

Summary
Здесь мы сообщаем о двойной маркировке нервных клеток гребня и кровеносных сосудов спомощью внутривидовой внутривидовой клетки GFP в сочетании с внутрисосудистой инъекцией ДиИ. Этот экспериментальный метод позволяет одновременно визуализировать и изучать развитие нервной системы и сосудистой системы, полученной из NCC, во время органогенеза.
Abstract
Все развивающиеся органы должны быть подключены как к нервной системе (для сенсорного и двигательного контроля), так и к сосудистой системе (для газозамена, жидкости и питательных веществ). Следовательно, нервная и сосудистая системы развиваются бок о бок друг с другом и имеют поразительное сходство в их разветвленной архитектуре. Здесь мы сообщаем о эмбриональных манипуляциях, которые позволяют изучать одновременное развитие нервной ткани нервной ткани (в данном случае и кишечная нервная система) и сосудистой системы. Это достигается путем генерации куриных химер путем трансплантации дискретных сегментов нервной трубки, и связанных с ними нервных гребней, в сочетании с сосудистой инъекцией ДиИ в том же эмбрионе. Наш метод использует трансгенныеэмбрионы цыпленка GFP для внутривидовой прививки, что делает технику трансплантации более мощной, чем классический межвидовой протокол прививки перепелиного птенца, используемый с большим эффектом с 1970-х годов. ChickGFP-chick внутривидовой прививки облегчает визуализацию пересаженных клеток и их прогнозы в нетронутых тканях, и устраняет любые потенциальные предубеждения в развитии клеток, связанные с видами различия. Этот метод в полной мере использует легкость доступа птичьего эмбриона (по сравнению с другими позвоночными эмбрионами) для изучения совместного развития кинеральной нервной системы и сосудистой системы.
Introduction
Куриный эмбрион является бесценной моделью организма в биологии развития позвоночных, не в последнюю очередь потому, что его развитие в ово позволяет экспериментальные манипуляции, которые в противном случае невозможно выполнить у позвоночных, которые развиваются в утробе матери. Такая доступность и простота манипуляций привели к тому, что эмбрион цыпленка сыграл ключевую роль во многих семенных открытиях в области биологии развития. Среди наиболее мощных методов было использование перепела цыпленок химерных эмбрионов для изучения судьбы клеток, метод, впервые профессор Николь Ле Douarin в 1970-х1-3. В частности, перепелино-куриные химерки были особенно полезны для генетической отметки и следовать высоко мигрирующих нейронных клеток гребня (NCC) популяций во время раннего развития. NCC - это многопотентная популяция мигрирующих клеток, возникающих в спинном эктодерме на полях нервной трубки, которые дают возможность для широкого спектра типов клеток по всему эмбриону позвоночных. К ним относятся черепно-мозговые структуры (хрящ, кости, мышцы), нейроны и глиа (в сенсорной и вегетативной нервной системе), меланоциты, а также субпопуляцияклеток эндокринной системы 2,4,5. Одним из наиболее важных факторов, влияющих на судьбу NCC является их первоначальное расположение вдоль передней-задней оси нервной трубки. Например, энтерический NCC, которые дают возможность нейронов и глии кинеральной нервной системы (ENS), возникают из двух дискретных поднаселения: первый расположен в области вагала (каудального заднего мозга), а второй в сакральной областинервной трубки 6-13. Интер или внутривидовой прививки соответствующих областей нервной трубки были методы выбора постоянно маркировать эти клетки, а затем позволяют отслеживать, от их рождения на полях нервной трубки, к их конечной назначения впищеварительном тракте 6,7,10.
Еще одна эмбриональная манипуляция, которую легче выполнять в птенце, по сравнению с другими моделями животных, является жизненно важной маркировкой сосудистой системы. Действительно, по мере развития эмбриона птенца, он лежит на верхней части вне эмбриональной сосудистой сети, которая циркулирует кислород и питательные вещества из желтка. Эта доступная сосудистая сеть, расположенная на поверхности желтка, может быть использована в качестве шлюза для обозначения развивающейся сосудистой системы эмбрионаво время органогенеза 12,14-17. Внутрисосудистая инъекция различных красителей, таких как липофилинический краситель DiI, позволяет очертить/запятнать все осветленные сосуды зарождающейся сосудистой сети.
Поскольку развивающиеся органы должны быть подключены как к нервной системе (для сенсорного и двигательного контроля), так и к сосудистой системе (для газозаменения, подачи жидкости и питательных веществ), обе сети развиваются бок о бок друг с другом и имеют поразительное сходство в их разветвленнойархитектуре 18-20. Здесь мы сообщаем о эмбриональных манипуляциях, которые позволяют нам изучать одновременное развитие NCC-производных ENS, наряду с сосудистой системой, во время органогенеза. Это достигается путем генерации куриных химер путем трансплантации дискретных сегментов нервной трубки, включая нервный гребень, в сочетании с сосудистой инъекцией DiI. В качестве аванса от перепелиных куриных химер, наш метод использует трансгенные эмбрионы GFP цыпленка для внутривидовой прививки, что делает технику трансплантации более мощным, с точки зрения визуализации клеток и их проекций, и устранение любых потенциальных предубеждений, связанных с различиями видов.
Protocol
1. Подготовка микро-скальпеля для аблейтаций нервной трубки
- Сформировой микро-скальпель из коммерчески доступной стальной швейной иглы.
- Сначала сгладить иглу с обеих сторон с помощью шлифовального колеса, установленного на питание скамейке шлифовальной машины.
- Начните формировать скальпель, сначала на грубой степени Арканзас камень с помощью контролируемого кругового движения, в альтернативных направлениях, по обе стороны от иглы.
- Носите на том же заточки движений на дополнительный тонкий камень класса Арканзас, чтобы сформировать ультра тонкой микро-скальпель, с четко определенным передний край (Рисунок 1A, B).
ПРИМЕЧАНИЕ: Альтернативы микро-скальпель может быть электролитически заточенные иглы, коммерчески доступны вольфрама иглы, или вытащил стеклянные иглы.
2. Инкубировать дикий тип и GFP яйца на нужную стадию
- Храните оплодотворенные куриные яйца и трансгенные куриные яйца GFP в охлажденом инкубаторе при температуре 14 - 15 градусов цельсия до инкубации, так как развитие при такой температуре останавливается. Храните яйца в течение нескольких дней, до одной недели.
- Чтобы начать разработку, поместите яйца дикого типа и GFP на поднос горизонтально и инкубировать одновременно во влажном (58 - 60%) инкубатор при 37,5 oC, так что эмбрионы находятся на соответствующих стадиях для пересадки нервной трубки.
- Для получения эмбрионов на стадии 10 - 12 сомитов для проведения пересадки вагалькой нервной трубки, инкубации яйцеклеток в течение 1,5 дней (33 - 38 часов) и стадии эмбрионов в соответствии с таблицами развития Гамбургер и Гамильтон21.
3. Подготовка яиц для окон и прививки
- Перемещение одного яйца в то время, на заказ держатель яйца для окон. Сделайте небольшое отверстие в яичной скорлупе, неоднократно нажав, с прямыми ножницами, на верхней поверхности остроконечного конца яйца.
- Удалить 2 - 3 мл альбумин из яйца с 181/2 G подкожной иглой и 5 мл шприца. Удаление альбумин снижает желток в яйцеклетке и облегчает последующее оконное окно, не вызывая каких-либо повреждений эмбриона.
- Отбросьте альбумин. Печать отверстие с небольшой полосой ясной лентой вырезать до размера с мелкими ножницами.
- Используя изогнутые ножницы, нажмите другое отверстие в верхней поверхности яичной скорлупы. Вставьте кончик ножниц в отверстие и, держа ножницы параллельно скамейке, работа в круговом движении, чтобы вырезать окно диаметром 2 см на верхней части корпуса.
- Держите ножницы в неподвижном положении и поверните яйцо. Отбросьте удаленный диск яичной скорлупы. На E1.5 эмбрион узнаваем как более темный желтый диск поверх желтка.
- Удалите любые обломки оболочки, которые упали в яйцо с помощью пинцета. Отбросьте любые неутилизованные яйца (идентифицированные небольшим белым пятном сверху на светло-желток).
4. Подготовьтесь к тому, чтобы эмбрион-хозяин получил привитую ткань
- Отрегулируйте стерео-микроскоп до уровня глаз и оптимизируйте ориентацию гусиного источника света, чтобы адекватно осветить эмбрион, не вызывая отражений.
- Чтобы визуализировать эмбрион надлежащего, вводить небольшое количество индийских чернил под центром темного желтого диска, используя рот трубки и вытащил стекла микро-пипетки (Рисунок 1C, Oii).
- Подготовка чернил 50:50 с PBS, содержащий пенициллин / стрептомицин при 100 мкг / мл окончательной концентрации. Вставьте микро-пипетку через желтковую мембрану за периметр бластодерма, затем осторожно угол его кончик прямо под эмбрионом.
- Доставка чернил под эмбрион, дуя на рот трубки. Если трубка для рта не допускается, используйте вместо этого шприц по 1 мл. Будьте осторожны, чтобы не ввести пузырьки воздуха под эмбрионом, что может привести к загрязнению, а затем тщательно удалить стекло микро-пипетки. Это деликатный шаг, который может привести к смерти эмбриона, если не сделать с точностью.
- Этап эмбриона со ссылкой на Гамбургер иГамильтон 21 и записать этап в лабораторной книге.
- Используя изготовленный на заказ микро-скальпель (или тонкую вольфрамовую иглу), установленную на держателе иглы, сделайте очень маленькую газу в вителлиновой мембране, рядом с областью, где будет проведена микрохирургия.
- Тщательно нанесите 2 - 3 капли PBS на мембранную слезу (с помощью стеклянной микро-пипетки и трубки рта), чтобы создать пространство между эмбрионом и мембраной. Вырежьте большее окно в мембране, чтобы разоблачить весь регион, где микро-хирургия будет проходить.
- Удалите область нервной трубки, представляющих интерес с помощью микро-скальпеля, начиная с ростральных и каудальных поперечных разрезов по всей спинной нервной трубки (на уровне сомита от 1 до 7 в видео).
- Вырезать на двусторонней основе между нервной трубки и somites отделить нервной трубки от окружающих тканей, не повреждая сомитов.
- Очень аккуратно отделить нервную трубку от основного нотохорда, который должен оставаться нетронутым. Обратите внимание, что успешное иссечение нервной трубки оставит все окружающие ткани совершенно нетронутыми(рисунок 2).
- Удалите вырезанную нейронную трубку, аспирируя ее в стеклянную микро-пипетку, а затем отбросьте.
- Замеская уровень абляции нервной трубки в лабораторной книге. Эмбрион-хозяин теперь готов к получению донорской нервной трубки.
5. Подготовка донорской ткани трансплантата
- Выберите оконный, сценический эмбрион GFP, просмотрев под флуоресцентным стерео-микроскопом с фильтром FITC. Флуоресценция GFP позволяет очень легко визуализировать сомиты и этап эмбриона.
- После того, как стадия соответствует эмбриона была определена, удалить эмбрион из яйца, сделав 4 разреза, с Pascheff-Wolff весенние ножницы (Рисунок 1C, l) в прямоугольной форме вокруг эмбриона затем осторожно забрать его с ложкой эмбриона.
- Поместите эмбрион в квадратное часовое стекло с силгардным полимерным основанием. Аккуратно встряхните эмбрион с помощью пинцета дюмона #5, чтобы удалить любой прикрепленный желток. Удалите вителлиновую мембрану и прикрепите эмбрион к полимерной основе с помощью булавок из нержавеющей мелочи(рисунок 1C).
- Используя весенние ножницы, сделайте 4 разреза в прямоугольной форме вокруг нервной трубки и окружающих сомитов, в той же области, которая была удалена из эмбриона хозяина.
- Используя пластиковую трубу передачи, передача нервной трубки и тканей сомита от эмбриона донора GFP в часы стекла, содержащие 0,2% поджелудочной железы в Pen / Strep PBS.
- Разрешить энзиматический пищеварения продолжать в течение 10 минут на RT, чтобы помочь отделить ткани. После инкубации фермента используйте булавки из нержавеющей минуты, установленные на ручке, чтобы вручную отделить нервную трубку от всех соседних тканей.
- Используя стеклянную микротрубку, перенесите диссоциированную нейронную трубку в другое часовое стекло, содержащее DMEM и 10%сыворотки(например, коза, лошадь или фетальный теленок) на льду, чтобы промыть избыток панкреатина и остановить ферментативное пищеварение. Через 5 минут, вскрытая нервная трубка готова быть привитых ортопедически в цыпленокхозяина (рисунок 2 и S1).
6. Привить ткань
- Используя стеклянную микро-пипетку, тщательно перенесите вскрытую нейронную трубку из часового стекла в эмбрион хозяина. Распоистите нервную трубку в правильной передней-задней ориентации и аккуратно нажмите explant рядом с вырезанной области цыпленка хозяина с помощью микро-скальпеля. Оставьте небольшой фрагмент эктодерма прилагается к, или путем резки небольшой ник в спинной поверхности для определения ориентации нервной трубки.
- При необходимости используйте микро-скальпель, чтобы обрезать эксплант до точного размера вырезанной области.
- Аккуратно направляйте нейронную трубку в абляционную область и распоивте ее так, чтобы спинная сторона была правильно ориентирована. Используйте стеклянную микро-пипетку, установленную на трубке рта, чтобы удалить PBS и/или жидкость, окружающую трансплантат. Это помогает донору и принимающим тканям придерживаться и трансплантата, чтобы стать создана.
- Печать всего окна с 24 мм в ширину ясной лентой для предотвращения обезвоживания и загрязнения.
- Этикетка химерного эмбриона, отмечая карандашом на яичной скорлупе и записывать его число в лабораторной книге. Верните яйцо в инкубатор для дальнейшего развития.
7. Ввимить диИ в кровеносные сосуды эмбриона хозяина
- В желаемую экспериментальную точку времени (здесь, 3 - 10 дней спустя), получить химерный эмбрион из инкубатора и удалить четкую ленту с помощью прямых ножниц, чтобы получить доступ к эмбриону в яйцеклетке.
- При необходимости, увеличить окно в оболочке с помощью ножниц. Будьте осторожны, чтобы не повредить chorioallantoic мембраны, если он прикреплен к оболочке, что приведет к кровоизлиянию и поставить под угрозу маркировку кровеносных сосудов.
- Выберите доступную вену на желтке, убедившись, что кровоток направлен к эмбриону. Выберите точку ветвления одной из вителлинных вен(рисунок 3B, C).
ПРИМЕЧАНИЕ: На E6.5 - E7.5, chorioallantoic мембраны, возможно, потребуется аккуратно отойти в сторону с пинцетом для доступа желток вены. После E8.5, единственный вариант заключается в том, чтобы ввести в один из chorioallantoic мембранных вен, поскольку, к этому этапу, chorioallantoic мембраны полностью покрывает эмбрион. - Удалите вителлиновую мембрану над выбранной точкой впрыска с помощью двух пинцетов Dumont#5 разрывая в противоположных направлениях.
- Разбейте вытянутую стеклянную иглу с помощью #5 и отрегулируйте ее диаметр до приблизительного размера вены перед загрузкой CellTracker CM-DiI. Сделайте раствор diI на складе при 40 мкг/зл в DMSO и храните при -20 градусах Цельсия. Подготовь рабочий раствор в 0,3 М сахарозы/PBS с концентрацией 4 мкг/хл.
- Аспират между 5 - 10 мкл ДиИ в 0,3 М сахарозы / PBS в иглу с помощью всасывания с ротовой трубки. Более старые зародыши могут требовать до 25 йл или больше. От E8.5, зародыши имеют более большие, более мышечные вены, которые могут быть притужены в положении с думонским #5 перед колоть с DiI-нагруженной стеклянной иглой.
- Быстро вставьте иглу в вену и удар неуклонно с ротовой трубки, чтобы позволить DiI присоединиться к кровотоку медленно, не образуя сгусток. Кроме того, используйте инжектор давления для доставки DiI.
8. Урожай эмбрионов для секциона или Wholemount экспертизы
- Чтобы сохранить как можно больше ДиИ в эмбрионе, собрать эмбрион сразу после инъекции, зачерпнув его на перфорированную ложку и разрезая кровеносные сосуды и соединительные ткани с помощью прямых ножниц, чтобы освободить эмбрион от желтка.
- Удалите любые свободные мембраны и рассекают органы, представляющие интерес(т.е.легкие и пищеварительный тракт в этом учебнике), принимая большую осторожность, чтобы не сжимать ткани, что создает диффузию ДиИ. Немедленно исправить ткани путем погружения в 4% PFA в течение 1 - 2 часов на RT.
- Промыть ткань в течение 5 мин в PBS, затем 15 мин в PBS, содержащий 5 мкг / мл DAPI. Намонтировать образцы на мостовом микроскоп слайд для всего маунт экспертизы или вставлять их для крио-секции.
Representative Results
На рисунке 1 показаны типичные инструменты, необходимые для проведения микрохирургической изоляции и трансплантации нервной трубки. На рисунке 2 показана процедура трансплантации. После трансплантации эмбрионы проверяются на успешное пересадку. Это включает в себя изучение эмбриона под стерео флуоресценции микроскопа, как правило, на следующее утро после микрохирургии, на наличие трансплантата полученных (GFP) NCC. Если трансплантация была успешной, то GFP NCC можно наблюдать в непосредственной близости от нервной трубки и в ранних путей миграции, ведущих к foregut. Если процедура не была успешной, GFP NCC не будет наблюдаться за пределами нервной трубки, или если они присутствуют в хосте, они могут быть в меньших количествах. Эти неудачные эмбрионы отбрасываются. Как правило, 5-8 трансплантаций нервной трубки выполняются в один день, и из них 80% являются успешными. Причины неудачной трансплантации нервной трубки включают смерть эмбриона из-за повреждения тканей, понесенных во время микрохирургии, или отказ нервной трубки интегрироваться в эмбрион хозяина. Последнее может быть результатом плохого размещения нервной трубки внутри хозяина или из-за низкого качества нервной трубки из-за плохой техники вскрытия или от чрезмерного воздействия диссоциаation фермента. Первоначальный этап скрининга, а также аналогичные более поздние исследования для клеток GFP, является полезным, поскольку это означает, что время и ресурсы не впустую при проведении экспериментов на эмбрионах, которые не имеют GFP помечены NCC в кишечнике.
На рисунке 3 показана процедура инъекции ДиИ кровеносных сосудов. Эффективность/успех метода инъекций DiI зависит от: во-первых, резки иглы инъекции до оптимального диаметра для целевой вены, второй точный жест при вставке иглы в вену (так, чтобы не прокалывать другую сторону), и третий избежать иглы получать подключен во время инъекции, дуя с постоянной скоростью. Если какой-либо из этих трех параметров делается неправильно, эмбрион будет кровоточить или потребуется несколько часов, чтобы восстановиться, прежде чем вторая попытка сделана, как кровоизлияние делает его почти невозможно повторно вводить немедленно. Успешные эмбрионы должны быть выбраны немедленно при просмотре под стереофлюоресценцией микроскопа и должны быть вскрыты быстро. В успешных эмбрионов, DiI помечены кровеносные сосуды присутствуют по всемуэмбриону (рисунок 3C,D), включая капиллярные кровати (Рисунок 3D).
При сборе эмбрионов и обследовании секций тканей или целых желудочно-кишечных путей, Типичные результаты показывают, GFP NCC в примитивных ENS и тонкая структура DiI-маркированных сетей кровеносных сосудов кишечника(рисунок 4) Wholemount препараты могут быть рассмотрены с помощью конфокальные микроскопии которой изображения стеки производят трехмерные (3D) реконструкции, показывающие взаимосвязи между прекрасными проекциями клеток GFP' ENS и Окрашенные сосудистой системы DiI (Рисунок 4 A-C; G-I; Видео 1 и 2).

Рисунок 1. Рекомендуемые приборы микрохирургии. (A) микро-скальпель в форме швейной иглы. (B) прекрасный камень Арканзаса для формирования микро-скальпеля. (C) а) прямые ножницы, б) изогнутые ножницы, в) 5 мл шприца с 181/2 Г подкожной иглой, г) пластиковая пипетка, е) изготовленный на заказ держатель яйца, г) черные чернила, г) квадратные часы стекла, ч) квадратные часы стекла с черным силгард базы, я) микро-скальпель на держатель иглы, j) minutien булавки, к) minutien или вольфовой иглы на игле держатель, л) Pascheff-Wolff весенние ножницы, м) Дюмон #5 пинцет, п) перфорированной ложкой, oi) короткий огонь вытащил передачи иглы, о. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

Рисунок 2. Внутривидовая пересадка нервной трубки. Чик эмбриона / GFP нейронных изображений трубки были изменены из Делаланд и др. 12. Васкуляризация не является необходимым для колонизации кишечника кишечных нервных клеток гребня. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

Рисунок 3. Внутривенная инъекция дии. (A) Рекомендуемые инструменты: a) CellTracker CM-DiI капля на парафильме, б) вытащил стеклянную инъекцию иглы, в) рот трубки. (B) Схематическая диаграмма внутривенной инъекции DiI в эмбрион химерного птенца E4. (C) в ovo DiI внутривенной инъекции с указанием тонкой стеклянной иглы, содержащей DiI вставляется в вену (стрелка). (D) E4 химерных эмбрионов после инъекции DiI (красный) с GFP' нейронной трубки (стрелка). (E) DiI окрашенных тонкой сети кровеносных сосудов в живом эмбрионе, 24 часов после инъекции. Br: мозг; H: сердце; LB: бутон конечностей; A: allantois. Изображения в (C) и (D) были изменены из Делаланд и др. 12 Лет Васкуляризация не является необходимым для колонизации кишечника кишечных нервных клеток гребня. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

Рисунок 4: Представитель приводит к желудку и caecum эмбриона E5.5 цыпленка. (A-C) Трехмерная (3D) реконструкция конфокального стека изображения в области желудка, показывающего (D) клетки кишечного нервного гребня GFP (ENCC) (E) окрашенные сосудистую систему DiI и (F) объединенное изображение обеих сетей D-F Гистологических секций на уровне желудка, показывающего (G) GFP- ENCC (H) окрашенные в ДиИ сосуды и (I) изображение обеих сетей. Ядра окрашены DAPI (cyan). (G-H) 3D-реконструкция конфокального стека изображений в регионе caecum, показывающего (A) миграционный фронт GFP ENCC зеленым цветом, (B) окрашенную сосудистую систему DiI в красный цвет и (C) объединенное изображение обеих сетей. Изображения (A-F) были изменены из Делаланда и др. 12 Лет Васкуляризация не является необходимым для колонизации кишечника кишечных нервных клеток гребня. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

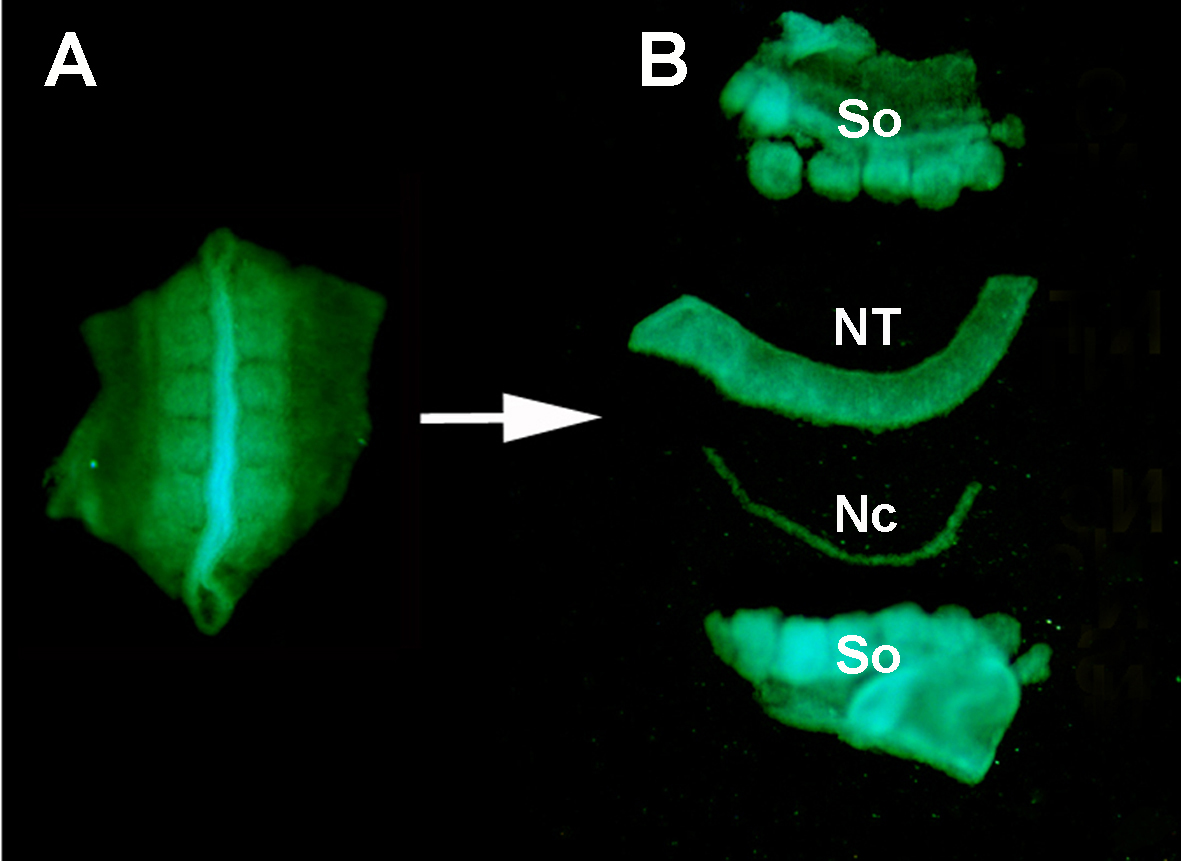
Рисунок S1. Изоляция донорской нервной трубки GFP' от окружающих тканей путем энзиматической пищеварения и микропарации. (A) GFP' нейронная трубка и прилегающие somites расчленены из эмбриона донора. (B) Изолированная нервная трубка после пищеварения поджелудочной железы и микро-рассечения с использованием нержавеющей минутиен булавки. Итак: somites; NT: нейронная трубка; Nc: Нонохорд.

Видео 1. трехмерное 360 "вращение изображения на рисунке 4C, показывая сосудистой системы и ENCC в желудке на E5.5 (HH27-28). Пожалуйста, нажмите здесь, чтобы просмотреть это видео.

Видео 2. трехмерное 360"вращение изображения на рисунке 4I, показывающее сосудистую систему и миграционный фронт ENCC в области caecum на E5.5 (HH27-28). Пожалуйста, нажмите здесь, чтобы просмотреть это видео.
Discussion
Метод внутривидовой трансплантации нервной трубки в сочетании с описанной здесь маркировкой кровеносных сосудов в полной мере использует легкость доступа птичьего эмбриона в яйцеклетку (по сравнению с другими позвоночными эмбрионами) для изучения совместного развития элемента вегетативной нервной системы (ENS) и сосудистой системы.
Для маркировки производных NCC, цыпленокGFP-цыпленоквнутривидовой прививки метод мы описываем имеет ряд преимуществ по сравнению с классическим перепела цыпленок химера метод, который был создан более 40 летназад 1-3. Во-первых, при свете FITC флуоресценция GFP чрезвычайно яркая, в той мере, в какой клетки GFP легко различимы в живых химерных эмбрионах. Это позволяет проверить успешное использование трансплантата в ово, в то времякак прививка перепелиного птенца требует, чтобы эмбрион был убит, обработан и иммунотелинирован с помощью ККПН, прежде чем успех трансплантата может быть установлен2. Во-вторых, экспрессия GFP втрансгенном цыпленок GFP является цитоплазмацией, поэтому она не только маркирует клеточные тела, но и позволяет визуализировать проекции пересаженныхклеток 22. Это позволяет наблюдать сложные нейронные сети с высоким разрешением (обратите внимание, что тонкие проекции лучше всего визуализировать, когда образец иммунотелен анти-GFP антитела). Поскольку маркировка ККПН ограничивается ядром перепелиных клеток, такие сети не выясвляются с использованием химер перепелиного цыпленка. В-третьих, внутривидовая прививка устраняет любые потенциальные видовые различия между клетками химерного эмбриона. Поскольку перепелиные эмбрионы имеют более короткий инкубационный период, чем цыпленок (19 дней против 21 дней), было высказано предположение, что перепелиные клетки имеют более высокий уровень распространения, чем клетки птенца, которые потенциально могут повлиять на развитие химерныхтканей 23. Интересно также, что было показано в растениях, что межвидовая прививка может производить обширные изменения в шаблонах метилирования ДНК в принимающей 24. В-четвертых,цыпленок GFP облегчает обратно трансплантации экспериментов для решения таких тем, как судьба NCC и клеточнойприверженности 25. В-пятых,трансгенный цыпленок GFP также полезен для многих других методов, включая сортировку FACS клеточной субпопуляции GFP, органотипической культуры органов, содержащих клетки GFP, генетические манипуляции привитой ткани GFP, через электропорацию плазмидовэкспрессии 26, и другие технологии визуализации, такие как оптическаяпроекционная томография 27.
Подход к трансплантации нервной трубки может быть изменен микрохирургически заменяя более короткие количества нервной трубки. С помощью небольших сегментов нервной трубки микрохирургия потенциально менее вредна для эмбриона и выживание может быть улучшено. Тем не менее, оборотная сторона трансплантации меньше нервной трубки является то, что число GFP NCC в принимающей будет сокращена. Пользователи могли бы попытаться достичь баланса между количеством нервной трубки пересажены, чтобы дать оптимальное выживание эмбрионов, и число GFP NCC в кишечнике хозяина достаточно, чтобы дать информативные результаты.
Для покраски сосудов, DiI имеет то преимущество, что его флуоресценция очень яркая и надежная. Кроме того, он имеет возможность диффузного во время фиксации обеспечения окрашивания лучших открытых капилляров. Так как это жизненно важный краситель, эмбрионы могут выжить процедуры инъекции и продолжать развиваться с окрашенных сосудистой системы (до 24 часов в наших руках, хотя окрашивание становится более пунктуатным с течением времени, см. Рисунок 3E). Таким образом, сочетаниепрививки цыпленка GFP с сосудистой живописью DiI совместимо с живой визуализацией. Помимо всех этих преимуществ, важно отметить, что сосудистые инъекции только маркируют люминесцентные сосуды и поэтому не идентифицируют неоткрытые капилляры, эндотелиальные кончики клеток или изолированные эндотелиальные клетки. Тем не менее, дальнейший прогресс в птичьем трансгенезе может предоставить новые способы обойти такие проблемы, о чем свидетельствуют эксперименты с использованием перепелиных эмбрионов Tg (tie1:H2B-eYFP) для изучения сосудистогоморфогенеза 28. Еще одним ограничением этого метода является то, что для эффективной маркировки сосудов в эмбрионах на E7.5 и за его пределами необходимо вводить большее количество красителя, что может сделать эксперименты дорогими. Тем не менее, модификация техники может включать в себя низкую стоимость кровеносных сосудов маркировки маркерчернил 14, хотя этот подход не был опробован в наших руках.
Критические шаги процедур включают процесс визуализации эмбриона путем введения чернил под бластодиском. Если мембрана, покрывающая желток, разрывается чернильной иглой на данном этапе, то выживаемость эмбриона серьезно скомпрометирована. Также важно, при подготовке донора нервной трубки, чтобы ткань не осталась на чрезмерно долгое время в панкреатине (считайте примерно 10 мин максимум). Длительное воздействие панкреатита повреждения тканей и нервной трубки, то трудно справиться, и он не будет включать хорошо в хозяина. Получение опыта метода инъекций DiI на эмбрионах дикого типа имеет важное значение перед введением химерных эмбрионов, так как только одна попытка инъекций, как правило, возможно для каждого эмбриона. Объем DiI и диаметр иглы являются критическими параметрами для каждого эмбриона и должны быть оценены по дикому типу, этапу соответствия элементов управления.
В заключение, наш метод двойной маркировки трансплантации нервной трубки и рисования сосудов DiI в живых эмбрионах цыпленка может быть использован для исследования взаимосвязей между NCC и сетями кровеносных сосудов во время органогенеза. Учитывая механизмы, ответственные за установление правильной иннервации цели и васкуляризации во время развития органов, до сих пор в значительной степени неизвестны, эта методология имеет потенциал для будущих открытий в этой области.
Disclosures
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Acknowledgments
Оплодотворенные куриные яйца GFP были поставлены профессором Хелен Санг, Институтом Рослина, и Эдинбургским университетом, Великобритания. Рослин Трансгенный куриный фонд финансируется Wellcome Trust и Научно-исследовательский совет по биотехнологии и биологическим наукам (BBSRC). Работа была частично профинансирована, и NT поддерживается, Грейт-Ормонд-стрит больницы Детская благотворительность, Лондон, Великобритания. Авторы благодарят Бена Джевана (Ben Jevans), Институт здоровья детей UCL, за помощь в подготовке эмбрионов к пересадке.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}