

Summary
Aqui relatamos rotulagem dupla de células de crista neural e vasos sanguíneos usando enxerto de intraespécie do tubo neuralGFP combinado com injeção intra-vascular diI. Esta técnica experimental nos permite visualizar e estudar simultaneamente o desenvolvimento do sistema nervoso derivado do NCC (entérico) e do sistema vascular, durante a organogênese.
Abstract
Todos os órgãos em desenvolvimento precisam estar conectados tanto ao sistema nervoso (para controle sensorial e motor) quanto ao sistema vascular (para troca de gás, fluido e fornecimento de nutrientes). Consequentemente, tanto os sistemas nervosos quanto vasculares se desenvolvem lado a lado e compartilham semelhanças marcantes em sua arquitetura ramificada. Aqui relatamos manipulações embrionárias que nos permitem estudar o desenvolvimento simultâneo do tecido nervoso derivado da crista neural (neste caso o sistema nervoso entérico) e o sistema vascular. Isso é conseguido através da geração de quimeras de frango através do transplante de segmentos discretos do tubo neural, e crista neural associada, combinada com injeção de DII vascular no mesmo embrião. Nosso método usa embriõesGFP de filhotes transgênicos para enxerto de subespécies, tornando a técnica de transplante mais poderosa do que o protocolo clássico de enxerto interespécie de codorna usado com grande efeito desde a década de 1970. O enxerto intraespécie dopintinho-filhotefacilita a imagem de células transplantadas e suas projeções em tecidos intactos, e elimina qualquer viés potencial no desenvolvimento celular ligado às diferenças de espécies. Este método aproveita ao máximo a facilidade de acesso do embrião aviário (comparado com outros embriões vertebrados) para estudar o co-desenvolvimento do sistema nervoso entérico e do sistema vascular.
Introduction
O embrião de frango é um organismo modelo inestimável na biologia do desenvolvimento de vertebrados, até porque seu desenvolvimento em ovo permite manipulações experimentais que são impossíveis de realizar em vertebrados que se desenvolvem no útero. Essa acessibilidade e facilidade de manipulações levou o embrião de pintinhos a desempenhar papéis-chave em muitas descobertas seminais no campo da biologia do desenvolvimento. Entre as técnicas mais poderosas tem sido o uso de embriões quiméricos de codorna para estudar o destino celular, método pioneiro pela professora Nicole Le Douarin na década de 19701-3. Em particular, quimeras de quail-chick têm sido especialmente úteis para marcar geneticamente e seguir populações de células de crista neural altamente migratórias (NCC) durante o desenvolvimento precoce. NCC é uma população multipotente de células migratórias, surgindo no ectoderme dorsal às margens do tubo neural, que dão origem a uma ampla gama de tipos celulares em todo o embrião vertebrado. Estes incluem estruturas craniofaciais (cartilagem, osso, músculos), neurônios e glia (nos sistemas nervosos sensorial e autônomo), melanócitos e subpopulação de células do sistema endócrino2,4,5. Um dos fatores mais importantes que influenciam o destino do NCC é sua localização inicial ao longo do eixo anterior-posterior do tubo neural. Por exemplo, o NCC entérico, que dá origem aos neurônios e à glia do sistema nervoso entérico (ENS), surge de duas subsu populações discretas: a primeira localizada na região do vagal (cérebro traseiro caudal) e a segunda na região sacral do tubo neural6-13. O enxerto inter ou intraespédicos das regiões correspondentes do tubo neural tem sido a técnica de escolha para rotular permanentemente essas células e, posteriormente, permitir o rastreamento, desde seu nascimento nas margens do tubo neural, até seus destinos finais dentro do trato digestivo6,7,10.
Outra manipulação embrionária mais fácil de realizar em filhotes, em comparação com outros modelos animais, é a rotulagem vital do sistema vascular. De fato, à medida que o embrião do filhote se desenvolve, ele fica em cima de uma rede vascular extraembrionária que circula oxigênio e nutrientes da gema. Esta rede vascular acessível, localizada na superfície da gema, pode ser usada como porta de entrada para rotular o sistema vascular em desenvolvimento do embrião durante a organogênese12,14-17. A injeção intravascular de vários corantes, como o corante lipofílico DiI, permite delinear/manchar todos os vasos luminosos da rede vascular nascente.
Como os órgãos em desenvolvimento precisam estar conectados tanto ao sistema nervoso (para controle sensorial e motor) quanto ao sistema vascular (para troca de gás, fluxo e fornecimento de nutrientes), as duas redes se desenvolvem lado a lado e compartilham semelhanças marcantes em sua arquiteturaramificada 18-20. Aqui relatamos manipulações embrionárias que nos permitem estudar o desenvolvimento simultâneo do ENS derivado do NCC, juntamente com o sistema vascular, durante a organogênese. Isso é conseguido através da geração de quimeras de frango através do transplante de segmentos discretos do tubo neural, incluindo a crista neural, combinada com a injeção vascular de DII. Como um avanço das quimeras de codorna-galinha, nosso método usa embriões transgênicos de filhotes de GFP para enxerto de subespécies, tornando a técnica de transplante mais poderosa, em termos de células de imagem e suas projeções, e eliminando qualquer viés potencial ligado às diferenças de espécies.
Protocol
1. Preparação de Micro-bisturi para ablações de tubos neurais
- Modele um micro-bisturi a partir de uma agulha de costura de aço comercialmente disponível.
- Primeiro achate a agulha em ambos os lados usando uma roda de moagem montada em um moedor de banco alimentado.
- Comece a moldar o bisturi, primeiro em uma pedra grosseira do Arkansas usando um movimento circular controlado, em direções alternativas, em ambos os lados da agulha.
- Continue os mesmos movimentos de afiação em uma pedra arkansas de grau extra fino para moldar um micro-bisturi ultra fino, com uma borda de corte bem definida(Figura 1A, B).
NOTA: Alternativas ao micro-bisturi podem ser agulhas eletroliticamente afiadas, agulhas de tungstênio comercialmente disponíveis ou agulhas de vidro puxadas.
2. Incubar ovos selvagens e gfp para o estágio desejado
- Armazene ovos de galinha fertilizados e ovos de galinha GFP transgênicos em uma incubadora resfriada a 14 - 15 °C antes da incubação à medida que o desenvolvimento é interrompido a esta temperatura. Guarde ovos por alguns dias, até uma semana.
- Para iniciar o desenvolvimento, coloque ovos de tipo selvagem e GFP em uma bandeja horizontalmente e incubar simultaneamente em umidificado (58 - 60%) incubadora a 37,5 ºC, de modo que os embriões estão em estágios correspondentes para enxerto de tubo neural.
- Para obter embriões no estágio de desenvolvimento de 10 a 12 somites para a realização do enxerto de tubo neural vagal, incubar ovos por 1,5 dias (33 - 38 horas) e embriões de estágio de acordo com as tabelas de desenvolvimento de Hamburger e Hamilton21.
3. Prepare ovos para janelas e enxertos
- Mova um ovo de cada vez para um suporte de ovos feito sob medida para janelas. Faça um pequeno furo na casca do ovo batendo repetidamente, com uma tesoura reta, na superfície superior da extremidade pontiaguda do ovo.
- Remova 2 - 3 ml de albumina do ovo com uma agulha hipodérmica de 181/2 G e 5 ml de seringa. A remoção da albumina abaixa a gema dentro do ovo e facilita a janela subsequente sem causar qualquer dano ao embrião.
- Descarte a albumina. Sele o orifício com uma pequena tira de fita clara cortada em tamanho com uma tesoura fina.
- Usando uma tesoura curva, bata outro orifício na superfície superior da casca do ovo. Insira a ponta da tesoura no orifício e, mantendo a tesoura paralela ao banco, trabalhe em um movimento circular para cortar uma janela de ~2 cm de diâmetro em cima da concha.
- Mantenha a tesoura em posição estacionária e gire o ovo. Descarte o disco removido da casca de ovo. Em E1.5, o embrião é reconhecível como um disco amarelo mais escuro em cima da gema.
- Remova os detritos da casca que caíram dentro do ovo usando pinças. Descarte quaisquer ovos não fertilizados (identificados por uma pequena mancha branca em cima da gema amarela clara).
4. Prepare o embrião hospedeiro para receber tecido enxertado
- Ajuste o microscópio estéreo ao nível dos olhos e otimize a orientação da fonte de luz gooseneck para iluminar adequadamente o embrião sem causar reflexos.
- Para visualizar o embrião propriamente dito,injete uma pequena quantidade de tinta indiana sob o centro do disco amarelo mais escuro, usando um tubo bucal e uma micro-pipeta de vidro puxado(Figura 1C, Oii).
- Prepare a tinta 50:50 com PBS contendo Penicilina/Estreptomicina a 100 μg/ml de concentração final. Insira a micro-pipeta através da membrana da gema fora do perímetro da blastoderm e, em seguida, angeta cuidadosamente sua ponta diretamente abaixo do embrião.
- Entregue tinta debaixo do embrião soprando no tubo bucal. Se a pipetação bucal não for permitida, use uma seringa de 1 ml em vez disso. Tenha cuidado para não introduzir bolhas de ar sob o embrião, que pode levar à contaminação, em seguida, remova cuidadosamente a micro-pipeta de vidro. Este é um passo delicado que pode levar à morte do embrião se não for feito com precisão.
- Enseia o embrião por referência a Hamburger e Hamilton21 e grava o palco em um livro de laboratório.
- Usando um micro-bisturi feito sob medida (ou uma agulha de tungstênio fino) montado em um suporte de agulha, faça um corte muito pequeno na membrana vitelline, ao lado da área onde a microcirurgia será realizada.
- Aplique cuidadosamente 2 - 3 gotas de PBS sobre a ruptura da membrana (usando uma micro-pipeta de vidro e o tubo bucal) para criar espaço entre o embrião e a membrana. Corte uma janela maior na membrana para expor toda a região onde ocorrerá a microcirurgia.
- Remova a região de interesse do tubo neural usando o micro-bisturi, começando com incisões transversais rostral e caudal em todo o tubo neural dorsal (ao nível de somite 1 a 7 no vídeo).
- Corte bilateralmente entre o tubo neural e os somitas para separar o tubo neural dos tecidos circundantes, sem danificar os somites.
- Muito gentilmente separe o tubo neural do notochord subjacente, que deve permanecer intacto. Note que a excisão do tubo neural bem sucedida deixará todos os tecidos circundantes perfeitamente intactos(Figura 2).
- Remova o tubo neural excisado aspirando-o em uma micro-pipeta de vidro, em seguida, descarte.
- Regisso nível de ablação de tubo neural em um livro de laboratório. O embrião hospedeiro está pronto para receber o tubo neural do doador.
5. Prepare o tecido do enxerto de doador
- Selecione um embrião GFP com janelas e partidas de palco, visualizando sob um microscópio estéreo fluorescente com filtro FITC. A fluorescência GFP torna muito fácil visualizar os somites e encenar o embrião.
- Uma vez identificado um embrião compatível com o estágio, remova o embrião do ovo fazendo 4 incisões, com uma tesoura de mola Pascheff-Wolff(Figura 1C, l) em uma forma de retângulo ao redor do embrião e, em seguida, pegá-lo suavemente com uma colher de embrião.
- Coloque o embrião em um vidro de relógio quadrado com uma base de polímero sylgard. Agite suavemente o embrião com Dumont #5 pinças para remover qualquer gema presa. Remova a membrana vitelina e fixe o embrião na base do polímero usando pinos de minutien inoxidáveis(Figura 1C).
- Usando a Tesoura de Mola, faça 4 incisões em uma forma retangular ao redor do tubo neural e somitas circundantes, na mesma região que foi removida do embrião hospedeiro.
- Usando uma pipeta de transferência de plástico, transfira o tubo neural e os tecidos de somite do embrião GFP doador para um vidro de relógio contendo 0,2% de pancreatina em Pen/Strep PBS.
- Permita que a digestão enzimática prossiga por 10 minutos no RT para ajudar a separar os tecidos. Após a incubação em enzima, use pinos de minutien inoxidáveis montados em uma alça para separar manualmente o tubo neural de todos os tecidos adjacentes.
- Usando uma micro-pipeta de vidro, transfira o tubo neural dissociado para outro vidro de relógio contendo DMEM + 10% de soro (por exemplo,cabra, cavalo ou bezerro fetal) no gelo, para enxaguar o excesso de pancreatina e parar a digestão enzimática. Após 5 min, o tubo neural dissecado está pronto para ser enxertado ortotopicamente no hospedeiro filhote(Figura 2 e S1).
6. Enxertar o tecido
- Usando uma micro-pipeta de vidro, transfira cuidadosamente o tubo neural dissecado do vidro do relógio para o embrião hospedeiro. Posicione o tubo neural na orientação anterior-posterior correta e empurre suavemente a explanta adjacente à região excisada do hospedeiro do filhote usando o micro-bisturi. Deixe um pequeno fragmento de ectoderme ligado, ou cortando um pequeno corte na superfície dorsal para identificar a orientação do tubo neural.
- Se necessário, use o micro-bisturi para aparar a explanta ao tamanho exato da região excisada.
- Guie suavemente o tubo neural para a região ablada e posicione-o de tal forma que o lado dorsal seja corretamente orientado. Use uma micro-pipeta de vidro, montada em um tubo bucal, para remover PBS e/ou fluido ao redor do enxerto. Isso ajuda o doador e os tecidos de acolhimento a aderir e o enxerto a se estabelecer.
- Sele toda a janela com fita clara de 24 mm de largura para evitar desidratação e contaminação.
- Rotule o embrião quimérico marcando com um lápis na casca de ovo e grave seu número no livro de laboratório. Devolva o ovo à incubadora para maior desenvolvimento.
7. Injete DII em vasos sanguíneos de embrião hospedeiro
- No ponto de tempo experimental desejado (aqui, 3 - 10 dias depois), recupere o embrião quimérico da incubadora e remova a fita clara usando uma tesoura reta para ter acesso ao embrião dentro do ovo.
- Se necessário, amplie a janela da concha usando a tesoura. Tenha cuidado para não danificar a membrana corioallantónica se estiver presa à casca, o que resultaria em hemorragia e colocaria em risco a rotulagem dos vasos sanguíneos.
- Escolha uma veia acessível na gema certificando-se de que o fluxo sanguíneo seja direcionado para o embrião. Escolha um ponto de ramificação de uma das veias vitelline(Figura 3B, C).
NOTA: Em E6.5 - E7.5, a membrana corioallantórica pode precisar ser suavemente movida de lado com pinças para acessar as veias da gema. Após o E8.5, a única opção é injetar em uma das veias de membrana corioallantóica, já que, por esta fase, a membrana corioallantóica cobre totalmente o embrião. - Remova a membrana vitelline acima do ponto de injeção escolhido usando duas pinças Dumont #5 rasgando em direções opostas.
- Quebre uma agulha de vidro puxada usando um #5 Dumont e ajuste seu diâmetro para o tamanho aproximado da veia antes de carregar com CellTracker CM-DiI. Faça a solução de estoque DiI a 40 μg/μl em DMSO e armazene a -20 °C. Prepare a solução de trabalho em 0,3 M de sacarose/PBS a uma concentração de 4 μg/μl.
- Aspire entre 5 - 10 μl de DII em 0,3 M de sacarose/PBS na agulha usando sucção com um tubo bucal. Embriões mais velhos podem exigir até 25 μl ou mais. A partir de E8.5, os embriões têm veias maiores e mais musculares, que podem precisar ser mantidas em posição com um #5 Dumont antes de esfaquear com a agulha de vidro carregada de DII.
- Insira rapidamente a agulha na veia e sopre constantemente com o tubo bucal para permitir que o DII se junte ao fluxo sanguíneo lentamente sem formar um coágulo. Alternativamente, use um injetor de pressão para a entrega de DII.
8. Colher embriões para secção ou exame de integral
- Para reter o máximo de DII dentro do embrião possível, colde o embrião imediatamente após a injeção, recolhendo-o em uma colher perfurada e cortando os vasos sanguíneos e tecidos conjuntivos com um par de tesouras retas, para libertar o embrião da gema.
- Remova quaisquer membranas soltas e disseque os órgãos de interesse (ou seja,os pulmões e o trato digestivo neste tutorial), tomando muito cuidado para não comprimir o tecido, o que cria difusão do DII. Fixar imediatamente os tecidos por imersão em 4% pfa por 1 - 2 horas na RT.
- Enxágüe o tecido por 5 min em PBS, depois 15 min em PBS contendo 5 μg/ml DAPI. Monte as amostras em um slide de microscópio ponte para exame de montagem inteiro ou incorpore-as para seção criobiósa.
Representative Results
A Figura 1 mostra instrumentos típicos necessários para realizar isolamento microcirúrgico e transplante do tubo neural. A Figura 2 mostra o procedimento de transplante. Após transplante, os embriões são examinados para o sucesso do transplante. Isso envolve examinar o embrião sob um microscópio de fluorescência estéreo, tipicamente na manhã seguinte à microcirurgia, para a presença de NCC derivado do enxerto (GFP+). Se o transplante foi um sucesso, então o GFP+ NCC pode ser observado nas proximidades do tubo neural e nas vias migratórias precoces que levam ao foregut. Se o procedimento não tiver sido bem sucedido, o GFP+ NCC não será observado fora do tubo neural, ou se eles estiverem presentes no hospedeiro, eles podem estar em menor número. Estes embriões mal sucedidos são descartados. Normalmente, 5-8 transplantes de tubo neural são realizados em um dia, e destes 80% são bem sucedidos. As razões para o transplante de tubo neural mal sucedido incluem a morte do embrião devido a danos teciduais incorridos durante a microcirurgia, ou falha do tubo neural para se integrar ao embrião hospedeiro. Este último pode resultar da má colocação do tubo neural dentro do hospedeiro ou de um tubo neural de má qualidade devido à má técnica de dissecção ou da exposição excessiva à enzima dissociação. A etapa inicial de triagem, bem como exames posteriores semelhantes para células GFP+, é útil, pois significa que tempo e recursos não são desperdiçados pela realização de experimentos em embriões que não têm NCC rotulado por GFP dentro do intestino.
A Figura 3 mostra o procedimento para injeção dii dos vasos sanguíneos. A eficiência/sucesso da técnica de injeção dii depende: primeiro, cortar a agulha de injeção para o diâmetro ideal para a veia alvo, segundo um gesto preciso ao inserir a agulha na veia (para não perfurar pelo outro lado), e terceiro evitando que a agulha seja plugada durante a injeção soprando a uma taxa constante. Se algum desses três parâmetros for feito incorretamente, o embrião sangrará ou precisará de várias horas para se recuperar antes que uma segunda tentativa seja feita, pois a hemorragia torna quase impossível reinjetar imediatamente. Embriões bem sucedidos devem ser selecionados imediatamente visualizando sob um microscópio de estereofluorescência e devem ser dissecados rapidamente. Em embriões bem sucedidos, os vasos sanguíneos rotulados por DII estão presentes em todo o embrião (Figura 3C,D), incluindo leitos capilares(Figura 3D).
Após a colheita de embriões e exame de seções teciduais ou tratos gastrointestinais de toda adoeção, resultados típicos revelam que o GFP+ NCC dentro do ENS primitivo e a estrutura fina das redes de vasos sanguíneos rotulados por DiI(Figura 4) Preparações integrais podem ser examinadas por meio de microscopia confocal pela qual as pilhas de imagem produzem reconstruções tridimensionais (3D) mostrando as inter-relações entre as projeções finas das células GFP+ ENS e o sistema vascular manchado de DI(Figura 4 A-C; G-I; Vídeos 1 e 2).

Figura 1. Instrumentos de microcirurgia recomendados. (A) micro-bisturi em forma de agulha de costura. (B) pedra fina do Arkansas para moldar um micro-bisturi. (C) a) tesoura reta, b) tesoura curva, c) seringa de 5 ml com agulha hipodérmica de 181/2 G, d) pipeta plástica, e) suporte de ovo feito sob medida, f) tinta preta, g) vidro de relógio quadrado, h) vidro de relógio quadrado com base sylgard preta, i) micro-bisturi no porta-agulhas, j) pinos minutien, k) minutien ou agulha de tungstênio no porta-agulhas, l) Tesoura de mola Pascheff-Wolff, m) Dumont #5 pinças, n) colher perfurada, oi) agulha de transferência de fogo curta, oii) longo fogo puxado na agulha, p) Clique aqui para ver uma versão maior desta figura.

Figura 2. Transplante de tubo neural intraespécie. As imagens do tubo neural chick/GFP foram modificadas de Delalande et al. 12. A vascularização não é necessária para a colonização intestinal por células de crista neural entárica. Clique aqui para ver uma versão maior desta figura.

Figura 3. Injeção intravenosa de DiI. (A) Instrumentos recomendados: a) Queda do CellTracker CM-DiI no parafilme, b) agulha de injeção de vidro puxada, c) tubo bucal. (B) Diagrama esquemático da injeção dii intravenosa em embrião quimérico E4. (C) em ovo DiI injeção intravenosa mostrando agulha de vidro fina contendo DI Inserida na veia (seta). (D) Embrião quimérico E4 pós injeção dii (vermelho) com tubo neural GFP+ (seta). (E) DiI manchou rede de vasos sanguíneos finos em um embrião vivo, 24 horas após a injeção. Br: cérebro; H: coração; LB: broto de membro; A: allantois. As imagens em (C) e (D) foram modificadas de Delalande et al. 12 A vascularização não é necessária para a colonização intestinal por células de crista neural entônica. Clique aqui para ver uma versão maior desta figura.

Figura 4: Resulta representativo no estômago e caecum de um embrião de pintinho E5.5. (A-C) 3D reconstrução dimensional (3D) de uma pilha de imagens confocal na região do estômago mostrando (D) as células de crista neural enteric GFP+ (ENCC) (E) o sistema vascular manchado de DiI e (F) uma imagem mesclada de ambas as redes D-F Seções histológicas ao nível do estômago mostrando (G) o GFP+ ENCC (H) o sistema vascular manchado do DII e (I) uma imagem mesclada de ambas as redes. Os núcleos estão manchados com DAPI (ciano). (G-H) reconstrução 3D de uma pilha de imagens confocal na região do caecum mostrando (A) a frente de migração GFP+ ENCC em verde, (B) o sistema vascular manchado de DiI em vermelho, e (C) uma imagem mesclada de ambas as redes. As imagens (A-F) foram modificadas de Delalande et al. 12 A vascularização não é necessária para a colonização intestinal por células de crista neural entônica. Clique aqui para ver uma versão maior desta figura.

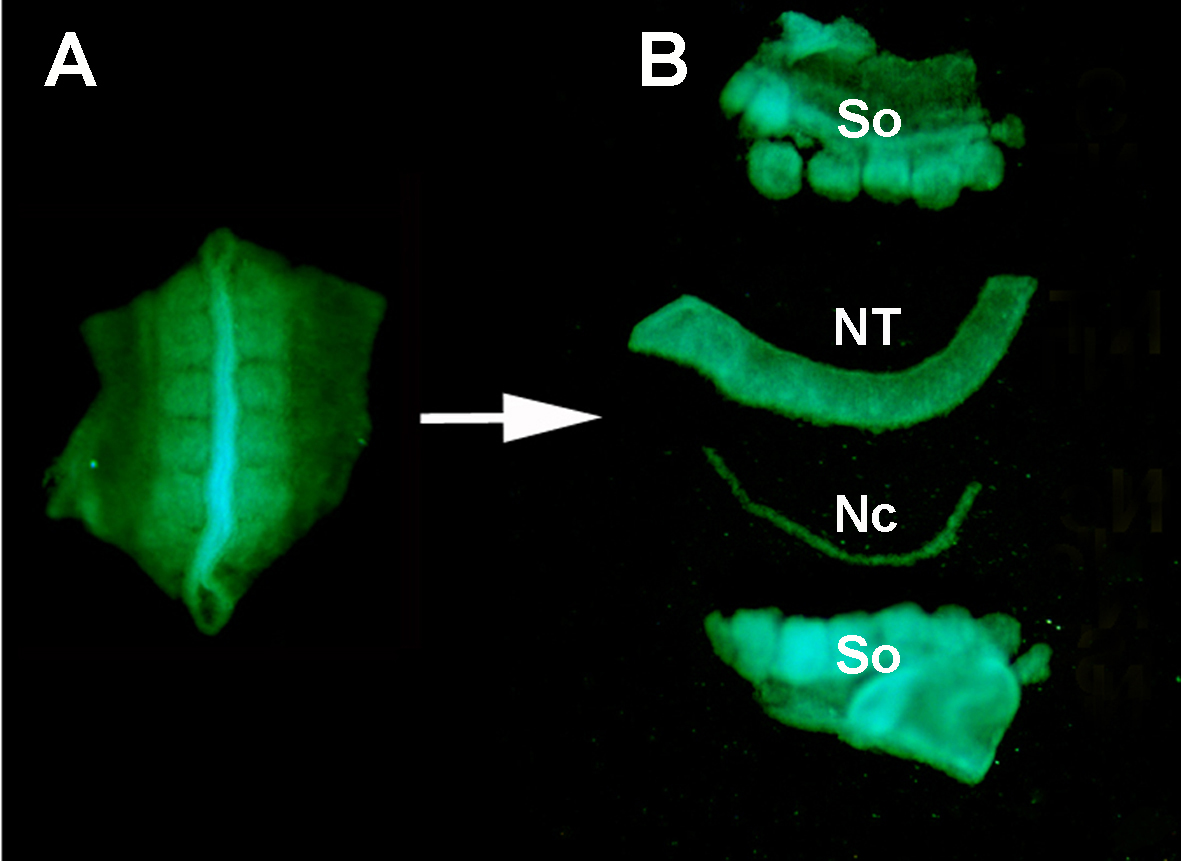
Figura S1. Isolamento de um tubo neural GFP+ doador dos tecidos circundantes por digestão enzimática e microdiscção. (A) Tubo neural GFP+ e somitas adjacentes dissecados do embrião doador. (B) Tubo neural isolado após digestão de pancreatina e micro-dissecção usando pinos minutien inoxidáveis. Então: somitas; NT: tubo neural; Nc: Notochord.

Vídeo 1. rotação 360° tridimensional da imagem na Figura 4C, mostrando o sistema vascular e o ENCC no estômago em E5.5 (HH27-28). Clique aqui para ver este vídeo.

Vídeo 2. rotação 360° tridimensional da imagem na Figura 4I, mostrando o sistema vascular e a frente de migração ENCC na região do caecum em E5.5 (HH27-28). Clique aqui para ver este vídeo.
Discussion
O método de enxerto de tubo neural intraespécie, combinado com a rotulagem dos vasos sanguíneos aqui descrito, aproveita ao máximo a facilidade de acesso do embrião aviário dentro do óvulo (comparado com outros embriões vertebrados) para estudar o co-desenvolvimento de um elemento do sistema nervoso autônomo (o ENS) e do sistema vascular.
Para rotular derivados NCC, o método de enxerto de subespécie de pintinhosGFP-chickque descrevemos tem uma série de vantagens sobre o método clássico de quimera de codorna que foi estabelecido há mais de 40 anos1-3. Primeiro, sob a luz FITC, a fluorescência GFP é extremamente brilhante, na medida em que as células GFP+ são facilmente discerníveis em embriões quiméricos vivos. Isso permite que o sucesso do enxerto seja verificado no ovo,enquanto o enxerto de codorna exige que o embrião seja morto, processado e imunossuído usando QCPN, antes que o sucesso do enxerto possa ser apurado2. Em segundo lugar, a expressão GFP no filhote transgênicoGFP é citoplasmática, portanto não apenas rotula corpos celulares, mas também permite que as projeções das células transplantadas sejam visualizadas22. Isso permite que redes neuronais intrincadas sejam observadas em alta resolução (observe que as projeções finas são melhor visualizadas quando a amostra é imunossuada com anticorpo anti-GFP). Como a rotulagem de QCPN é restrita ao núcleo da célula de codorna, tais redes não são reveladas usando quimeras de codorna. Em terceiro lugar, o enxerto de intraespécie elimina quaisquer diferenças potenciais de espécies entre as células dentro do embrião quimérico. Uma vez que os embriões de codorna têm um período de incubação menor do que o filhote (19 dias versus 21 dias) tem sido sugerido que as células de codorna têm uma taxa de proliferação maior do que as células-pintinhos, o que poderia afetar potencialmente o desenvolvimento dos tecidos quiméricos23. Curiosamente, também tem sido demonstrado em plantas que o enxerto interespécie poderia produzir alterações extensas nos padrões de metilação de DNA no hospedeiro 24. Em quarto lugar, oChick GFP facilita experimentos de transplante suplementar para abordar temas como destino NCC e comprometimento celular25. Em quinto lugar, o filhote transgênicoGFP também é útil para muitas outras técnicas, incluindo a triagem FACS de subpopulações celulares GFP+, cultura organotípica de órgãos contendo células GFP+, manipulação genética do tecido enxertado GFP+ através de eletroporação de plasmídeos de expressão26,e outras tecnologias de imagem, como a tomografia de projeção óptica27.
A abordagem de transplante de tubo neural pode ser modificada substituindo microsurgicamente quantidades mais curtas de tubo neural. Usando segmentos menores do tubo neural, a microcirurgia é potencialmente menos prejudicial ao embrião e a sobrevivência pode ser melhorada. No entanto, a desvantagem de transplantar menos tubo neural é que o número de GFP+ NCC no hospedeiro será reduzido. Os usuários poderiam tentar obter um equilíbrio entre a quantidade de tubo neural transplantado para dar a melhor sobrevida de embriões, e o número de GFP+ NCC dentro do intestino hospedeiro suficiente para dar resultados informativos.
Para a pintura de vasos, DiI tem a vantagem de que sua fluorescência é muito brilhante e robusta. Além disso, tem a capacidade de difundir durante a fixação garantindo a coloração dos melhores capilares abertos. Por ser um corante vital, os embriões podem sobreviver ao procedimento de injeção e continuar se desenvolvendo com um sistema vascular manchado (até 24 horas em nossas mãos, embora a coloração se torne mais pontual ao longo do tempo, ver Figura 3E). A combinação de enxertoGFP com pintura vascular dii é, portanto, compatível com imagens vivas. Além de todas essas vantagens, é importante notar que a injeção vascular só rotula vasos luminosos e, portanto, não identifica capilares não abertos, células de ponta endotelial ou células endoteliais isoladas. No entanto, novos progressos na transgênese aviária poderiam fornecer novas maneiras de contornar tais questões, como exemplificado por experimentos usando embriões de codorna Tg (tie1:H2B-eYFP) para estudar morfognese vascular28. Outra limitação dessa técnica é que, para uma rotulagem eficaz de vasos em embriões em E7.5 e além, maiores quantidades de corante precisam ser injetadas, o que pode tornar os experimentos caros. No entanto, uma modificação da técnica pode incluir rotulagem de vaso sanguíneo de baixo custo com tinta de destaque14,embora essa abordagem não tenha sido testada em nossas mãos.
As etapas críticas dos procedimentos incluem o processo de visualização do embrião injetando tinta sob o blastodisc. Se a membrana que cobre a gema for rasgada pela agulha cheia de tinta nesta fase, então a sobrevivência do embrião será severamente comprometida. Além disso, é importante, ao preparar um tubo neural doador, que o tecido não seja deixado por um tempo excessivamente longo em pancreatina (considere aproximadamente 10 min como um máximo). A exposição prolongada à pancreatina danifica o tecido e o tubo neural é então difícil de manusear e não incorporará bem no hospedeiro. Ganhar experiência da técnica de injeção dii em embriões do tipo selvagem é essencial antes de injetar embriões quiméricos, pois apenas uma tentativa de injeção é geralmente possível para cada embrião. O volume de DI E o diâmetro da agulha são parâmetros críticos para cada embrião e devem ser avaliados em tipo selvagem, controles combinados de estágio.
Em conclusão, nosso método de rotulagem dupla de transplante de tubos neurais e pintura de vasos dii em embriões de filhotes vivos pode ser usado para investigar as inter-relações entre ncc e redes de vasos sanguíneos durante a organogênese. Considerando que os mecanismos responsáveis por estabelecer a inervação e vascularização corretas durante o desenvolvimento dos órgãos ainda são amplamente desconhecidos, essa metodologia tem potencial para futuras descobertas nesse campo.
Disclosures
Os autores declaram que não têm interesses financeiros concorrentes.
Acknowledgments
Os ovos de galinha GFP fertilizados foram fornecidos pela Prof. Helen Sang, o Instituto Roslin e a Universidade de Edimburgo, Reino Unido. O Roslin Transgenic Chicken Facility é financiado pelo Wellcome Trust e pelo Conselho de Pesquisa em Biotecnologia e Ciências Biológicas (BBSRC). O trabalho foi parcialmente financiado, e nt apoiado, por Great Ormond Street Hospital Children's Charity, Londres, Reino Unido. Os autores agradecem a Ben Jevans, do Instituto de Saúde infantil da UCL, pela ajuda na preparação de embriões para enxerto.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}