

Summary
Xenopus embryonic ectoderm has become an attractive model for studies of cell polarity. An assay is described, in which subcellular distribution of fluorescent proteins is assessed in ectoderm cells. This protocol will help address questions related to spatial control of signaling.
Abstract
Cell polarity is a fundamental property of eukaryotic cells that is dynamically regulated by both intrinsic and extrinsic factors during embryonic development 1, 2. One of the signaling pathways involved in this regulation is the Wnt pathway, which is used many times during embryogenesis and critical for human disease3, 4, 5. Multiple molecular components of this pathway coordinately regulate signaling in a spatially-restricted manner, but the underlying mechanisms are not fully understood. Xenopus embryonic epithelial cells is an excellent system to study subcellular localization of various signaling proteins. Fluorescent fusion proteins are expressed in Xenopus embryos by RNA microinjection, ectodermal explants are prepared and protein localization is evaluated by epifluorescence. In this experimental protocol we describe how subcellular localization of Diversin, a cytoplasmic protein that has been implicated in signaling and cell polarity determination6, 7 is visualized in Xenopus ectodermal cells to study Wnt signal transduction8. Coexpression of a Wnt ligand or a Frizzled receptor alters the distribution of Diversin fused with red fluorescent protein, RFP, and recruits it to the cell membrane in a polarized fashion 8, 9. This ex vivo protocol should be a useful addition to in vitro studies of cultured mammalian cells, in which spatial control of signaling differs from that of the intact tissue and is much more difficult to analyze.
Protocol
1. In Vitro Fertilization of Xenopus Eggs
- Obtain eggs from female frogs that were injected with human chorionic gonadotropin (400 unit/frog) 12 hours before the experiment.
- Place eggs into a small amount (0.5-1 ml) of 1 x Marc's modified Ringer solution (MMR)10to the eggs and fertilize them in vitro with a small piece of dissected testis. After 2-3 min, add 0.1 x MMR to cover the whole surface of the eggs. In 20 minutes, egg jelly coat is removed by 3 % cysteine - HCL (adjusted to pH 8 with sodium hydroxide). Eggs are washed with 0.1 x MMR three times and left in a cold incubator (13° C) for injections.
- Fertilized eggs are allowed to develop to 2-4 cell stage. For injections, the embryos are transferred into the solution containing 3 % Ficoll, 0.5 x MMR.
2. RNA Microinjection
- RNAs are synthesized from linearized DNA templates using mMessage mMachine kit (Ambion) and diluted with RNase-free water at the stock concentration of 0.1-1 μg/μl. Optimal doses of RNAs for injections are determined in pilot experiments. RNAs for Diversin-RFP, the membrane marker GFP-CAAX and Frizzled 8 are used at 0.1-1 ng per injection.
- Injection needles are prepared with a needle puller from a capillary and then with a needle grinder. Before injection, each needle is calibrated with water to eject 10 nl of liquid per injection.
- For injections, embryos are placed on a plastic dish into a large droplet of 3 % Ficoll, 0.5 x MMR. One microliter of RNA solution is sucked into an injection needle with Narishige microinjector. 10 nl of RNA is injected into animal blastomeres of 8 cell embryos 2-3 times. The injected embryos are transferred into a well of the 12-well plate.
3. Preparing Ectodermal Explants
- When the injected embryos reach early gastrula stage, they are transferred into 0.6 x MMR solution in a 3 cm plastic dish coated with 1 % agarose. Vitelline membrane is removed manually with a pair of forceps. Ectodermal explants are excised from the embryos using a Tungsten needle and a hairloop.
- Ectodermal explants are transferred into a glass vial and fixed with 3.7 % formaldehyde in phosphate buffered saline (PBS) for 30 minutes. Fixed explants are washed with PBS three times (10 minutes each). DAPI is included into the third wash to stain nuclei.
- Explants are mounted on a slide glass. Two strips of scotch tape are attached to the slide and the explants are placed between the two strips. Since the outer surface of explants is pigmented, the inner side of the explants should face the microscope objective. Add two-three 20 μl drops of the mounting solution (70 % glycerol in PBS including 25 mg/ml DABCO, anti-fading reagent)11. Put a coverslip on top.
4. Imaging of Explants Under a Fluorescent Microscope
- Samples are viewed under Zeiss Axioplan fluorescent microscope with appropriate filters.
- Images are taken with the Apotome attachment to visualize a specific plane from several independent explants.
5. Cryosectioning
Cryosectioning is an alternative way to visualize the distribution of fluorescent proteins in the cell and more applicable for immunostaining. At stage 10, embryos are fixed for 1-2 hours with Dent's fixative (20% DMSO, 80% methanol), washed with PBS, and embedded in 15 % fish gelatin/15 % sucrose solution11. The embedded embryos are quickly frozen on dry ice and cryosections are generated on Leica Cryostat. Cross sections would include ectodermal cells that inherit injected RNAs and their translated protein products. The sections retain fluorescence and can be immunostained with specific antibodies and then labeled with secondary antibodies conjugated with fluorescence. Nuclei are stained with DAPI. The mounting media are the same as described above. Imaging can be performed as described above.
6. Representative Results:
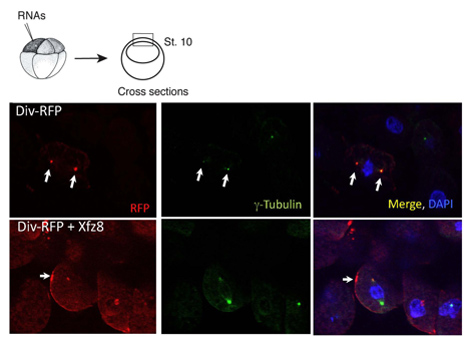
Figure 1. Frizzled receptor recruits Diversin to the cell membrane. Ectoderm cells expressing Frizzled 8 (Xfz8) and Div-RFP RNAs reveal Div-RFP at the cell membrane, instead of the centrosome (as revealed by g-tubulin co-staining). The scheme of the experiment is at the top; a typical cross-section is shown.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
We have used the above protocol to characterize the subcellular localization of Diversin. In animal pole explants, Diversin-RFP was detected next to the nucleus and colocalized with g-tubulin, a centrosomal marker, in cryosections (Figure 1). Once the subcellular localization of a protein is identified, deletion constructs can be generated to establish which protein domains are necessary and sufficient for the subcellular localization. Using this approach, the centrosomal localization domains of Diversin have been mapped to the middle and in the carboxy-terminal domains of the protein, both containing the coiled-coil motif 8.
The same protocol can be used in studies, in which protein localization is altered in response to signaling. We found that Wnt secreted proteins act to relocate Div-RFP to punctate structures adjacent to the cell membrane, whereas Fz8 recruits Div-RFP to cell membrane patches (Figure 1). We further discovered that the carboxy-terminal domain is not essential for membrane recruitment per se, but required for polarized membrane recruitment.
In summary, the above experimental protocol will help in diverse studies of protein-protein interactions and protein localization to different cellular compartments after stimulation of cells with specific growth factors or signaling proteins.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
No conflicts of interest declared.
Acknowledgments
Research in the Sokol laboratory is sponsored by the National Institues of Health.
References
- Gurdon, J. B. Embryonic induction --- molecular aspects. Development. 99, 285-306 (1987).
- Principles of Developmental Genetics. Moody, S. A. , Academic Press. (2007).
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell. 127, 469-480 (2006).
- Klaus, A., Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat Rev Cancer. 8, 387-398 (2008).
- Gordon, M. D., Nusse, R. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem. 281, 22429-22433 (2006).
- Schwarz-Romond, T., Asbrand, C., Bakkers, J., Kuhl, M., Schaeffer, H. J., Huelsken, J., Behrens, J., Hammerschmidt, M., Birchmeier, W. The ankyrin repeat protein Diversin recruits Casein kinase Iepsilon to the beta-catenin degradation complex and acts in both canonical Wnt and Wnt/JNK signaling. Genes Dev. 16, 2073-2084 (2002).
- Moeller, H., Jenny, A., Schaeffer, H. J., Schwarz-Romond, T., Mlodzik, M., Hammerschmidt, M., Birchmeier, W. Diversin regulates heart formation and gastrulation movements in development. Proc Natl Acad Sci U S A. 103, 15900-15905 (2006).
- Itoh, K., Jenny, A., Mlodzik, M., Sokol, S. Y. Centrosomal localization of Diversin and its relevance to Wnt signaling. J. Cell Sci. 122, 3791-3798 (2009).
- Itoh, K., Jacob, J., Sokol, Y. S. A role for Xenopus Frizzled 8 in dorsal development. Mech Dev. 74, 145-157 (1998).
- Newport, J., Kirschner, M. A major developmental transition in early Xenopus embryos: I. characterization and timing of cellular changes at the midblastula stage. Cell. 30, 675-686 (1982).
- Itoh, K., Brott, B. K., Bae, G. U., Ratcliffe, M. J., Sokol, S. Y. Nuclear localization is required for Dishevelled function in Wnt/beta-catenin signaling. J Biol. 4, 3-3 (2005).
