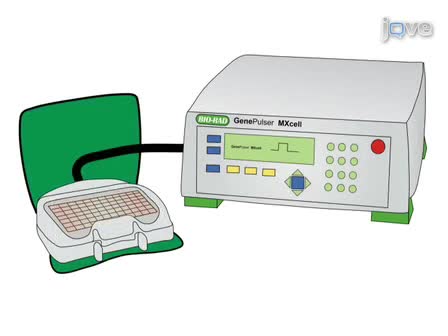
In vitro culture systems have proven indispensible to our understanding of vertebrate myogenesis. However, much remains to be learned about nonmammalian skeletal muscle development and growth, particularly in basal taxa. An efficient and robust protocol for isolating the adult stem cells of this tissue, the myogenic precursor cells (MPCs), and maintaining their self-renewal, proliferation, and differentiation in a primary culture setting allows for the identification of conserved and divergent regulatory mechanisms throughout the vertebrate lineages.