

Summary
여기에서 우리는 혈관 내 DiI 주입과 결합된 병아리GFP 신경관 내종 이식을 사용하여 신경 문장 세포 및 혈관의 이중 라벨을 보고합니다. 이 실험 기술은 우리가 동시에 시각화하고 유기 발생 동안 NCC 파생 (장내) 신경계와 혈관 시스템의 개발을 연구 할 수 있습니다.
Abstract
모든 개발 기관은 신경계 (감각 및 운동 제어용)뿐만 아니라 혈관 시스템 (가스 교환, 유체 및 영양 공급)에 연결되어야합니다. 따라서 신경 과 혈관 시스템은 서로 함께 개발하고 분기 아키텍처에서 눈에 띄는 유사성을 공유합니다. 여기서 우리는 우리가 신경 문장 유래 신경 조직의 동시 발달을 연구할 수 있는 배아 조작을 보고합니다 (이 경우 장 신경계), 및 혈관 시스템. 이것은 신경관의 이산 세그먼트의 이식을 통해 닭 키메라를 생성하고, 관련 신경 문장, 같은 태아에 있는 혈관 DiI 주입과 결합하여 달성됩니다. 우리의 방법은 종 내 이식을 위한 형질 전환병아리 GFP 배아를 사용하여, 이식 기술은 1970 년대 부터 큰 효과로 사용되는 고전적인 메추라기 병아리 종 이식 프로토콜보다 더 강력한 만들기. ChickGFP-병아리종 이식은 이식된 세포의 화상 진찰을 용이하게 하고 그대로 조직에 있는 그들의 돌출을, 종 다름에 연결된 세포 발달에 있는 어떤 잠재적인 편견든지 제거합니다. 이 방법은 조류 배아의 접근의 용이성을 최대한 활용하여 (다른 척추 동물 배아와 비교하여) 장 신경계 및 혈관 시스템의 공동 개발을 연구합니다.
Introduction
닭 배아는 척추 동물 발달 생물학에서 귀중한 모형 유기체입니다, 적어도 오보에 있는 그것의 발달은 자궁에서발전하는 척골가에서 발전하는 척골가에서 능력을 발휘하는 것을 허용하는 실험적인 조작을 허용하기 때문에. 이러한 접근성과 조작의 용이성은 발달 생물학 분야에서 많은 정액 발견에서 중요한 역할을 하는 병아리 배아로 이어졌습니다. 가장 강력한 기술 중 세포 운명을 연구하기 위해 메추라기 병아리 키메라 배아의 사용이었다, 교수 니콜 르 두아린에 의해 개척 방법 1970의1-3. 특히, 메추라 병아리 키메라는 초기 개발 중에 유전적으로 유전적으로 유방 신경 문장 세포 (NCC) 인구를 따르는 데 유용했습니다. NCC는 척골가 태아를 통해 세포 모형의 넓은 범위를 초래하는 신경관의 여백에 등등 외항에서 생기는 철새 세포의 다능한 인구입니다. 여기에는 두개안면 구조(연골, 뼈, 근육), 신경 및 신경교(감각 및 자율 신경계), 멜라닌세포 및 내분비 계의 세포의 하위집단이 포함된다 2,4,5. NCC 운명에 영향을 미치는 가장 중요한 요인 중 하나는 신경 관의 전방 후방 축을 따라 그들의 초기 위치. 예를 들어, 장신경계(ENS)의 뉴런과 신경교를 야기하는 장내 NCC는 두 개의 이산 하위 집단에서 발생합니다: 첫 번째는 vagal (caudal hindbrain) 부위에 위치하고, 신경관6-13의천년 영역에서 두 번째이다. 신경관의 해당 영역의 종 내 이식은 이러한 세포를 영구적으로 라벨링하고 이후에 신경관의 여백에 출생에서 부터 소화관 내의 최종 목적지까지 추적을 허용하는 데 선택하는 기술이었다6,7,10.
다른 동물 모델에 비해 병아리에서 수행하기 쉬운 또 다른 배아 조작은 혈관 시스템의 중요한 라벨링입니다. 실제로 병아리 배아가 발달함에 따라 노른자에서 산소와 영양분을 순환시키는 배아 혈관 네트워크 위에 놓입니다. 노른자 표면에 위치한 이 접근 가능한 혈관 네트워크는 유기발생 시 배아의 발달 혈관 시스템을 라벨로 표시하는 게이트웨이로 사용될 수 있다12,14-17. 리포필성 염료 DiI와 같은 다양한 염료의 내트라바산 분사는 초기 혈관 네트워크의 모든 발광 된 혈관을 묘사 / 얼룩시킬 수 있게합니다.
개발 기관은 신경계 (감각 및 운동 제어를 위한) 뿐만 아니라 혈관 시스템 (가스 교환, 유체 및 영양 공급)에 연결되어야하기 때문에, 두 네트워크는 서로 함께 개발하고 분기 아키텍처에 눈에 띄는 유사성을 공유18-20. 여기서 우리는 우리가 유기발생 도중 혈관 시스템과 더불어 NCC 파생 된 ENS의 동시 발달을 연구할 수 있는 배아 조작을 보고합니다. 이것은 신경 관을 포함하여 신경관의 이산 세그먼트의 이식을 통해 닭 키메라를 생성하여 달성됩니다, 혈관 DiI 주입과 결합. 메추라기 닭 키메라의 진보로, 우리의 방법은 종 이식을 위한 형질 전환 GFP 병아리 배아를 사용하여, 화상 진찰 세포 및 그들의 투영의 관점에서 이식 기술을 더 강력하게 만들고, 종 다름에 연결된 잠재적인 편견을 제거합니다.
Protocol
1. 신경관 절제용 마이크로 메스 준비
- 시판되는 강철 재봉 바늘에서 마이크로 메스를 형성한다.
- 먼저 전원 벤치 그라인더에 장착 된 연삭 휠을 사용하여 양쪽의 바늘을 평평하게합니다.
- 메스를 형성하기 시작, 먼저 거친 학년 아칸소 돌에 제어 된 원형 운동을 사용하여, 대체 방향으로, 바늘의 양쪽에.
- 매우 미세한 미세한 미세 한 미세 메스를 형성하기 위해 여분의 미세 한 학년 아칸소 돌에 동일한 선명 한 움직임을 수행, 잘 정의 된 절삭 날(그림 1A, B).
참고: 마이크로 메스에 대한 대안은 전정적으로 날카로운 바늘, 시판되는 텅스텐 바늘 또는 당겨진 유리 바늘일 수 있습니다.
2. 원하는 단계에 야생 유형과 GFP 계란을 인큐베이션
- 이 온도에서 개발이 중단됨에 따라 인큐베이션 전에 14-15°C의 냉각된 인큐베이터에 수정된 닭고기 계란과 트랜스제닉 GFP 치킨 알을 보관합니다. 계란을 며칠 동안 보관하십시오.
- 개발을 시작하려면 야생 유형과 GFP 알을 수평으로 트레이에 놓고 가습 (58 - 60 %) 동시에 배양합니다. 37.5 ºC에서 배아가 신경관 이식을 위한 일치하는 단계에 있도록 인큐베이터.
- 10-12소미트 단계에서 혈관 신경관 이식을 수행하기 위한 개발 단계, 1.5일 동안 계란을 배양(33-38시간) 및 함부르크와해밀턴(21)의개발 테이블에 따라 단계 배아를 배양한다.
3. 창과 접목을 위한 계란 준비
- 한 번에 한 개의 달걀 을 주문 제작 한 계란 홀더로 이동하여 창을 지정합니다. 계란의 뾰족한 끝의 상부 표면에 직선 가위를 가진 반복적으로 두드려 계란 껍질에 작은 구멍을 만듭니다.
- 181/2 G 피하 바늘과 5ml 주사기로 달걀에서 알부민 2-3 ml를 제거하십시오. 알부민을 제거하면 알란 내의 노른자를 낮추고 배아에 손상을 주지 않으면서 후속 창을 용이하게 합니다.
- 알부민을 폐기합니다. 미세 한 가위로 크기로 잘라 투명 테이프의 작은 스트립으로 구멍을 밀봉.
- 곡선 가위를 사용하여 달걀 껍질의 상부 표면에 있는 다른 구멍을 누릅니다. 가위의 끝을 구멍에 삽입하고 가위를 벤치와 평행하게 유지하여 원단 동작으로 작동하여 쉘 위에 ~2cm 직경의 창을 자른다.
- 가위를 고정 된 위치에 유지하고 계란을 회전시십시오. 제거된 계란 껍질 디스크를 폐기합니다. E1.5에서 배아는 노른자 위에 어두운 노란색 디스크로 인식할 수 있습니다.
- 핀셋을 사용하여 계란 내에 떨어진 껍질 파편을 제거합니다. 수정되지 않은 계란을 버리십시오 (그렇지 않으면 밝은 노란색 노른자 위에 작은 흰색 반점으로 식별됨).
4. 접목 조직을 받기 위해 호스트 배아를 준비합니다.
- 스테레오 현미경을 눈 높이로 조정하고 구스넥 광원의 방향을 최적화하여 반사를 일으키지 않고 배아를 적절하게 비춥춥시킵니다.
- 적절한배아를 시각화하려면, 입 튜브와 당겨진 유리 마이크로 파이펫(도 1C, Oii)를사용하여 어두운 노란색 디스크의 중앙 아래에 소량의 인도 잉크를 주입하십시오.
- 100 μg/ml 최종 농도에서 페니실린/스트렙토마이신을 함유한 PBS로 잉크 50:50을 준비한다. blastoderm의 둘레 바깥에 노른자 멤브레인을 통해 마이크로 파이펫을 삽입한 다음 배아 바로 아래에 팁을 조심스럽게 각도로 기울입니다.
- 입 튜브에 불어 배아 아래에 잉크를 전달합니다. 구강 파이프팅이 허용되지 않는 경우 대신 1ml 주사기를 사용하십시오. 오염으로 이어질 수있는 배아 아래에 기포를 도입하지 않도록주의한 다음 유리 마이크로 파이펫을 조심스럽게 제거하십시오. 이것은 정밀하게 행하지 않을 경우 배아의 죽음으로 이어질 수있는 섬세한 단계입니다.
- 햄버거와 해밀턴(21)을 참조하여 배아를 무대와 실험실 책에서 무대를 기록합니다.
- 바늘 홀더에 장착 된 사용자 정의 만든 마이크로 메스 (또는 미세 텅스텐 바늘)를 사용하여, 마이크로 수술이 수행 될 영역 옆에, vitelline 막에 매우 작은 가스를 합니다.
- 조심스럽게 배아와 멤브레인 사이의 공간을 만들기 위해 멤브레인 눈물 (유리 마이크로 파이펫과 입 튜브를 사용하여) 막 눈물 위에 PBS 의 2 - 3 방울을 적용합니다. 멤브레인의 더 큰 창을 잘라 미세 수술이 일어날 전체 영역을 노출시킵니다.
- 전체 등쪽 신경관에서 로스트랄 및 caudal 횡단 절개 (비디오에서 소미트 1 에서 7의 수준에서)에서 시작하여 마이크로 메스를 사용하여 관심있는 신경 관 영역을 제거하십시오.
- 신경관과 소미트 사이를 양측으로 잘라 주변 조직으로부터 신경관을 분리하소미를 손상시키지 않고 잘라냅니다.
- 매우 부드럽게 그대로 유지해야 기본 notochord에서 신경 관을 분리합니다. 성공적인 신경관 절개는 모든 주변 조직을 완벽하게 그대로 둡니다(그림2).
- 절제된 신경관을 유리 마이크로 파이펫으로 흡입하여 제거한 다음 폐기하십시오.
- 실험실 책에서 신경관 절제 수준을 기록합니다. 호스트 태아는 지금 기증자 신경관을 수신할 준비가 되었습니다.
5. 기증자 이식 조직 준비
- FITC 필터를 사용하는 형광 스테레오 현미경으로 볼 수 있는 창, 스테이지 일치 GFP 배아를 선택합니다. GFP 형광은 소닉을 시각화하고 배아를 단계화하는 것을 매우 쉽게 합니다.
- 단계일치 배아가 확인되면, 파셰프-울프 스프링 가위(도1C, l)를배아 주위의 직사각형 모양으로 4절개하여 달걀에서 배아를 제거한 다음, 배아 숟가락으로 부드럽게 집어들게 한다.
- 실가드 폴리머 베이스가 있는 사각형 시계 유리에 배아를 놓습니다. 두몬트 #5 핀셋으로 배아를 부드럽게 흔들어 부착된 노른자를 제거합니다. 비텔린 멤브레인을 제거하고 스테인레스 미노티엔핀(도 1C)을사용하여 배아를 폴리머 베이스에 고정한다.
- 스프링 가위를 사용하여, 주조 배아에서 제거 된 동일한 영역에서 신경관 과 주변 소미트 주위에 직사각형 모양으로 4 절개를합니다.
- 플라스틱 이송 파이펫을 사용하여 기증자 GFP 배아에서 펜/스트렙 PBS에서 0.2%의 췌장이 들어있는 시계 유리로 신경관 및 소미트 조직을 이송합니다.
- 효소 소화가 RT에서 10분 동안 진행하여 조직을 분리하도록 허용합니다. 효소에서 인큐베이션 후 손잡이에 장착된 스테인리스 미노티엔 핀을 사용하여 모든 인접한 조직으로부터 신경관을 수동으로 분리하십시오.
- 유리 마이크로 파이펫을 사용하여, DMEM + 10 % 혈청(예 :염소, 말 또는 태아 송아지)을 포함하는 다른 시계 유리로 해리 된 신경 관을 얼음에 옮기고, 여분의 췌장을 헹구고 효소 소화를 중지합니다. 5 분 후, 해부 신경 관은 병아리 호스트(도 2 및 S1)에정형 소식으로 이식 할 준비가되어 있습니다.
6. 조직 접목
- 유리 마이크로 파이펫을 사용하여 해부된 신경관을 시계 유리에서 숙주 배아로 조심스럽게 이송합니다. 신경관을 올바른 전방 후방 방향으로 배치하고 마이크로 메스를 사용하여 병아리 숙주의 절제 부위에 인접한 엑셀을 부드럽게 밀어 넣습니다. 신경관의 방향을 확인하기 위해 등대 표면에 작은 닉을 자르거나 작은 닉을 절단하여 작은 자궁 절제술조각을 둡니다.
- 필요한 경우 마이크로 메스를 사용하여 절제된 영역의 정확한 크기로 엑셀을 다듬습니다.
- 신경관을 압착 부위로 부드럽게 안내하고 등쪽측이 올바르게 방향을 지정하는 것을 배치합니다. 입 튜브에 장착된 유리 마이크로 파이펫을 사용하여 접목을 둘러싼 PBS 및/또는 유체를 제거합니다. 이것은 기증자와 호스트 조직이 준수하고 접목이 확립되는 것을 돕습니다.
- 탈수및 오염을 방지하기 위해 24mm 너비의 투명 테이프로 창 전체를 밀봉하십시오.
- 달걀 껍질에 연필로 표시하여 키메라 배아에 라벨을 부착하고 실험실 책에 그 숫자를 기록합니다. 추가 개발을 위해 인큐베이터에 계란을 반환합니다.
7. 호스트 배아의 혈관에 DiI주입
- 원하는 실험 시점(여기서 3-10일 후)에서 인큐베이터에서 키메라 배아를 회수하고 직선 가위를 사용하여 투명 테이프를 제거하여 계란 내의 배아에 접근할 수 있습니다.
- 필요한 경우 가위를 사용하여 셸의 창을 확대합니다. 껍질에 부착되어 출혈을 초래하고 혈관 라벨링을 위태롭게 하는 조개막을 손상시키지 않도록 주의하십시오.
- 노른자에서 혈액 흐름이 배아를 향해 향하도록 접근 가능한 정맥을 선택하십시오. 진동 정맥 중 하나의 분기점을 선택합니다(그림3B, C).
참고: E6.5 - E7.5에서, 초리알란토믹 멤브레인은 노른자 정맥에 접근하기 위해 핀셋으로 부드럽게 옆으로 이동해야 할 수도 있습니다. E8.5 이후, 유일한 옵션은 이 단계에 의해, chorioallantoic 막이 완전히 배아를 커버하기 때문에 chorioallantoic 막 정맥 의 하나에 주입하는 것입니다. - 두 개의 두 개의 두 #5 개의 두 개의 핀셋을 사용하여 선택한 주입 점 위의 비텔린 멤브레인을 제거하여 반대 방향으로 찢어.
- Dumont #5 사용하여 당겨진 유리 바늘을 깨고 CellTracker CM-DiI로 적재하기 전에 정맥의 대략적인 크기로 직경을 조정합니다. DMSO의 40 μg/μl에서 DiI 스톡 솔루션을 만들고 -20°C에 보관하십시오. 작업 용액을 0.3 M 자당/PBS로 4 μg/μl 농도로 준비합니다.
- 입 튜브를 사용하여 바늘로 0.3 M 자당 / PBS에서 DiI의 5 - 10 μl 사이를 흡입합니다. 더 오래된 태아는 25 μl 이상을 요구할 수 있습니다. E8.5에서 배아는 DiI로드 유리 바늘로 찌르기 전에 더 큰, 더 많은 근육 정맥이 #5 위치에 개최될 필요가 있을 지도 모릅니다.
- 바늘을 정맥에 신속하게 삽입하고 입 튜브로 꾸준히 날려 DiI가 혈전을 형성하지 않고 천천히 혈류에 참여할 수 있도록 합니다. 또는 DiI 전달을 위해 압력 인젝터를 사용하십시오.
8. 단면 또는 전산 검사를위한 배아를 수확
- 가능한 한 많은 DiI를 배아 내에서 유지하려면, 천공 숟가락에 스쿠핑하고 혈관과 직선 가위 쌍과 결합 조직을 절단하여 주입 직후 배아를 수확하여 노른자에서 배아를 해방하십시오.
- 느슨한 막을 제거하고 관심기관(즉,이 튜토리얼의폐 및 소화관)을 해부하여 DiI의 확산을 일으키는 조직을 압축하지 않도록 주의를 기울입니다. RT에서 1-2 시간 동안 4 % PFA에 침수하여 조직을 즉시 수정하십시오.
- PBS에서 5분 동안 조직을 헹구고, PBS에서는 5μg/ml DAPI를 함유합니다. 전체 마운트 검사를 위해 브리지 현미경 슬라이드에 샘플을 장착하거나 냉동 절위를 위해 샘플을 포함시킴이 에 포함시킴이 됩니다.
Representative Results
도 1은 신경관의 미세 수술 격리 및 이식을 수행하는 데 필요한 전형적인 기기를 나타낸다. 도 2는 이식 절차를 나타낸다. 이식 배아는 이식 성공을 위해 검열됩니다. 이것은 접목 유래 (GFP+) NCC의 존재를 위해, 일반적으로 미세 수술 후 아침 스테레오 형광 현미경의 밑에 태아를 검토하는 관련시킵니다. 이식이 성공적이라면 GFP+ NCC는 신경관 부근과 전구로 이어지는 초기 이동 경로에서 관찰될 수 있습니다. 절차가 성공하지 못하면 GFP+ NCC는 신경관 외부에서 관찰되지 않거나 호스트에 있는 경우 더 적은 수의 경우 관찰될 수 있습니다. 이 실패한 태아는 버려지않습니다. 전형적으로, 5-8 신경관 이식은 하루에 수행되고, 이 80%는 성공적입니다. 신경관 이식에 실패한 이유는 미세 수술 중에 발생한 조직 손상으로 인한 배아의 죽음, 또는 숙주 배아에 통합하는 신경관의 실패를 포함한다. 후자는 숙주 내의 신경관배치 불량 또는 열악한 해부 기술 또는 해리 효소에 과도하게 노출되어 있기 때문에 품질이 좋지 않아 발생할 수 있습니다. GFP+ 세포에 대한 초기 검사 단계뿐만 아니라 유사한 후기 검사는 장 내에 GFP 라벨이 부착된 NCC가 없는 배아에 대한 실험을 수행함으로써 시간과 자원이 낭비되지 않는다는 것을 의미하기 때문에 유용합니다.
도 3은 혈관의 DiI 주입 절차를 나타낸다. DiI 주입 기술의 효율성/성공은 먼저, 표적 정맥에 대한 최적의 직경으로 주사 바늘을 절단하고, 정맥에 바늘을 삽입할 때 정확한 제스처를 두 번째로 (다른 쪽을 관통하지 않음), 세 번째는 일정한 속도로 불어 주입 하는 동안 바늘이 연결되는 것을 피한다. 이 세 가지 매개 변수 중 어느 것이 잘못 수행되면, 배아는 출혈이 발생하기 전에 회복하는 데 몇 시간이 필요하거나 출혈로 인해 즉시 다시 주입하는 것이 거의 불가능합니다. 성공적인 배아는 입체형 현미경의 밑에 보기에 의해 즉시 선택되어야 하고 급속하게 해부되어야 합니다. 성공적인 배아에서, DiI 표지혈관은 모세혈관(도3D)을포함하는 배아(도3C,D)를통해 존재한다.
배아를 수확하고 조직 섹션 또는 전체 산 위장관의 검사시, 전형적인 결과는 원시적인 ENS 내의 GFP+ NCC및 DiI 표지된 창자 혈관 네트워크의 미세구조(도4)전신탈 제제는 공초점 현미경검사를 사용하여 검사될 수 있으며, 이에 따라 이미지 스택이 GFP+ ENS 세포의 미세투영과 DiI 염색 혈관 계통 간의 상호 관계를 보여주는 3차원(3D) 재구성을 생성한다(그림4; A-C. G-I; 동영상 1과 2).

그림 1. 권장 미세 수술 기구. (A) 바느질 바늘로 모양의 마이크로 메스. (B) 미세 메스를 형성하기위한 고급 아칸소 돌. (C) 직선 가위, b) 곡선 가위, c) 181/2 G 피하 바늘, d) 플라스틱 파이펫, e) 사용자 정의 만든 계란 홀더, f) 검은 잉크, g) 사각형 시계 유리, h) 검은 실가드 베이스, i) 바늘 홀더에 마이크로 메스, j) minutien 핀, k) 미누티엔 또는 텅스텐 바늘 홀더, l) Pascheff-Wolff 봄 가위, m) 듀몬트 #5 핀셋, n) 천공 숟가락, oi) 짧은 불-모는 바늘, oi) 바늘 을 뽑아. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 2. 종 내 신경 관 이식. 병아리 배아/GFP 신경관 이미지는 델랄란데 외에서수정되었습니다. 12.혈관화는 장내 신경 문장 세포에 의한 장 식민지화에 필요하지 않습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 3. 정맥 내 DiI 주입. (A) 권장 기기: a) 셀트래커 CM-DiI 파라필름에 드롭, b) 유리 주입 바늘, c) 입 튜브를 뽑아. (B) E4 키메라 병아리 배아에 정맥 내 DiI 주입의 회로도 도표. (C) 오보 디이 정맥 주사에서 정맥(arrow)에 삽입된 DiI를 함유한 미세유리 바늘을 나타낸다. (D) GFP+ 신경관(arrow)을 가진 E4 키메라 배아 포스트 DiI 주입(red). (E) DiI는 살아있는 태아에서 미세 혈관 네트워크를 염색, 24 시간 후 주입. Br: 뇌; H: 심장; LB: 사지 싹; A: 알란토이스. (C) 및 (D)의 이미지는 Delalande 외에서수정되었습니다. 12 혈관은 장 내 신경 문장 세포에 의해 창 자 식민지에 대 한 필요 하지 않습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 4: E5.5 병아리 배아의 위장과 케이쿰을 대표합니다. (A-C) 3차원(3D) GFP+ 장내 신경문장 세포(ENCC)(ENCC)를 보여주는 위 부위에 공초점 이미지 스택의 재구성(E) DiI 염색 혈관 계통 및 (F) 위 의 수준에서 D-F 조직학적 섹션의 병합된 이미지 (G) GFP+ ENCC(H) 양상과 디큘러(H) 양탄자 및 혈관의 배아(H) 결합된 배의 배다(H) 및 혈관 의 양상(H) 및 혈관 의 원반(H) 및 혈관 의 원고형(H) 결합된 영상( H) 및 혈관 의 원고형(H) 결합된 배암체 시스템. 핵은 DAPI (시안)로 염색됩니다. (G-H) (G-H) 녹색으로 GFP+ ENCC 마이그레이션 전면을 보여주는 케이쿰 영역에서 공초점 이미지 스택의 3D 재구성, (B) DiI 염색 혈관 시스템은 빨간색으로, (C) 두 네트워크의 병합 된 이미지. 이미지(A-F)는 델랄랜드 등에서수정되었습니다. 12 혈관은 장 내 신경 문장 세포에 의해 창 자 식민지에 대 한 필요 하지 않습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

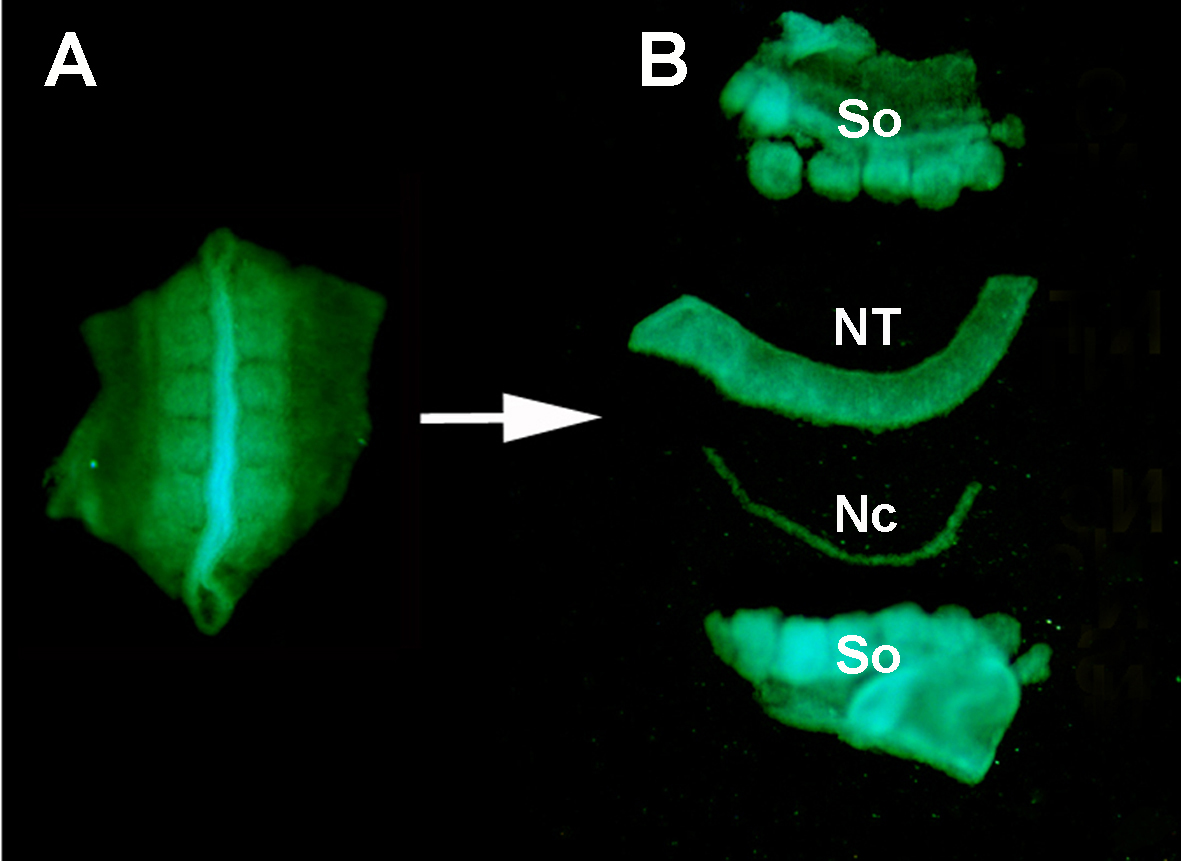
그림 S1. 효소 소화 및 미세 해부에 의해 주변 조직에서 기증자 GFP + 신경 관의 격리. (A) 기증자 배아로부터 해부된 GFP+ 신경관 및 인접 소미트. (B) 스테인레스 미노티엔 핀을 사용하여 췌장 소화 및 미세 해부 후 분리 된 신경 관. 그래서: 소미; NT: 신경관; NC: 노토코드.

비디오 1. 도 4C에서 이미지의 3차원 360° 회전, E5.5(HH27-28)에서 위장의 혈관 시스템과 ENCC를 나타낸다. 이 비디오를 보려면 여기를 클릭하십시오.

영상 2. 도 4I에서 영상의 3차원 360° 회전, E5.5(HH27-28)에서 케이쿰 의 영역에서 혈관 시스템 및 ENCC 이동 전면을 보여 주는 영상. 이 비디오를 보려면 여기를 클릭하십시오.
Discussion
여기에 설명된 혈관 라벨링과 결합된 종내 신경관 이식 방법은, 뇌신경계(ENS)와 혈관 계통의 원소의 공동 개발을 연구하기 위해 계란 내의 조류 배아의 접근의 용이성을 최대한 활용한다.
NCC 유도체라벨링의 경우, 우리가 설명하는 병아리GFP-병아리종 이식 방법은 40년 전1-3이상확립된 고전적인 메추라 병아리 키메라 방법에 비해 여러 가지 장점이 있다. 첫째, FITC 빛 하에서 GFP 형광은 매우 밝으며, GFP+ 세포가 살아있는 키메라 배아에서 쉽게 식별할 수 있는 정도까지. 이를 통해 접목의 성공은 오보에서검사될 수 있는 반면, 메추라기 병아리 이식은 접목의 성공이 확인되기 전에 QCPN을 사용하여 배아를 죽이고, 가공하고, 면역염색해야 하는반면, 2. 둘째, 형질형 병아리GFP에서의 GFP 발현은 세포세포에 라벨이 붙을 뿐만 아니라 이식된 세포의 투영을시각화할 수 있도록 한다.22. 이것은 복잡한 신경 망이 고해상도에서 관찰될 수 있게 합니다(시료가 항-GFP 항체로 면역염색될 때 미세 한 프로젝션이 가장 잘 시각화된다는 점에 유의하십시오). QCPN 라벨링은 메추라기 세포 핵으로 제한되기 때문에, 이러한 네트워크는 메추라 병아리 키메라를 사용하여 공개되지 않습니다. 셋째, 이식종 이식은 키메라 배아 내의 세포 들 간의 잠재적인 종 차이를 제거합니다. 메추라기 배아는 병아리 (19 일 대 21 일)보다 더 짧은 잠복기 기간을 가지고 있기 때문에 메추라기 세포가 병아리 세포보다 더 높은 증식률을 가지고 있으며, 이는 잠재적으로 키메라 조직의 발달에 영향을 미칠 수있습니다(23). 흥미롭게도, 종 간 접목이 숙주 24에서DNA 메틸화 패턴에 광범위한 변화를 일으킬 수 있다는 식물에서 나타났습니다. 넷째, 병아리GFP는 NCC 운명및 세포 약정25와같은 주제를 해결하기 위해 백 이식 실험을 용이하게한다. 다섯째, 형질전환 병아리GFP는 GFP+ 세포 하위 인구의 FACS 선별, GFP+ 세포를 포함하는 기관의 조직화 배양, 발현 플라스미드(26)의 전분화를 통한 GFP+ 이식된 조직의 유전자 조작, 광학 프로젝션 단층촬영(27)과같은 기타 이미징 기술에도 유용하다.
신경관 이식 접근법은 미세 외과적으로 신경관의 더 짧은 양을 대체하여 수정될 수 있습니다. 신경관의 작은 세그먼트를 사용하여 미세 수술은 잠재적으로 태아에 덜 손상되고 생존이 향상될 수 있다. 그러나, 덜 신경관을 이식하는 단점은 호스트에 있는 GFP+ NCC의 수가 감소될 것이라는 것입니다. 사용자는 배아의 최적 생존을 제공하기 위해 이식 된 신경 관의 양과 유익한 결과를 제공하기에 충분한 숙주 장 내의 GFP + NCC 의 수 사이의 균형을 시도하고 달성 할 수 있습니다.
선박 페인팅의 경우 DiI는 형광이 매우 밝고 견고하다는 장점이 있습니다. 또한, 고정 시 확산이 가능한 능력을 가지고 있어 최고급 개포모세혈관의 염색을 보장합니다. 중요한 염료이기 때문에, 배아는 주입 절차를 생존하고 스테인드 혈관 시스템으로 개발을 계속할 수 있습니다 (우리의 손에 최대 24 시간, 염색은 시간이 지남에 따라 더 많은 천공이되고 있지만, 그림 3E참조). 따라서 병아리GFP접목과 DiI 혈관 페인팅의 조합은 라이브 이미징과 호환됩니다. 이러한 모든 장점 외에도 혈관 주사는 발광 용기만 표시하므로 미개봉 모세 혈관, 내피 팁 세포 또는 격리 된 내피 세포를 식별하지 않는다는 점에 유의하는 것이 중요합니다. 그러나, 조류 간 유전자의 추가 진전은 혈관 형태발생(28)을연구하기 위해 Tg(tie1:H2B-eYFP) 메추라기 배아를 이용한 실험에 의해 예시되는 바와 같이 이러한 문제를 우회하는 새로운 방법을 제공할 수 있다. 이 기술의 또 다른 한계는, E7.5 이상에 태아에 있는 효과적인 혈관 표지를 위해, 염료의 더 많은 양을 주입할 필요가 있다는 것입니다, 실험을 비싸게 만들 수 있는. 그러나, 기술의 수정은 하이라이터 잉크를 라벨링 하는 저비용 혈관을 포함할 수 있습니다14,비록이 접근은 우리의 손에 시도 되지 않았습니다.
절차의 중요한 단계는 blastodisc 의 밑에 잉크를 주입하여 태아를 시각화하는 과정을 포함합니다. 노른자를 덮는 멤브레인이 단계에서 잉크로 채워진 바늘에 의해 찢어지면 배아 생존이 심각하게 손상됩니다. 또한, 기증자 신경관을 준비할 때, 조직이 췌장에서 지나치게 오랜 시간 동안 방치되지 않는 것이 중요합니다(최대로 약 10분 고려). 췌장에 장기간 노출되면 조직이 손상되고 신경관은 처리하기가 어렵고 호스트에 잘 통합되지 않습니다. 야생 형 배아에 DiI 주입 기술의 경험을 얻는 것은 키메라 배아를 주입하기 전에 필수적이다, 각 배아에 대한 일반적으로 가능한 주입에 하나의 시도가 있기 때문에. DiI 부피 및 바늘 직경은 각 배아에 대한 중요한 매개 변수이며 야생 유형, 스테이지 일치 제어에 대해 평가되어야합니다.
결론적으로, 살아있는 병아리 배아에 있는 신경관 이식 및 DiI 혈관 그림의 우리의 이중 라벨링 방법은 유기발생 도중 NCC와 혈관 네트워크 사이 상호 관계를 조사하기 위하여 이용될 수 있습니다. 장기 발달 중 올바른 표적 내분 및 혈관화를 확립하는 메커니즘을 고려하면 여전히 크게 알려지지 않았으며,이 방법론은이 분야에서 미래의 발견을위한 잠재력을 보유하고 있습니다.
Disclosures
저자는 경쟁적인 재정적 이익이 없다고 선언합니다.
Acknowledgments
수정된 GFP 치킨 계란은 헬렌 상 교수, 로슬린 연구소 및 영국 에든버러 대학에 의해 공급되었습니다. 로슬린 트랜스제닉 치킨 시설은 웰컴 트러스트와 생명 공학 및 생물 과학 연구 위원회 (BBSRC)에 의해 투자됩니다. 이 사업은 부분적으로 지원되었고, NT는 영국 런던 의 그레이트 오몬드 스트리트 병원 어린이 자선 단체에 의해 지원되었습니다. 저자는 벤 Jevans, 아동 건강의 UCL 연구소, 접목에 대 한 배아를 준비에 대 한 도움에 대 한 감사.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}