

Summary
Hier melden we dubbele etikettering van neurale kuifcellen en bloedvaten met behulp van chickGFP neurale buis intraspecies enting in combinatie met intra-vasculaire DiI injectie. Deze experimentele techniek stelt ons in staat om tegelijkertijd de ontwikkeling van het NCC-afgeleide (enterische) zenuwstelsel en het vasculaire systeem te visualiseren en te bestuderen, tijdens organogenese.
Abstract
Alle zich ontwikkelende organen moeten worden aangesloten op zowel het zenuwstelsel (voor sensorische en motorische controle) als het vasculaire systeem (voor gasuitwisseling, vloeistof- en nutriëntentoevoer). Bijgevolg ontwikkelen zowel het zenuwstelsel als het vasculaire systeem zich naast elkaar en vertonen ze opvallende overeenkomsten in hun vertakkende architectuur. Hier rapporteren we embryonale manipulaties die ons in staat stellen om de gelijktijdige ontwikkeling van neurale kuif-afgeleid zenuwweefsel (in dit geval het enterische zenuwstelsel) en het vasculaire systeem te bestuderen. Dit wordt bereikt door het genereren van kipchimera's via transplantatie van discrete segmenten van de neurale buis en bijbehorende neurale top, gecombineerd met vasculaire DiI-injectie in hetzelfde embryo. Onze methode maakt gebruik van transgene chickGFP embryo's voor intraspecies enting, waardoor de transplantatietechniek krachtiger is dan het klassieke kwartel-kuiken interspecies enting protocol dat sinds de jaren 1970 met groot effect wordt gebruikt. ChickGFP-chick intraspecies enting vergemakkelijkt de beeldvorming van getransplanteerde cellen en hun projecties in intacte weefsels, en elimineert elke mogelijke bias in celontwikkeling die verband houdt met soortverschillen. Deze methode maakt optimaal gebruik van het gemak van toegang van het aviaire embryo (in vergelijking met andere gewervelde embryo's) om de co-ontwikkeling van het darm zenuwstelsel en het vasculaire systeem te bestuderen.
Introduction
Het kippenembryo is een onschatbaar modelorganisme in de gewervelde ontwikkelingsbiologie, niet in de laatste plaats omdat de ontwikkeling ervan in ovo experimentele manipulaties mogelijk maakt die anders onmogelijk te uitvoeren zijn bij gewervelde dieren die zich in de baarmoederontwikkelen . Deze toegankelijkheid en het gemak van manipulaties heeft ertoe geleid dat het kuikenembryo een sleutelrol speelt in vele baanbrekende ontdekkingen op het gebied van ontwikkelingsbiologie. Een van de krachtigste technieken is het gebruik van kwartelkuikenchimerembryo's om het lot van cellen te bestuderen, een methode die in de jaren zeventig door professor Nicole Le Douarin werd geïntroduceerd1-3. In het bijzonder zijn kwartelkuikenchimera's vooral nuttig geweest om populaties van over grote afstanden trekkende neurale kuifcellen (NCC) genetisch te markeren en te volgen tijdens de vroege ontwikkeling. NCC is een multipotente populatie van migrerende cellen, ontstaan in het dorsale ectoderm aan de randen van de neurale buis, die aanleiding geven tot een breed scala aan celtypen in het gewervelde embryo. Deze omvatten craniofaciale structuren (kraakbeen, bot, spieren), neuronen en glia (in het zintuiglijke en autonome zenuwstelsel), melanocyten en een subpopulatie van cellen van het endocriene systeem2,4,5. Een van de belangrijkste factoren die het lot van NCC beïnvloeden, is hun oorspronkelijke locatie langs de voorste-achterste as van de neurale buis. Enterische NCC, die aanleiding geven tot de neuronen en glia van het enterische zenuwstelsel (ENS), ontstaan bijvoorbeeld uit twee afzonderlijke subpopulaties: de eerste in het vagal (caudale hindbrain) gebied, en de tweede in het sacrale gebied van de neurale buis6-13. Inter- of intra-species enting van de overeenkomstige gebieden van de neurale buis zijn de technieken bij uitstek geweest om deze cellen permanent te labelen en vervolgens tracking mogelijk te maken, vanaf hun geboorte aan de randen van de neurale buis, tot hun eindbestemmingen binnen het spijsverteringskanaal6,7,10.
Een andere embryonale manipulatie die gemakkelijker te uitvoeren is bij kuikens, in vergelijking met andere diermodellen, is de vitale etikettering van het vasculaire systeem. Inderdaad, terwijl het kuikenembryo zich ontwikkelt, ligt het bovenop een extra-embryonaal vasculair netwerk dat zuurstof en voedingsstoffen uit de dooier circuleert. Dit toegankelijke vasculaire netwerk, gelegen op het oppervlak van de dooier, kan worden gebruikt als een toegangspoort om het zich ontwikkelende vasculaire systeem van het embryo te labelen tijdens organogenese12,14-17. Intravasculaire injectie van verschillende kleurstoffen, zoals de lipofiele kleurstof DiI, maakt het mogelijk om alle verlichte vaten van het ontluikende vasculaire netwerk af te bakenen / beitsen.
Omdat ontwikkelende organen moeten worden aangesloten op zowel het zenuwstelsel (voor sensorische en motorische controle) als het vasculaire systeem (voor gasuitwisseling, vloeistof- en nutriëntentoevoer), ontwikkelen de twee netwerken zich naast elkaar en delen ze opvallende overeenkomsten in hun vertakkende architectuur18-20. Hier rapporteren we embryonale manipulaties die ons in staat stellen om de gelijktijdige ontwikkeling van het NCC-afgeleide ENS, samen met het vasculaire systeem, tijdens organogenese te bestuderen. Dit wordt bereikt door het genereren van kipchimera's via transplantatie van discrete segmenten van de neurale buis, inclusief de neurale top, gecombineerd met vasculaire DiI-injectie. Als opmars van kwartel-kip chimera's maakt onze methode gebruik van transgene GFP kuikenembryo's voor intraspecies enting, waardoor de transplantatietechniek krachtiger wordt, in termen van beeldvormingscellen en hun projecties, en het elimineren van mogelijke bias die verband houdt met soortenverschillen.
Protocol
1. Voorbereiding van Micro-scalpel voor Neural Tube Ablations
- Vorm een micro-scalpel van een in de handel verkrijgde stalen naainaald.
- Maak eerst de naald aan beide zijden plat met behulp van een slijpschijf die op een aangedreven bankmolen is gemonteerd.
- Begin met het vormen van de scalpel, eerst op een grove Arkansas-steen met behulp van een gecontroleerde cirkelvormige beweging, in afwisselende richtingen, aan beide zijden van de naald.
- Zet dezelfde slijpbewegingen voort op een extra fijne Arkansas-steen om een ultrafijne micro-scalpel te vormen, met een goed gedefinieerde snijkant (figuur 1A, B).
OPMERKING: Alternatieven voor de micro-scalpel kunnen elektrolitair geslepen naalden, in de handel verkrijgbaar wolfraamnaalden of getrokken glazen naalden zijn.
2. Incubeer Wild Type en GFP Eggs naar de gewenste fase
- Bewaar bevruchte kippeneieren en transgene GFP-kippeneieren in een gekoelde incubator bij 14 - 15 °C voorafgaand aan de incubatie, omdat de ontwikkeling bij deze temperatuur wordt gestopt. Bewaar eieren een paar dagen, tot een week.
- Om de ontwikkeling te beginnen, plaatst u wilde type- en GFP-eieren horizontaal op een dienblad en incubeert u tegelijkertijd in een bevochtigde (58 - 60%) incubator bij 37,5 ºC, zodat embryo's zich in overeenkomende stadia bevinden voor neurale buistransplantatie.
- Om embryo's te verkrijgen in het 10 - 12 somietstadium van ontwikkeling voor het uitvoeren van vagal neurale buistransplantatie, incubeer eieren gedurende 1,5 dagen (33 - 38 uur) en stadiumembryo's volgens de ontwikkelingstabellen van Hamburger en Hamilton21.
3. Bereid eieren voor windowing en enten
- Verplaats één ei tegelijk naar een op maat gemaakte eierhouder voor windowing. Maak een klein gaatje in de eierschaal door herhaaldelijk, met een rechte schaar, op het bovenste oppervlak van het puntige uiteinde van het ei te tikken.
- Haal 2 - 3 ml albumine uit het ei met een 181/2 G injectienaald en een spuit van 5 ml. Het verwijderen van de albumine verlaagt de dooier in het ei en vergemakkelijkt de daaropvolgende venstering zonder schade aan het embryo te veroorzaken.
- Gooi de albumine weg. Sluit het gat af met een kleine strook doorzichtige tape die op maat is gesneden met een fijne schaar.
- Tik met een gebogen schaar op een ander gat in het bovenste oppervlak van de eierschaal. Steek de punt van de schaar in het gat en werk, door de schaar evenwijdig aan de bank te houden, in een cirkelvormige beweging om een venster met een diameter van ~ 2 cm bovenop de schaal te snijden.
- Houd de schaar in een stationaire positie en draai het ei. Gooi de verwijderde schijf van de eierschaal weg. Bij E1.5 is het embryo herkenbaar als een donkerdere gele schijf bovenop de dooier.
- Verwijder alle schelpresten die in het ei zijn gevallen met een pincet. Gooi onbevruchte eieren weg (te herkennen aan een kleine witte vlek bovenop de verder lichtgele dooier).
4. Bereid het gastheerembryo voor op geënte weefsels
- Stel de stereomicroscoop in op ooghoogte en optimaliseer de oriëntatie van de zwanenhalslichtbron om het embryo adequaat te verlichten zonder reflecties te veroorzaken.
- Om het embryo goedte visualiseren, injecteer een kleine hoeveelheid Indiase inkt onder het midden van de donkerder gele schijf, met behulp van een mondbuis en een getrokken glazen micropipet(figuur 1C, Oii).
- Bereid de inkt 50:50 met PBS met Penicilline/Streptomycine bij een eindconcentratie van 100 μg/ml. Steek de micropipet door het dooiermembraan buiten de omtrek van het blastoderm en richt de punt voorzichtig direct onder het embryo.
- Lever inkt onder het embryo door op de mondbuis te blazen. Als mondpijpen niet zijn toegestaan, gebruik dan in plaats daarvan een spuit van 1 ml. Zorg ervoor dat u geen luchtbellen onder het embryo introduceert, wat kan leiden tot besmetting, en verwijder vervolgens voorzichtig de glazen micropipet. Dit is een delicate stap die kan leiden tot de dood van het embryo als het niet met precisie wordt gedaan.
- Stage het embryo op verwijzing naar Hamburger en Hamilton21 en noteer het podium in een labboek.
- Maak met behulp van een op maat gemaakte micro-scalpel (of een fijne wolfraamnaald) gemonteerd op een naaldhouder een zeer kleine snee in het vitellinemembraan, naast het gebied waar de micro-operatie zal worden uitgevoerd.
- Breng voorzichtig 2 - 3 druppels PBS aan over de membraanscheur (met behulp van een glazen micropipet en de mondbuis) om ruimte te creëren tussen het embryo en het membraan. Snijd een groter venster in het membraan om het hele gebied bloot te leggen waar de micro-operatie zal plaatsvinden.
- Verwijder het neurale buisgebied van belang met behulp van de micro-scalpel, te beginnen met rostrale en caudale transversale incisies over de hele dorsale neurale buis (ter hoogte van somiet 1 tot 7 in de video).
- Snijd bilateraal tussen de neurale buis en de somieten om de neurale buis te scheiden van de omliggende weefsels, zonder de somieten te beschadigen.
- Scheid heel voorzichtig de neurale buis van de onderliggende notochord, die intact moet blijven. Merk op dat succesvolle neurale buis excisie alle omliggende weefsels perfect intact zal laten (Figuur 2).
- Verwijder de verwijderde neurale buis door deze in een glazen micropipet te aspireren en gooi deze vervolgens weg.
- Noteer het niveau van neurale buis ablatie in een labboek. Het gastheerembryo is nu klaar om de donorneurale buis te ontvangen.
5. Bereid het donortransplantaatweefsel voor
- Selecteer een GFP-embryo met venster en een gematcht GFP door te bekijken onder een fluorescerende stereomicroscoop met FITC-filter. De GFP fluorescentie maakt het heel gemakkelijk om de somieten te visualiseren en het embryo in scène te zet.
- Zodra een stadiumgematcht embryo is geïdentificeerd, verwijdert u het embryo uit het ei door 4 incisies te maken, met pascheff-Wolff veerschaar (figuur 1C, l) in een rechthoekvorm rond het embryo en pak het vervolgens voorzichtig op met een embryolepel.
- Plaats het embryo in een vierkant horlogeglas met een sylgard polymeerbasis. Schud het embryo voorzichtig met Dumont #5 pincet om eventuele aangehechte dooier te verwijderen. Verwijder het vitellinemembraan en speld het embryo op de polymeerbasis met behulp van roestvrijstalen minutienpennen(afbeelding 1C).
- Maak met behulp van de lenteschaar 4 incisies in een rechthoekige vorm rond de neurale buis en de omliggende somieten, in dezelfde regio die uit het gastheerembryo is verwijderd.
- Breng met behulp van een plastic transferpipet de neurale buis en somietweefsels van het donor GFP-embryo over in een horlogeglas met 0,2% pancreas in Pen/Strep PBS.
- Laat de enzymatische spijsvertering 10 minuten doorgaan bij RT om de weefsels te helpen scheiden. Gebruik na incubatie in enzym roestvrijstalen minutienpennen die op een handvat zijn gemonteerd om de neurale buis handmatig te scheiden van alle aangrenzende weefsels.
- Breng met behulp van een glazen micropipet de gedissocieerde neurale buis over naar een ander horlogeglas met DMEM + 10% serum(bijv.geiten-, paarden- of foetale kalf) op ijs, om de overtollige pancreas te spoelen en de enzymatische spijsvertering te stoppen. Na 5 minuten is de ontlede neurale buis klaar om orthotopisch in de kuikengastheer te worden geënt (figuur 2 en S1).
6. Transplantaat het weefsel
- Breng met behulp van een glazen micropipet de ontlede neurale buis voorzichtig over van het horlogeglas naar het gastheerembryo. Plaats de neurale buis in de juiste voorste-posterieure oriëntatie en duw de explant voorzichtig naast het verwijderde gebied van de kuikengastheer met behulp van de micro-scalpel. Laat een klein stukje ectoderm bevestigd aan, of door een kleine inkeping in het dorsale oppervlak te snijden om de oriëntatie van de neurale buis te identificeren.
- Gebruik indien nodig de micro-scalpel om de explant te trimmen tot de exacte grootte van het accijnsgebied.
- Leid de neurale buis voorzichtig naar het ablated-gebied en plaats deze zo dat de dorsale kant correct is georiënteerd. Gebruik een glazen micropipet, gemonteerd op een mondbuis, om PBS en/of vloeistof rondom de ent te verwijderen. Dit helpt de donor- en gastheerweefsels zich te hechten en de ent te vestigen.
- Sluit het hele raam af met 24 mm brede doorzichtige tape om uitdroging en vervuiling te voorkomen.
- Label het chimerische embryo door te markeren met een potlood op de eierschaal en noteer het nummer in het labboek. Breng het ei terug naar de incubator voor verdere ontwikkeling.
7. Injecteer DiI in bloedvaten van gastheerembryo
- Haal op het gewenste experimentele tijdstip (hier, 3 - 10 dagen later) het chimerische embryo uit de incubator en verwijder de doorzichtige tape met een rechte schaar om toegang te krijgen tot het embryo in het ei.
- Vergroot indien nodig het venster in de schaal met behulp van de schaar. Zorg ervoor dat u het chorioallantoïsche membraan niet beschadigt als het aan de schaal is bevestigd, wat zou leiden tot een bloeding en de etikettering van het bloedvat in gevaar zou brengen.
- Kies een toegankelijke ader op de dooier en zorg ervoor dat de bloedstroom naar het embryo is gericht. Kies een vertakkingspunt van een van de vitellineaders (figuur 3B, C).
OPMERKING: Bij E6.5 - E7.5 moet het chorioallantoïsche membraan mogelijk voorzichtig opzij worden geschoven met een pincet om toegang te krijgen tot de dooieraders. Na E8.5 is de enige optie om in een van de chorioallantoïsche membraanaders te injecteren, omdat het chorioallantoïsche membraan in dit stadium het embryo volledig bedekt. - Verwijder het vitellinemembraan boven het gekozen injectiepunt met twee Dumont #5 pincet door in tegengestelde richting te scheuren.
- Breek een getrokken glazen naald met behulp van een Dumont #5 en pas de diameter aan de geschatte grootte van de ader aan voordat u deze laadt met CellTracker CM-DiI. Maak de DiI-voorraadoplossing bij 40 μg/μl in DMSO en bewaar bij -20 °C. Bereid de werkoplossing in 0,3 M sucrose/PBS bij een concentratie van 4 μg/μl.
- Aspireer tussen 5 - 10 μl DiI in 0,3 M sucrose/PBS in de naald met behulp van zuiging met een mondbuis. Oudere embryo's hebben mogelijk tot 25 μl of meer nodig. Vanaf E8.5 hebben embryo's grotere, gespierdere aderen, die mogelijk op hun plaats moeten worden gehouden met een Dumont-#5 voordat ze worden gestoken met de dii-geladen glazen naald.
- Steek de naald snel in de ader en blaas gestaag met de mondbuis om de DiI in staat te stellen langzaam aan te sluiten bij de bloedstroom zonder een stolsel te vormen. U kunt ook een drukinjector gebruiken voor dii-levering.
8. Embryo's oogsten voor sectie- of volkorenonderzoek
- Om zoveel mogelijk DiI in het embryo te behouden, oogst u het embryo onmiddellijk na de injectie door het op een geperforeerde lepel te scheppen en de bloedvaten en bindweefsels met een rechte schaar te snijden, om het embryo uit de dooier te bevrijden.
- Verwijder eventuele losse membranen en ontleed de organen van belang(d.w.z.de longen en het spijsverteringskanaal in deze tutorial), waarbij u er goed op moet letten het weefsel niet te comprimeren, wat diffusie van de DiI creëert. Repareer de weefsels onmiddellijk door onderdompeling in 4% PFA gedurende 1 - 2 uur bij RT.
- Spoel het weefsel gedurende 5 minuten in PBS en vervolgens 15 minuten in PBS met 5 μg/ml DAPI. Monteer de monsters op een overbrugde microscoopschuif voor het hele montageonderzoek of sluit ze in voor cryo-sectioning.
Representative Results
Figuur 1 toont typische instrumenten die nodig zijn om microchirurgische isolatie en transplantatie van de neurale buis uit te voeren. Figuur 2 toont de transplantatieprocedure. Na transplantatie worden embryo's gescreend op transplantatiesucces. Dit omvat het onderzoeken van het embryo onder een stereofluorescentiemicroscoop, meestal de ochtend na microchirurgie, op de aanwezigheid van ent-afgeleide (GFP+) NCC. Als transplantatie een succes is geweest, kan GFP + NCC worden waargenomen in de buurt van de neurale buis en in vroege migratieroutes die naar de foregut leiden. Als de procedure niet succesvol is geweest, zal GFP+ NCC niet buiten de neurale buis worden waargenomen, of als ze aanwezig zijn in de gastheer kunnen ze in kleinere aantallen zijn. Deze mislukte embryo's worden weggegooid. Meestal worden 5-8 neurale buistransplantaties op één dag uitgevoerd en van deze 80% zijn succesvol. Redenen voor mislukte neurale buistransplantatie zijn de dood van het embryo als gevolg van weefselschade opgelopen tijdens microchirurgie, of het falen van de neurale buis om te integreren in het gastheerembryo. Dit laatste kan het gevolg zijn van een slechte plaatsing van de neurale buis in de gastheer of van een neurale buis van slechte kwaliteit als gevolg van een slechte dissectietechniek of van overmatige blootstelling aan dissociatie-enzym. De eerste screeningstap, evenals soortgelijke latere onderzoeken voor GFP+ cellen, is nuttig omdat het betekent dat tijd en middelen niet worden verspild door experimenten uit te voeren op embryo's die geen NCC met GFP-label in de darm hebben.
Figuur 3 toont de procedure voor diI-injectie van de bloedvaten. De efficiëntie/het succes van de DiI-injectietechniek hangt af van: ten eerste, het snijden van de injectienaald tot de optimale diameter voor de beoogde ader, ten tweede een nauwkeurig gebaar bij het inbrengen van de naald in de ader (dus niet door de andere kant prikken), en ten derde het vermijden van het aansluiten van de naald tijdens de injectie door met een constante snelheid te blazen. Als een van deze drie parameters onjuist wordt uitgevoerd, zal het embryo bloeden of enkele uren nodig hebben om te herstellen voordat een tweede poging wordt gedaan, omdat de bloeding het bijna onmogelijk maakt om onmiddellijk opnieuw te injecteren. Succesvolle embryo's moeten onmiddellijk worden geselecteerd door te bekijken onder een stereofluorescentiemicroscoop en moeten snel worden ontleed. In succesvolle embryo's zijn dii-gelabelde bloedvaten in het hele embryo aanwezig (figuur 3C,D) inclusief capillaire bedden (figuur 3D).
Bij het oogsten van embryo's en onderzoek van weefselsecties of hele maagdarmkanaalorganen, typische resultaten onthullen GFP + NCC binnen het primitieve ENS en de fijne structuur van de DiI-gelabelde darmbloedvatnetwerken (Figuur 4) Wholemount-preparaten kunnen worden onderzocht met confocale microscopie waarbij beeldstapels driedimensionale (3D) reconstructies produceren die de onderlinge relaties tussen de fijne projecties van GFP + ENS-cellen en het DiI-bevlekte vasculaire systeem aantonen (Figuur 4 ; G-I; Video's 1 en 2).

Figuur 1. Aanbevolen microchirurgie-instrumenten. A) micro-scalpel in de vorm van een naainaald. (B) fijne Arkansas steen voor het vormen van een micro-scalpel. c) a) rechte schaar, b) gebogen schaar, c) 5 ml spuit met 181/2 G injectienaald, d) plastic pipet, e) op maat gemaakte eierhouder, f) zwarte inkt, g) vierkant horlogeglas, h) vierkant horlogeglas met zwarte sylgardbasis, i) micro-scalpel op naaldhouder, j) minutienpennen, k) minutien of wolfraamnaald op naaldhouder, l) Pascheff-Wolff veerschaar, m) Dumont #5 pincet, n) geperforeerde lepel Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 2. Intraspecies neurale buistransplantatie. Chick embryo / GFP neurale buis beelden zijn gewijzigd van Delalande et al. Vascularisatieis niet nodig voor darmkolonisatie door enterische neurale kuifcellen. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 3. Intraveneuze diI injectie. (A) Aanbevolen instrumenten: a) CellTracker CM-DiI drop op parafilm, b) getrokken glazen injectienaald, c) mondbuis. (B) Schematisch schema van intraveneuze DiI-injectie in E4 chimerisch kuikenembryo. C) in ovo DiI intraveneuze injectie met fijne glazen naald met DiI ingebracht in ader (pijl). (D) E4 chimerisch embryo na DiI-injectie (rood) met GFP+ neurale buis (pijl). (E) DiI gekleurd fijn bloedvatnetwerk in een levend embryo, 24 uur na injectie. Br: hersenen; H: hart; LB: ledemaat knop; A: Allantois. Afbeeldingen in (C) en (D) zijn gewijzigd van Delalande et al. 12 Vascularisatie is niet nodig voor darmkolonisatie door enterische neurale kuifcellen. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 4: Representatieve resultaten in de maag en caecum van een E5.5 kuikenembryo. (A-C) 3 dimensionale (3D) reconstructie van een confocale beeldstapel in het gebied van de maag met (D) de GFP+ enterische neurale kuifcellen (ENCC) (E) het DiI-bevlekte vasculaire systeem en (F) een samengevoegd beeld van beide netwerken D-F Histologische secties op het niveau van de maag met (G) het GFP+ ENCC (H) het DiI-bevlekte vasculaire systeem en (I) een samengevoegd beeld van beide netwerken. Kernen zijn gekleurd met DAPI (cyaan). (G-H) 3D-reconstructie van een confocale beeldstapel in het caecumgebied met (A) het GFP+ ENCC-migratiefront in het groen, (B) het dii-bevlekte vasculaire systeem in rood en (C) een samengevoegd beeld van beide netwerken. Afbeeldingen (A-F) zijn gewijzigd van Delalande et al. 12 Vascularisatie is niet nodig voor darmkolonisatie door enterische neurale kuifcellen. Klik hier om een grotere versie van deze afbeelding te bekijken.

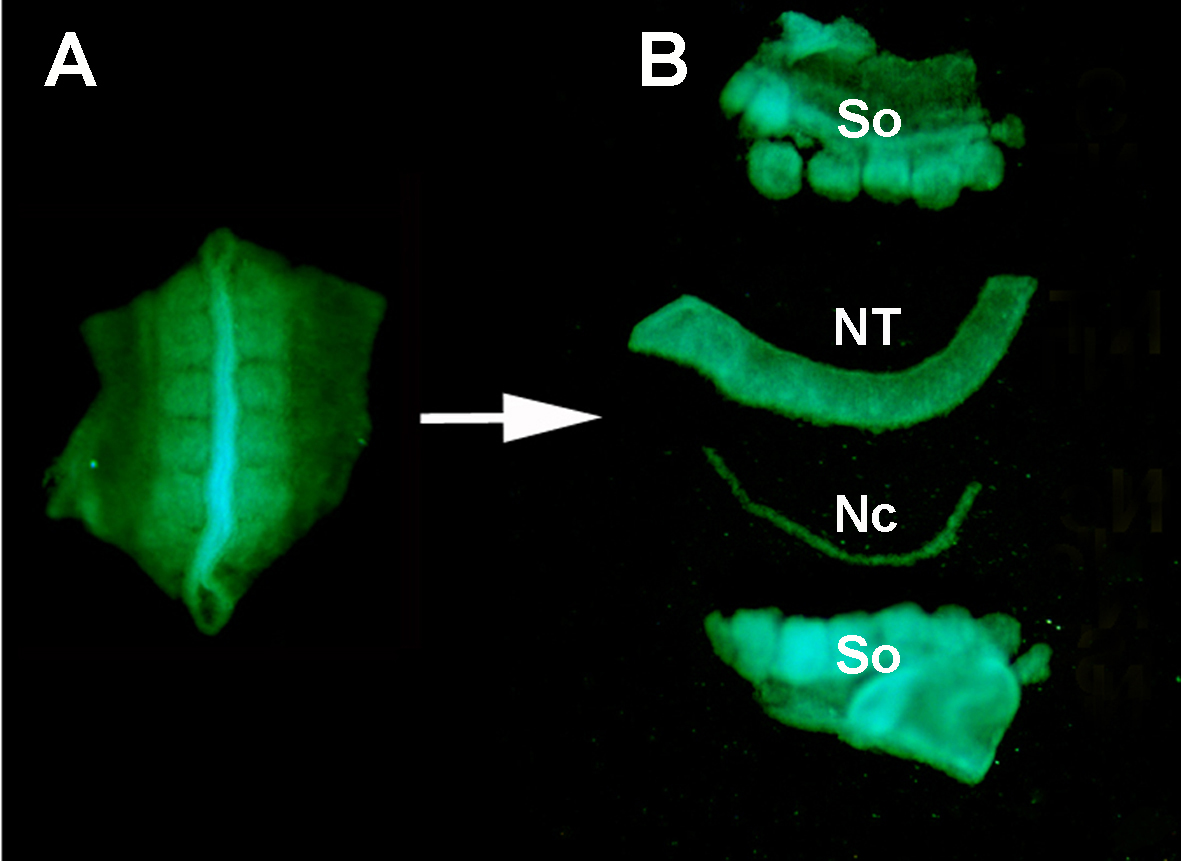
Figuur S1. Isolatie van een donor GFP+ neurale buis uit de omliggende weefsels door enzymatische spijsvertering en microdissectie. (A) GFP+ neurale buis en aangrenzende somieten ontleed uit het donorembryo. (B) Geïsoleerde neurale buis na pancreasvertering en micro-dissectie met behulp van roestvrije minutienpennen. Dus: somieten; NT: neurale buis; Nc: Notochord.

Video 1. 3-dimensionale 360° rotatie van het beeld in figuur 4C, met het vasculaire systeem en de ENCC in de maag bij E5.5 (HH27-28). Klik hier om deze video te bekijken.

Video 2. 3-dimensionale 360° rotatie van het beeld in figuur 4I, met het vasculaire systeem en het ENCC-migratiefront in het gebied van het caecum bij E5.5 (HH27-28). Klik hier om deze video te bekijken.
Discussion
De methode van intraspecies neurale buistransplantatie, gecombineerd met hier beschreven bloedvatetikettering, maakt optimaal gebruik van het gemak van toegang van het aviaire embryo in het ei (in vergelijking met andere gewervelde embryo's) om de co-ontwikkeling van een element van het autonome zenuwstelsel (het ENS) en het vasculaire systeem te bestuderen.
Voor het labelen van NCC-derivaten heeft de chickGFP-chick intraspecies entingsmethode die we beschrijven een aantal voordelen ten opzichte van de klassieke kwartel-kuikenchimera-methode die meer dan 40 jaar geleden1-3werd vastgesteld. Ten eerste is GFP-fluorescentie onder FITC-licht extreem helder, in die mate dat GFP + -cellen gemakkelijk te onderscheiden zijn in levende chimerische embryo's. Hierdoor kan het succes van de ent in ovoworden gecontroleerd , terwijl kwartelkuikentransplantatie vereist dat het embryo wordt gedood, verwerkt en geïmmuntaineerd met QCPN, voordat het succes van de ent kan worden vastgesteld2. Ten tweede is GFP-expressie in het transgene kuikenGFP cytoplasmatisch, daarom labelt het niet alleen cellichamen, maar maakt het ook mogelijk om de projecties van de getransplanteerde cellen te visualiseren22. Hierdoor kunnen ingewikkelde neuronale netwerken met hoge resolutie worden waargenomen (merk op dat fijne projecties het beste kunnen worden gevisualiseerd wanneer het monster wordt geïmmuniseerde met anti-GFP-antilichamen). Aangezien QCPN-etikettering beperkt is tot de kwartelcelkern, worden dergelijke netwerken niet onthuld met behulp van kwartelkuikenchimera's. Ten derde elimineert intraspecies enting mogelijke soortverschillen tussen cellen in het chimerische embryo. Aangezien kwartelpbryo's een kortere incubatietijd hebben dan kuikencellen (19 dagen versus 21 dagen), is gesuggereerd dat kwartelcellen een hogere proliferatiesnelheid hebben dan kuikencellen, wat mogelijk de ontwikkeling van de chimerische weefsels kan beïnvloeden23. Interessant is ook dat bij planten is aangetoond dat interspecies enting uitgebreide veranderingen in DNA-methylatiepatronen in de gastheer 24kan veroorzaken. Ten vierde faciliteert chickGFP rugtransplantatie-experimenten om onderwerpen als het lot van de NCC en celverbintenis aan te pakken25. Ten vijfde is het transgene kuikenGFP ook nuttig voor vele andere technieken, waaronder FACS-sortering van GFP+ celsubpopulaties, organotypische cultuur van organen die GFP + cellen bevatten, genetische manipulatie van GFP + geënt weefsel via elektroporatie van expressieplasmiden26, en andere beeldvormingstechnologieën zoals optische projectietomografie27.
De neurale buistransplantatie benadering kan worden gewijzigd door microchirurgische vervanging van kortere hoeveelheden neurale buis. Door het gebruik van kleinere segmenten van neurale buis is de microchirurgie potentieel minder schadelijk voor het embryo en kan de overleving worden verbeterd. Het nadeel van het transplanteren van minder neurale buis is echter dat het aantal GFP + NCC in de gastheer zal worden verminderd. Gebruikers konden proberen een evenwicht te vinden tussen de hoeveelheid getransplanteerde neurale buis om optimale overleving van embryo's te geven, en de aantallen GFP + NCC in de gastheerdarm voldoende om informatieve resultaten te geven.
Voor scheepsschilderen heeft DiI het voordeel dat de fluorescentie zeer helder en robuust is. Het heeft ook de capaciteit om te diffuus te maken tijdens fixatie, waardoor vlekken van de fijnste geopende haarvaten worden verzekerd. Omdat het een vitale kleurstof is, kunnen embryo's de injectieprocedure overleven en zich blijven ontwikkelen met een gekleurd vasculair systeem (tot 24 uur in onze handen, hoewel de kleuring na verloop van tijd punctueeler wordt, zie figuur 3E). De combinatie van chickGFP enting met DiI vasculaire schilderkunst is daarom compatibel met live beeldvorming. Naast al deze voordelen is het belangrijk op te merken dat vasculaire injectie alleen geluminiseerde vaten labelt en daarom geen ongeopende haarvaten, endotheelpuntcellen of geïsoleerde endotheelcellen identificeert. Verdere vooruitgang op het gebied van aviaire transgenese zou echter nieuwe manieren kunnen bieden om dergelijke problemen te omzeilen, zoals blijkt uit experimenten met kwartelembryo's met Tg(tie1:H2B-eYFP) om vasculaire morfogenese te bestuderen28. Een andere beperking van deze techniek is dat voor effectieve etikettering van bloedvaten in embryo's bij E7.5 en daarna grotere hoeveelheden kleurstof moeten worden geïnjecteerd, wat experimenten duur kan maken. Een wijziging van de techniek kan echter goedkope bloedvatetiketteringsinkt14omvatten, hoewel deze aanpak niet in onze handen is geprobeerd.
Kritieke stappen van de procedures omvatten het proces van het visualiseren van het embryo door inkt onder de blastodisc te injecteren. Als het membraan dat de dooier bedekt in dit stadium door de met inkt gevulde naald wordt gescheurd, wordt de overleving van embryo's ernstig aangetast. Ook is het belangrijk, bij het voorbereiden van een donorneurale buis, dat het weefsel niet te lang in de pancreas achterbleef (beschouw ongeveer 10 minuten als een maximum). Langdurige blootstelling aan pancreas beschadigt het weefsel en de neurale buis is dan moeilijk te hanteren en het zal niet goed in de gastheer worden opgenomen. Het opdoen van ervaring met de DiI-injectietechniek op embryo's van het wilde type is essentieel voordat chimerische embryo's worden geïnjecteerd, omdat slechts één poging tot injecteren over het algemeen mogelijk is voor elk embryo. DiI-volume en naalddiameter zijn kritieke parameters voor elk embryo en moeten worden beoordeeld op wilde, gefaseerde controles.
Kortom, onze dual labeling methode van neurale buistransplantatie en DiI vessel painting in levende kuikenembryo's kan worden gebruikt om de interrelaties tussen NCC en bloedvatnetwerken tijdens organogenese te onderzoeken. Gezien de mechanismen die verantwoordelijk zijn voor het vaststellen van de juiste doelinvatie en vascularisatie tijdens orgaanontwikkeling nog grotendeels onbekend zijn, heeft deze methodologie potentieel voor toekomstige ontdekkingen op dit gebied.
Disclosures
De auteurs verklaren dat ze geen concurrerende financiële belangen hebben.
Acknowledgments
Bevruchte GFP kippeneieren werden geleverd door Prof. Helen Sang, The Roslin Institute en University of Edinburgh, UK. De Roslin Transgenic Chicken Facility wordt gefinancierd door de Wellcome Trust en door de Biotechnology and Biological Sciences Research Council (BBSRC). Het werk werd gedeeltelijk gefinancierd, en NT gesteund, door Great Ormond Street Hospital Children's Charity, Londen, UK. De auteurs danken Ben Jevans, UCL Institute of Child Health, voor hulp bij het voorbereiden van embryo's op enten.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}