min, and carefully remove the supernatant so as not to lose sedimented cells.
- Reconstitute the cells in ~1 mL of PBS.
NOTE: It is important to collect detached floating cells from the medium to accurately quantify cellular damage.
- Stain attached cells with propidium iodide.
- Wash attached cells twice with PBS gently, then discard PBS.
- Incubate the cells with the diluted propidium iodide solution for 15 min in the dark.
- Aspirate the propidium iodide solution.
- Detach the cells using 0.25% trypsin. Move the cell solution into a centrifuge tube.
- Centrifuge the tube at 1,000 × g for 5 min, and carefully remove the supernatant so as not to lose sedimented cells.
- Reconstitute the cells in ~1 mL of PBS.
- Mix the floating and attached cells for fluorescence activated cell sorting (FACS) analysis.
- Pass the cell solution through a 30 μm filter.
NOTE: Passing the cells to a filter is very important for an accurate FACS measurement. - Analyze the samples using a FACS system.
7. Immunostaining
- Fix the cell sample.
- Aspirate the culture medium.
- Add 4% paraformaldehyde in PBS for 10 min at room temperature.
- Wash the cells three times with PBS.
NOTE: Fresh paraformaldehyde solution is recommended for optimal fixation.
- Permeabilize the cells with 0.2% Triton X-100 for 15 min, then discard the reagent.
- Add 3% bovine serum albumin to block the cells for 30 min.
- Apply primary antibody.
- Discard bovine serum albumin solution from the culture plate.
- Incubate the cells with primary antibodies overnight at 4 °C.
- Remove the antibody solution.
- Wash the cells three times with PBS.
- Apply secondary antibody.
- Discard PBS solution from the culture plate.
- Incubate the cells with secondary antibodies for 30 min at room temperature in the dark.
NOTE: Primary anti-cardiac troponin T (TNNT2) mouse monoclonal antibody is used at a dilution of 1:750 in 3% BSA. The Alexa Fluor 488-conjugated secondary goat anti-mouse antibody is diluted to 1:1,000 in 3% BSA.
- Additional staining of nucleus and actin fibers.
- Remove the antibody solution.
- Incubate the cells in nucleus staining reagent (Table of Materials) and actin staining reagent (Table of Materials) in PBS for 30 min at room temperature in the dark.
NOTE: 4’, 6-diamidino-2-phenylindole (DAPI) or Hoechst 33342 can be used for nuclear staining. - Wash the cells three times with PBS.
- Capture fluorescence images using a fluorescence microscope.
Successfully differentiated cells show spontaneous contraction under the microscope (Video 1). Typically, 50% of wells show spontaneous contraction in <20 days (Supplementary Figure 1). Cardiac marker protein (e.g., cTnT) can be used to confirm successful differentiation (Figure 2).
Typically, cells from the ischemic group show lower viability in MTT assays (Figure 3A) and contractility (Figure 3E,F, Supplementary Figure 2) than those from normoxic control group. Also, the ratio of propidium iodide-positive cells is higher in the ischemic group than in the control group (Figure 3B–D), which indicates higher cellular damage.

Figure 1: Folder structure for displacement vector analysis using imageJ.
“joblist.txt” describes path to movie files each line. If there are three files to analyze, there will be three folders (movie1, movie2, and movie3), in which “analyze.avi” (the movie of interest) should be placed. After the analysis is performed by “vector_analysis.ijm” code, files will be generated (indicated in blue) in each movie folder. Information stored in each file is explained in detail in the main text. Please click here to view a larger version of this figure.
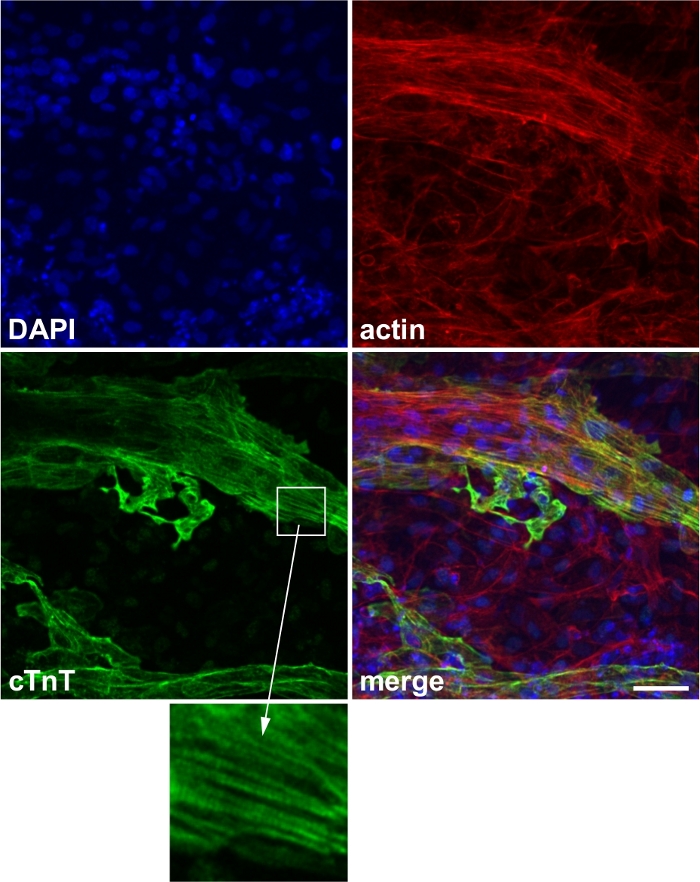
Figure 2: Representative image of cardiac marker staining.
Expression of the cardiac marker protein cardiac troponin T (cTnT). Blue: 4’,6-diamidino-2-phenylindole (DAPI), red: actin, green: cTnT. Inset: striated expression of cTnT, which corresponds to the sarcomere structure. Scale bar, 50 μm. This figure has been modified from Wei et al.13. Please click here to view a larger version of this figure.

Figure 3: Representative images of cellular viability assay, assessment of cellular damage, and contractility.
(A) Comparison of cellular viability (MTT assay). Higher absorbance indicates higher viability. n = 5 for each condition. (B, C, and D) Flow cytometry analysis of propidium iodide (PI)-stained cells. Ischemic cells exhibit a reduced forward scatter intensity and increased PI fluorescence, compared with the control. (D) Percentage of PI-stained cells in the flow cytometry analysis. n = 3 for each condition. (E) Analysis of iPS-CMs contractility using ImageJ software. The red and blue vectors indicate the largest and smallest contractions, respectively. Scale bar, 100 μm. (F) Quantitative analysis of the iPS-CM contractility using code. n = 3 for control and n = 8 for ischemic condition. Error bars represent standard error of the mean. For the statistical analysis, unpaired two-tailed Student’s t-test was performed. *: p < 0.05, **: p < 0.01. ***: p < 0.0001. This figure is cited from Wei et al.13 Please click here to view a larger version of this figure.
Video 1: Differentiated cells show spontaneous contraction under the microscope. Please click here to download this video.
Supplementary Figure 1: Kaplan-Meier analysis of percentage of samples without spontaneous contraction. 50% of iPS-CM samples showed contraction on day 20. On day 30, 64.4% of the samples showed contraction. n = 83. Please click here to download this figure.
Supplementary Figure 2: Assessment of cardiac contractility. Scaled contractility obtained by dividing contractility C by the number of analysis points (x, y) was plotted for control and ischemic groups before and after the exposure to 24 h hypoxia. n = 3 for control and n = 8 for ischemic condition. Please click here to download this figure.
Supplementary Figure 3: Assessment of the content of cardiomyocytes. Immunostaining of cardiac marker protein on a 96 well plate. Green: cTnT, Blue: DAPI. Rate of the differentiated cells was calculated to be 20.7 ± 9.6% by dividing the area of cTnT-stained region by that of DAPI-stained region. Scale bar = 1 mm. n = 4 for control condition. Please click here to download this figure.
Supplementary Coding File. Please click here to download this file.
