For the simultaneous measurement of pH in both intracellular and endosomal/lysosomal cell compartments, the ratiometric fluorescent pH-sensing probes SNARF-1 and HPTS were used. SNARF-1 is restricted to the cytosolic compartment whereas HPTS allows ratiometric pH measurement of the endosomal/lysosomal compartment. Cell loading with HPTS for 16 hr allows its selective localization in endosomes/lysosomes4. A typical example of fluorescence recorded from HT-1080 cells loaded with the pH-sensing probes is shown in Figure 2A. Evidence that HPTS labels both endosomal and lysosomal compartments was obtained by co-staining with Alexa 546-conjugated transferrin (endosomes) or a fluorescent lysotrophic probe (Lysotracker Deep Reddye). Data clearly showed that HPTS stained both compartments Figure 2A.
Using the methodology described above, intracellular pH values in living cells were determined using a calibration curve for each probe Figures 2B and 2C, based on the ratiometric properties of each pH-sensitive dye. The five calibration solutions were used to convert fluorescence intensities to pH units. Data from three independent calibrations are reported for each pH-sensing probe Figure 2C. Each data point represents a ratio of fluorescence intensity at excitation wavelengths of 458/405 nm for HPTS or emission wavelengths of 644/584 nm for SNARF-1 at a given pH. The calibration curves for both probes showed a non-linear relationship that was best fitted with an exponential equation Figure 2C. The efficiency of the method for detecting cellular compartment-associated pH changes was evaluated in live HT-1080 cells using NH4Cl, a weak base that increases intracellular pH, bafilomycin A1, a pharmacological inhibitor of V-ATPases and EIPA, a NHE-1 inhibitor. NH4Cl triggered intravesicular and cytosolic pH increases from 6.35 to 7.10 and from 7.00 to 7.40, respectively Figures 3A and 3B. Bafilomycin A1 induced alkalinization of the endosomal and lysosomal compartments without altering cytosolic pH. In contrast, EIPA induced acidification of the cytosol reaching a pH value of 6.62 as compared to 7.00 in untreated cells Figures 3A and 3B.
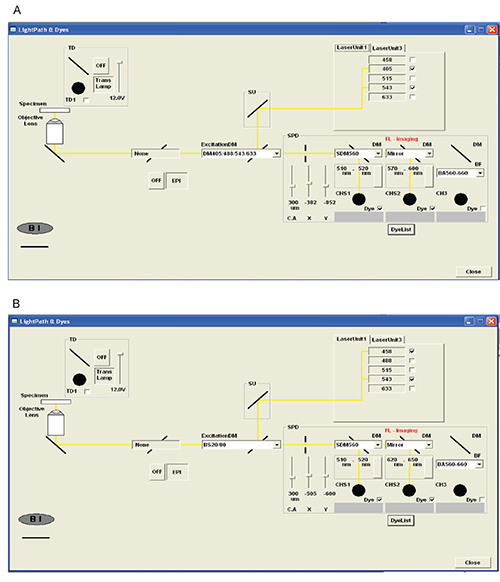
Figure 1. Diagrams of acquisition settings for the confocal laser scanning microscope. (A) The first phase is set to excite HPTS at 405 nm and SNARF-1 at 543 nm simultaneously through a dichroic (DM405/488/543/633) mirror. Fluorescence emission is first recorded for HPTS at 505-525 nm using a dichroic mirror (SDM560). Fluorescence emission of SNARF-1 is then monitored through a bandpass interference filter in the range of 570-600 nm. (B) The second phase is set to simultaneously collect fluorescence emission of HPTS (exc, 458 nm) and SNARF-1 (exc, 543 nm). Probe excitation generated by the laser beam passes through a beam splitter (BS20/80) and fluorescence emission for each probe is collected through the dichroic mirrors used in the first phase. Please click here to view a larger version of this figure.

Figure 2. In situ calibration of pH-sensing probes. (A) Representative images of HPTS and SNARF-1 sub-cellular localization in HT-1080 fibrosarcoma cells. Upper images show that SNARF-1 labels the cytoplasm whereas HPTS-associated fluorescence shows intravesicular localization. HT-1080 cells were incubated for 30 min with Alexa 546-conjugated transferrin (5 μg/ml) or a lysotrophic dye (Lysotracker Deep Reddye) (100 nM), respectively. Lower images show that HPTS colocalizes with transferrin and the lysotrophic dye (60X). (B) Representative images of both pH-sensing probes in cells incubated for 15 min with calibration solutions at pH 6.0 and pH 7.5. (C) Relationship of fluorescence intensity ratios for HPTS and SNARF-1 determined using the calibration solutions as a function of pH. Please click here to view a larger version of this figure.

Figure 3. Modulation of intracellular pH by bafilomycin A1, EIPA and NH4Cl. Pharmacological inhibitors (bafilomycin A1, EIPA) and the intracellular pH modulator NH4Cl were used to validate the methodology. Graphs show pH values recorded in the endosomal/lysosomal compartment (A) or the cytoplasm (B) of HT-1080 cells loaded with HPTS and SNARF-1 and incubated for 30 min in the presence or absence (control) of NH4Cl (20 mM), EIPA (25 μM) or bafilomycin A1 (100 nM). Data are represented as the mean + SEM (** p 0.005, *** p 0.0001).
