

Summary
Qui segnaliamo la doppia etichettatura delle cellule della cresta neurale e dei vasi sanguigni utilizzando l'innesto intraspecie del tubo neuraleGFP del pulcino combinato con l'iniezione di DiI intra-vascolare. Questa tecnica sperimentale ci permette di visualizzare e studiare contemporaneamente lo sviluppo del sistema nervoso derivato dall'NCC (enterico) e del sistema vascolare, durante l'organogenesi.
Abstract
Tutti gli organi in via di sviluppo devono essere collegati sia al sistema nervoso (per il controllo sensoriale e motorio) che al sistema vascolare (per lo scambio di gas, l'apporto di liquidi e nutrienti). Di conseguenza sia il sistema nervoso che quello vascolare si sviluppano uno accanto all'altro e condividono sorprendenti somiglianze nella loro architettura ramificata. Qui segnaliamo manipolazioni embrionali che ci permettono di studiare lo sviluppo simultaneo del tessuto nervoso derivato dalla cresta neurale (in questo caso il sistema nervoso enterico) e del sistema vascolare. Ciò si ottiene generando chimere di pollo attraverso il trapianto di segmenti discreti del tubo neurale e della cresta neurale associata, combinata con l'iniezione vascolare di DiI nello stesso embrione. Il nostro metodo utilizza embrioni GFP dipulcini transgenici per l'innesto intraspecie, rendendo la tecnica di trapianto più potente del classico protocollo di innesto di interspecie quaglia-pulcino utilizzato con grande effetto dagli anni '70. L'innestointraspecie di pulcini GFP -pulcini facilita l'imaging delle cellule trapiantate e le loro proiezioni nei tessuti intatti ed elimina qualsiasi potenziale pregiudizio nello sviluppo cellulare legato alle differenze delle specie. Questo metodo sfrutta appieno la facilità di accesso dell'embrione aviario (rispetto ad altri embrioni di vertebrati) per studiare il co-sviluppo del sistema nervoso enterico e del sistema vascolare.
Introduction
L'embrione di pollo è un organismo modello inestimabile nella biologia dello sviluppo dei vertebrati, non da ultimo perché il suo sviluppo in ovo consente manipolazioni sperimentali che sono altrimenti impossibili da eseguire nei vertebrati che si sviluppano in utero. Questa accessibilità e facilità di manipolazione ha portato l'embrione pulcino a svolgere ruoli chiave in molte scoperte seminali nel campo della biologia dello sviluppo. Tra le tecniche più potenti c'è stato l'uso di embrioni chimerici di quaglia-pulcini per studiare il destino cellulare, un metodo introdotto dalla professoressa Nicole Le Douarin neglianni '70 1-3. In particolare, le chimere quaglia-pulcino sono state particolarmente utili per marcare e seguire geneticamente le popolazioni di cellule di cresta neurale altamente migratorie (NCC) durante lo sviluppo precoce. Gli NCC sono una popolazione multipotente di cellule migratorie, che si presenta nell'ectoderma dorsale ai margini del tubo neurale, che danno origine a una vasta gamma di tipi di cellule in tutto l'embrione vertebrato. Questi includono strutture craniofacciali (cartilagine, ossa, muscoli), neuroni e glia (nel sistema nervoso sensoriale e autonomo), melanociti e una sottopopolazione di cellule del sistema endocrino2,4,5. Uno dei fattori più importanti che influenzano il destino degli NCC è la loro posizione iniziale lungo l'asse anteriore-posteriore del tubo neurale. Ad esempio, gli NCC enterici, che danno origine ai neuroni e agli glia del sistema nervoso enterico (ENS), derivano da due sotto-popolazioni discrete: la prima situata nella regione vagala (hindbrain caudale) e la seconda nella regione sacrale del tubo neurale6-13. L'innesto inter-specie o intra-specie delle corrispondenti regioni del tubo neurale sono state le tecniche di scelta per etichettare permanentemente queste cellule e successivamente consentire il tracciamento, dalla loro nascita ai margini del tubo neurale, alle loro destinazioni finali all'interno del tratto digestivo6,7,10.
Un'altra manipolazione embrionale più facile da eseguire nel pulcino, rispetto ad altri modelli animali, è l'etichettatura vitale del sistema vascolare. Infatti, man mano che l'embrione del pulcino si sviluppa, si trova sopra una rete vascolare extra-embrionale che fa circolare ossigeno e sostanze nutritive dal tuorlo. Questa rete vascolare accessibile, situata sulla superficie del tuorlo, può essere utilizzata come porta d'accesso per etichettare il sistema vascolare in via di sviluppo dell'embrione durante l'organogenesi12,14-17. L'iniezione intravascolare di vari coloranti, come il colorante lipofilo DiI, consente di delineare/macchiare tutti i vasi luminati della nascente rete vascolare.
Poiché lo sviluppo di organi deve essere collegato sia al sistema nervoso (per il controllo sensoriale e motorio) che al sistema vascolare (per lo scambio di gas, l'apporto di liquidi e nutrienti), le due reti si sviluppano l'una accanto all'altra e condividono sorprendenti somiglianze nella loro architetturaramificata 18-20. Qui denunciamo manipolazioni embrionali che ci permettono di studiare lo sviluppo simultaneo dell'ENS derivato da NCC, insieme al sistema vascolare, durante l'organogenesi. Ciò si ottiene generando chimere di pollo attraverso il trapianto di segmenti discreti del tubo neurale, compresa la cresta neurale, combinato con l'iniezione vascolare di DiI. Come avanzamento delle chimere di quaglia-pollo, il nostro metodo utilizza embrioni transgenici di pulcini GFP per l'innesto intraspecie, rendendo la tecnica del trapianto più potente, in termini di cellule di imaging e delle loro proiezioni, ed eliminando qualsiasi potenziale pregiudizio legato alle differenze di specie.
Protocol
1. Preparazione del micro-bisturi per le ablazioni del tubo neurale
- Modella un micro-bisturi da un ago da cucito in acciaio disponibile in commercio.
- Per prima cosa appiattire l'ago su entrambi i lati utilizzando una mola montata su una smerigliatrice da banco motorizzazione.
- Inizia a modellare il bisturi, prima su una pietra grossolana dell'Arkansas usando un movimento circolare controllato, in direzioni alternative, su entrambi i lati dell'ago.
- Continuare gli stessi movimenti di affilata su una pietra arkansas di grado extra fine per modellare un micro-bisturi ultra fine, con un tagliente ben definito(Figura 1A, B).
NOTA: Le alternative al micro-bisturi potrebbero essere aghi elettroliticamente affilati, aghi di tungsteno disponibili in commercio o aghi di vetro tirato.
2. Incubare il tipo selvatico e le uova GFP allo stadio desiderato
- Conservare le uova di gallina fecondate e le uova di gallina GFP transgeniche in un'incubatrice raffreddata a 14 - 15 °C prima dell'incubazione poiché lo sviluppo viene interrotto a questa temperatura. Conservare le uova per alcuni giorni, fino a una settimana.
- Per iniziare lo sviluppo, posizionare il tipo selvatico e le uova GFP su un vassoio orizzontalmente e incubare contemporaneamente in un umidificato (58 - 60%) incubatore a 37,5 ºC, in modo che gli embrioni siano in fasi di corrispondenza per l'innesto di tubi neurali.
- Per ottenere embrioni allo stadio di sviluppo di 10 - 12 somite per effettuare l'innesto del tubo neurale vagale, incubare le uova per 1,5 giorni (33 - 38 ore) ed embrioni di stadio secondo le tabelle di sviluppo di Hamburger e Hamilton21.
3. Preparare le uova per la finestra e l'innesto
- Sposta un uovo alla volta su un portaovo su misura per la finestra. Fai un piccolo buco nel guscio d'uovo toccando ripetutamente, con forbici dritte, sulla superficie superiore dell'estremità appuntita dell'uovo.
- Rimuovere 2 - 3 ml di albumina dall'uovo con un ago ipodermico da 181/2 G e una siringa da 5 ml. La rimozione dell'albumina abbassa il tuorlo all'interno dell'uovo e facilita la successiva finestra senza causare alcun danno all'embrione.
- Scartare l'albumina. Sigillare il foro con una piccola striscia di nastro chiaro tagliato a misura con forbici fini.
- Usando forbici curve, tocca un altro foro nella superficie superiore del guscio d'uovo. Inserire la punta delle forbici nel foro e, mantenendo le forbici parallele alla panca, lavorare con un movimento circolare per tagliare una finestra di ~ 2 cm di diametro sopra il guscio.
- Tenere le forbici in posizione stazionaria e ruotare l'uovo. Scartare il disco rimosso del guscio d'uovo. A E1.5, l'embrione è riconoscibile come un disco giallo più scuro sopra il tuorlo.
- Rimuovere eventuali detriti di guscio caduti all'interno dell'uovo usando una pinzetta. Scartare eventuali uova non fecondate (identificate da una piccola macchia bianca sulla parte superiore sul tuorlo giallo altrimenti chiaro).
4. Preparare l'embrione ospite per ricevere il tessuto innestato
- Regolare lo stereo-microscopio a livello degli occhi e ottimizzare l'orientamento della sorgente luminosa a collo d'oca per illuminare adeguatamente l'embrione senza causare riflessi.
- Per visualizzare l'embrione vero e proprio, iniettare una piccola quantità di inchiostro indiano sotto il centro del disco giallo più scuro, utilizzando un tubo della bocca e una micro-pipetta di vetro tirato (Figura 1C, Oii).
- Preparare l'inchiostro 50:50 con PBS contenente penicillina/streptomicina a 100 μg/ml di concentrazione finale. Inserire la micro-pipetta attraverso la membrana del tuorlo all'esterno del perimetro del blastoderma, quindi inclinare attentamente la punta direttamente sotto l'embrione.
- Fornire inchiostro sotto l'embrione soffiando sul tubo della bocca. Se la pipettazione della bocca non è consentita, utilizzare invece una siringa da 1 ml. Fare attenzione a non introdurre bolle d'aria sotto l'embrione, che possono portare alla contaminazione, quindi rimuovere con cura la micro-pipetta di vetro. Questo è un passo delicato che può portare alla morte dell'embrione se non fatto con precisione.
- Scenae l'embrione con riferimento ad Hamburger e Hamilton21 e registrare la tappa di un libro di laboratorio.
- Utilizzando un micro-bisturi su misura (o un ago di tungsteno fine) montato su un portaaghi, fare un gash molto piccolo nella membrana vitellina, vicino all'area in cui verrà eseguita la microchirururghi.
- Applicare con cura 2 - 3 gocce di PBS sulla lacrima della membrana (utilizzando una micro-pipetta di vetro e il tubo della bocca) per creare spazio tra l'embrione e la membrana. Tagliare una finestra più grande nella membrana per esporre l'intera regione in cui avrà luogo la micro-chirurgia.
- Rimuovere la regione di interesse del tubo neurale utilizzando il micro-bisturi, a partire dalle incisioni trasversali rostrali e caudali su tutto il tubo neurale dorsale (a livello di somite da 1 a 7 nel video).
- Tagliare bilateralmente tra il tubo neurale e i somiti per separare il tubo neurale dai tessuti circostanti, senza danneggiare le somiti.
- Separare molto delicatamente il tubo neurale dal notocordo sottostante, che dovrebbe rimanere intatto. Si noti che l'escissione del tubo neurale di successo lascerà tutti i tessuti circostanti perfettamente intatti (Figura 2).
- Rimuovere il tubo neurale asportato aspirarlo in una micro-pipetta di vetro, quindi scartare.
- Registra il livello di ablazione del tubo neurale in un libro di laboratorio. L'embrione ospite è ora pronto a ricevere il tubo neurale del donatore.
5. Preparare il tessuto di innesto del donatore
- Selezionare un embrione GFP con finestre abbinato al palco visualizzando sotto un microscopio stereo fluorescente con filtro FITC. La fluorescenza GFP rende molto facile visualizzare i somiti e inscenare l'embrione.
- Una volta identificato un embrione abbinato allo stadio, rimuovere l'embrione dall'uovo facendo 4 incisioni, con forbici primaverili Pascheff-Wolff (Figura 1C, l) in una forma rettangolare intorno all'embrione, quindi raccoglierlo delicatamente con un cucchiaio embrionale.
- Posizionare l'embrione in un vetro quadrato con una base polimerica sylgard. Agitare delicatamente l'embrione con Dumont #5 pinzette per rimuovere il tuorlo attaccato. Rimuovere la membrana vitellina e inchiodare l'embrione sulla base polimerica utilizzando perni di minuzie inossidabile (Figura 1C).
- Usando le forbici primaverili, fai 4 incisioni in forma rettangolare attorno al tubo neurale e ai somiti circostanti, nella stessa regione che è stata rimossa dall'embrione ospite.
- Utilizzando una pipetta di trasferimento in plastica, trasferire il tubo neurale e i tessuti somite dall'embrione GFP donatore in un vetro da orologio contenente lo 0,2% di pancreatina in Pen/ Strep PBS.
- Consentire alla digestione enzimatica di procedere per 10 minuti a RT per aiutare a separare i tessuti. Dopo l'incubazione nell'enzima, utilizzare perni di minuzia inossidabile montati su un manico per separare manualmente il tubo neurale da tutti i tessuti adiacenti.
- Utilizzando una micro-pipetta di vetro, trasferire il tubo neurale dissociato su un altro vetro da orologio contenente DMEM + 10% siero(ad esempio,capra, cavallo o vitello fetale) sul ghiaccio, per risciacquare la pancreatina in eccesso e interrompere la digestione enzimatica. Dopo 5 minuti, il tubo neurale sezionato è pronto per essere innestato ortotopicamente nell'ospite del pulcino(Figura 2 e S1).
6. Innestare il tessuto
- Utilizzando una micro-pipetta di vetro, trasferire con cura il tubo neurale sezionato dal vetro dell'orologio all'embrione ospite. Posizionare il tubo neurale nel corretto orientamento anteriore-posteriore e spingere delicatamente l'espianto adiacente alla regione asportata dell'ospite del pulcino utilizzando il micro-bisturi. Lasciare un piccolo frammento di ectoderma attaccato, o tagliando un piccolo nick in, la superficie dorsale per identificare l'orientamento del tubo neurale.
- Se necessario, utilizzare il micro-bisturi per tagliare l'espianto alla dimensione esatta della regione asportata.
- Guidare delicatamente il tubo neurale nella regione ablata e posizionarlo in modo tale che il lato dorsale sia correttamente orientato. Utilizzare una microtubetta di vetro, montata su un tubo della bocca, per rimuovere PBS e/o fluido che circonda l'innesto. Questo aiuta il donatore e i tessuti ospiti ad aderire e l'innesto a stabilirsi.
- Sigillare l'intera finestra con nastro chiaro largo 24 mm per evitare disidratazione e contaminazione.
- Etichettare l'embrione chimerico contrassegnando con una matita sul guscio d'uovo e registrare il suo numero nel libro di laboratorio. Restituire l'uovo all'incubatrice per un ulteriore sviluppo.
7. Iniettare dii nei vasi sanguigni dell'embrione ospite
- Nel momento sperimentale desiderato (qui, 3 - 10 giorni dopo), recuperare l'embrione chimerico dall'incubatore e rimuovere il nastro chiaro usando forbici dritte per ottenere l'accesso all'embrione all'interno dell'uovo.
- Se necessario, ingrandire la finestra nel guscio usando le forbici. Fare attenzione a non danneggiare la membrana corioallantoica se è attaccata al guscio, il che comporterebbe un'emorragia e metterebbe a rischio l'etichettatura dei vasi sanguigni.
- Scegli una vena accessibile sul tuorlo assicurandoti che il flusso sanguigno sia diretto verso l'embrione. Scegliere un punto di ramificazione di una delle vene vitelline(Figura 3B, C).
NOTA: A E6.5 - E7.5, la membrana corioallantoica potrebbe dover essere delicatamente spostata da parte con una pinzetta per accedere alle vene del tuorlo. Dopo E8.5, l'unica opzione è iniettare in una delle vene della membrana corioallantoica poiché, in questa fase, la membrana corioallantoica copre completamente l'embrione. - Rimuovere la membrana vitellina sopra il punto di iniezione scelto utilizzando due #5 Dumont strappando in direzioni opposte.
- Rompere un ago di vetro tirato utilizzando un #5 Dumont e regolarne il diametro in base alle dimensioni approssimative della vena prima di caricarlo con CellTracker CM-DiI. Fare la soluzione di serie DiI a 40 μg/μl in DMSO e conservare a -20 °C. Preparare la soluzione di lavoro in saccarosio/PBS da 0,3 M ad una concentrazione di 4 μg/μl.
- Aspirare tra 5 e 10 μl di DiI in saccarosio/PBS da 0,3 M nell'ago usando l'aspirazione con un tubo della bocca. Gli embrioni più vecchi potrebbero richiedere fino a 25 μl o più. Dall'E8.5, gli embrioni hanno vene più grandi e muscolose, che potrebbero dover essere tenute in posizione con un #5 Dumont prima di pugnalare con l'ago di vetro caricato da DiI.
- Inserire rapidamente l'ago nella vena e soffiare costantemente con il tubo della bocca per consentire al DiI di unire lentamente il flusso sanguigno senza formare un coagulo. In alternativa, utilizzare un iniettore di pressione per la consegna DiI.
8. Raccogliere embrioni per il se sezione o l'esame wholemount
- Per trattenere il maggior dii all'interno dell'embrione possibile, raccogliere l'embrione immediatamente dopo l'iniezione raccogliendolo su un cucchiaio perforato e tagliando i vasi sanguigni e i tessuti connettivi con un paio di forbici dritte, per liberare l'embrione dal tuorlo.
- Rimuovere eventuali membrane sciolte e sezionare gli organi di interesse(cioè, i polmoni e il tratto digestivo in questo tutorial), facendo molta attenzione a non comprimere il tessuto, che crea diffusione del DiI. Fissare immediatamente i tessuti per immersione in PFA al 4% per 1 - 2 ore a RT.
- Risciacquare il tessuto per 5 minuti in PBS, quindi 15 minuti in PBS contenente 5 μg/ml DIPI. Montare i campioni su una diapositiva del microscopio a ponte per l'esame dell'intero supporto o incorporarli per la criosezione.
Representative Results
La figura 1 mostra gli strumenti tipici necessari per effettuare l'isolamento microchirurgico e il trapianto del tubo neurale. La figura 2 mostra la procedura di trapianto. Dopo il trapianto gli embrioni vengono vengono vengono vengono vengono Ciò comporta l'esame dell'embrione sotto un microscopio a fluorescenza stereo, tipicamente la mattina dopo la microchirurgia, per la presenza di NCC derivato dall'innesto (GFP+). Se il trapianto è stato un successo, allora GFP + NCC può essere osservato in prossimità del tubo neurale e nei percorsi di migrazione precoce che portano verso il prenega. Se la procedura non ha avuto esito positivo, GFP+ NCC non verrà osservato al di fuori del tubo neurale, o se sono presenti nell'host potrebbero essere in numero minore. Questi embrioni falliti vengono scartati. In genere, 5-8 trapianti di tubi neurali vengono eseguiti in un giorno e di questi l'80% ha successo. I motivi del trapianto di tubo neurale non riuscito includono la morte dell'embrione a causa di danni ai tessuti subiti durante la microchirurgia o l'incapacità del tubo neurale di integrarsi nell'embrione ospite. Quest'ultimo può derivare da uno scarso posizionamento del tubo neurale all'interno dell'ospite o da un tubo neurale di scarsa qualità a causa della scarsa tecnica di dissezione o dell'eccessiva esposizione all'enzima di dissociazione. La fase iniziale di screening, così come analoghi esami successivi per le cellule GFP+, è utile in quanto significa che il tempo e le risorse non vengono sprecati eseguendo esperimenti su embrioni che non hanno NCC marcato GFP all'interno dell'intestino.
La figura 3 mostra la procedura per l'iniezione di DiI dei vasi sanguigni. L'efficienza/successo della tecnica di iniezione DiI dipende da: in primo luogo, tagliare l'ago di iniezione al diametro ottimale per la vena mirata, secondo un gesto preciso quando si inserisce l'ago nella vena (in modo da non perforare l'altro lato), e terzo evitare che l'ago si colleghi durante l'iniezione soffiando a velocità costante. Se uno di questi tre parametri viene eseguito in modo errato, l'embrione dissapegna o avrà bisogno di diverse ore per riprendersi prima che si faccia un secondo tentativo poiché l'emorragia rende quasi impossibile reiettare immediatamente. Gli embrioni di successo devono essere selezionati immediatamente visualizzando al microscopio a stereofluorescenza e devono essere sezionati rapidamente. Negli embrioni di successo, i vasi sanguigni etichettati DiI sono presenti in tutto l'embrione (Figura 3C,D) compresi i letti capillari (Figura 3D).
Al momento della raccolta degli embrioni e dell'esame di sezioni tissutali o tratti gastrointestinali integrali, risultati tipici rivelano che GFP+ NCC all'interno dell'ENS primitivo e la struttura fine delle reti di vasi sanguigni intestinali con etichetta DiI (Figura 4) I preparati wholemount possono essere esaminati utilizzando la microscopia confocale in base alla quale le pile di immagini producono ricostruzioni tridimensionali (3D) che mostrano le interrelazioni tra le proiezioni fini delle cellule GFP+ ENS e il sistema vascolare macchiato DiI(figura 4 A-C; G-I; Video 1 e 2).

Figura 1. Strumenti di microchirurgia consigliati. (A) micro-bisturi a forma di ago da cucito. (B) pietra fine dell'Arkansas per modellare un micro-bisturi. c a) forbici dritte, b) forbici curve, e) siringa da 5 ml con ago ipodermico da 181/2 G, d) pipetta di plastica, e) portaovo su misura, f) inchiostro nero, g) vetro quadrato per orologi, h) vetro quadrato con base di sylgard nero, i) micro-bisturi sul supporto dell'ago, j) spille minuzie, k) minuzza o ago di tungsteno sul supporto dell'ago, l) Forbici a molla Pascheff-Wolff, m) Dumont #5 pinzette, n) cucchiaio perforato, oi) ago a trasferimento corto tirato dal fuoco, oii) ago inchiostrato lungo tirato a fuoco, p) tubo della bocca. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 2. Trapianto di tubo neurale intraspecie. Le immagini del tubo neurale dell'embrione di pulcino /GFP sono state modificate da Delalande et al. 12. La vascolarizzazione non è necessaria per la colonizzazione intestinale da parte delle cellule della cresta neurale enterica. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 3. Iniezione di dii endovenosa. (A) Strumenti raccomandati: a) CellTracker CM-DiI goccia su parafilm, b) ago di iniezione di vetro tirato, e) tubo della bocca. (B) Diagramma schematico dell'iniezione di DiI endovenosa nell'embrione di pulcino chimerico E4. (C) nell'iniezione endovenosa ovo DiI che mostra un ago di vetro fine contenente DiI inserito nella vena (freccia). (D) E4 embrione chimerico post iniezione DiI (rosso) con tubo neurale GFP+ (freccia). (E) Rete di vasi sanguigni fini macchiati di dii in un embrione vivo, 24 ore dopo l'iniezione. Br: cervello; H: cuore; LB: bocciolo d'arto; A: allantois. Le immagini in (C) e (D) sono state modificate da Delalande et al. 12 di cui: la commissione per i La vascolarizzazione non è necessaria per la colonizzazione intestinale da parte delle cellule enteriche della cresta neurale. Clicca qui per visualizzare una versione più grande di questa figura.

Figura 4: Risultati rappresentativi nello stomaco e nel feco di un embrione di pulcino E5.5. (A-C) Ricostruzione tridimensionale (3D) di una pila di immagini confocali nella regione dello stomaco che mostra (D) le cellule di cresta neurale enterica GFP+ (ENCC) (E) il sistema vascolare macchiato DiI e (F) un'immagine unita di entrambe le reti D-F Sezioni istologiche a livello dello stomaco che mostra (G) l'ENCC GFP+ (H) il sistema vascolare macchiato DiI e (I) un'immagine unita di entrambe le reti. I nuclei sono macchiati di DAPI (ciano). (G-H) ricostruzione 3D di una pila di immagini confocali nella regione del caecum che mostra (A) il fronte di migrazione ENCC GFP+ in verde, (B) il sistema vascolare macchiato DiI in rosso e (C) un'immagine unita di entrambe le reti. Le immagini (A-F) sono state modificate da Delalande et al. 12 di cui: la commissione per i La vascolarizzazione non è necessaria per la colonizzazione intestinale da parte delle cellule enteriche della cresta neurale. Clicca qui per visualizzare una versione più grande di questa figura.

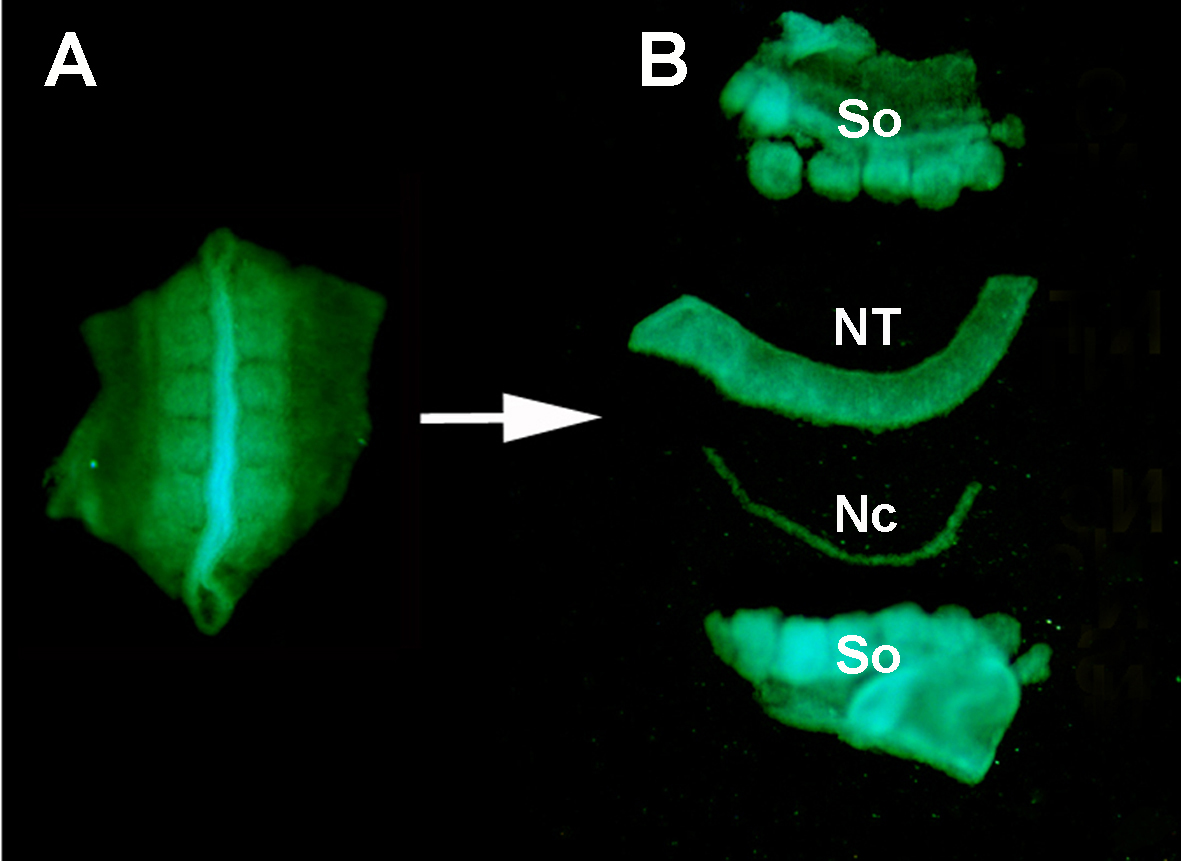
Figura S1. Isolamento di un tubo neurale GFP+ donatore dai tessuti circostanti mediante digestione enzimatica e microsezione. (A) Tubo neurale GFP+ e somiti adiacenti sezionati dall'embrione donatore. (B) Tubo neurale isolato dopo la digestione della pancreatina e la microsezione utilizzando perni di minuzie inossidabili. Quindi: somiti; NT: tubo neurale; Nc: Notochord.

Video 1. Rotazione tridimensionale a 360° dell'immagine nella figura 4C, che mostra il sistema vascolare e l'ENCC nello stomaco a E5.5 (HH27-28). Clicca qui per vedere questo video.

Video 2. Rotazione tridimensionale a 360° dell'immagine nella figura 4I, che mostra il sistema vascolare e il fronte di migrazione ENCC nella regione del caecum a E5.5 (HH27-28). Clicca qui per vedere questo video.
Discussion
Il metodo di innesto del tubo neurale intraspecie, combinato con l'etichettatura dei vasi sanguigni qui descritta, sfrutta appieno la facilità di accesso dell'embrione aviario all'interno dell'uovo (rispetto ad altri embrioni di vertebrati) per studiare il co-sviluppo di un elemento del sistema nervoso autonomo (l'ENS) e del sistema vascolare.
Per l'etichettatura dei derivati NCC, il metodo di innesto intraspecie di pulciniGFP-pulcini che descriviamo ha una serie di vantaggi rispetto al classico metodo di chimera quaglia-pulcino che è stato stabilito oltre 40 annifa 1-3. In primo luogo, sotto la luce FITC, la fluorescenza GFP è estremamente luminosa, al punto che le cellule GFP+ sono facilmente percepibili negli embrioni chimerici vivi. Ciò consente di verificare il successo dell'innesto in ovo , mentre l'innestoquaglia-pulcino richiede che l'embrione venga ucciso, lavorato e immunosostenibile utilizzando QCPN, prima che il successo dell'innesto possa essereaccertato 2. In secondo luogo, l'espressione GFP nelpulcino transgenico GFP è citoplasmatica, quindi non solo etichetta i corpi cellulari, ma consente anche di visualizzare le proiezioni delle celluletrapiantate 22. Ciò consente di osservare reti neuronali complesse ad alta risoluzione (si noti che le proiezioni fini sono meglio visualizzate quando il campione è immunosostenibile con anticorpi anti-GFP). Poiché l'etichettatura QCPN è limitata al nucleo cellulare delle quaglie, tali reti non vengono rivelate usando chimere di quaglia-pulcini. In terzo luogo, l'innesto intraspecie elimina eventuali differenze di specie tra le cellule all'interno dell'embrione chimerico. Poiché gli embrioni di quaglia hanno un periodo di incubazione più breve rispetto al pulcino (19 giorni contro 21 giorni) è stato suggerito che le cellule quaglie hanno un tasso di proliferazione più elevato rispetto alle cellule dei pulcini, il che potrebbe potenzialmente influenzare lo sviluppo dei tessuti chimerici23. È interessante notare anche che nelle piante è stato dimostrato che l'innesto di interspecie potrebbe produrre ampie alterazioni nei modelli di metilazione del DNA nell'ospite 24. In quarto luogo,chick GFP facilita gli esperimenti di trapianto di schiena per affrontare argomenti come il destino ncc e l'impegno cellulare25. In quinto luogo, ilGFP transgenico dei pulcini è utile anche per molte altre tecniche tra cui lo smistamento FACS delle sottopopolazioni cellulari GFP+, la coltura organotipica di organi contenenti cellule GFP+, la manipolazione genetica del tessuto innestato GFP+ tramite elettroporazione dei plasmidi di espressione26e altre tecnologie di imaging come la tomografia a proiezione ottica27.
L'approccio di trapianto di tubi neurali può essere modificato sostituendo microchirurgicamente quantità più brevi di tubo neurale. Utilizzando segmenti più piccoli di tubo neurale, la microchirurgia è potenzialmente meno dannosa per l'embrione e la sopravvivenza può essere migliorata. Tuttavia, il lato negativo del trapianto di meno tubo neurale è che il numero di NCC GFP + nell'ospite sarà ridotto. Gli utenti potrebbero cercare di raggiungere un equilibrio tra la quantità di tubo neurale trapiantato per dare una sopravvivenza ottimale degli embrioni e il numero di NCC GFP + all'interno dell'intestino ospite sufficiente a dare risultati informativi.
Per la verniciatura dei vasi, DiI ha il vantaggio che la sua fluorescenza è molto luminosa e robusta. Inoltre, ha la capacità di diffondersi durante la fissazione assicurando la colorazione dei migliori capillari aperti. Poiché si tratta di un colorante vitale, gli embrioni possono sopravvivere alla procedura di iniezione e continuare a svilupparsi con un sistema vascolare macchiato (fino a 24 ore nelle nostre mani, anche se la colorazione diventa più puncata nel tempo, vedi Figura 3E). La combinazione diinnesto GFP del pulcino con verniciatura vascolare DiI è quindi compatibile con l'imaging dal vivo. Oltre a tutti questi vantaggi, è importante notare che l'iniezione vascolare etichetta solo vasi luminati e quindi non identifica capillari non aperti, cellule di punta endoteliali o cellule endoteliali isolate. Tuttavia, ulteriori progressi nella transgenesi aviaria potrebbero fornire nuovi modi per aggirare tali problemi, come esemplificato da esperimenti che utilizzano embrioni di quaglia Tg(tie1:H2B-eYFP) per studiare la morfogenesi vascolare28. Un'altra limitazione di questa tecnica è che, per un'efficace etichettatura dei vasi negli embrioni a E7,5 e oltre, è necessario iniettare maggiori quantità di colorante, il che può rendere costosi gli esperimenti. Tuttavia, una modifica della tecnica potrebbe includere l'etichettatura dei vasi sanguigni a basso costo utilizzando l'inchiostro dell'evidenziatore14, anche se questo approccio non è stato provato nelle nostre mani.
Le fasi critiche delle procedure includono il processo di visualizzazione dell'embrione iniettando inchiostro sotto il blastodisc. Se la membrana che copre il tuorlo viene strappata dall'ago pieno di inchiostro in questa fase, la sopravvivenza dell'embrione è gravemente compromessa. Inoltre, è importante, quando si prepara un tubo neurale del donatore, che il tessuto non sia lasciato per un tempo eccessivamente lungo nella pancreatina (considerare circa 10 minuti come massimo). L'esposizione prolungata alla pancreatina danneggia il tessuto e il tubo neurale è quindi difficile da maneggiare e non si incorporerà bene nell'ospite. Acquisire esperienza della tecnica di iniezione DiI su embrioni di tipo selvatico è essenziale prima di iniettare embrioni chimerici, poiché è generalmente possibile un solo tentativo di iniezione per ogni embrione. Il volume di dii e il diametro dell'ago sono parametri critici per ogni embrione e devono essere valutati in base al tipo selvaggio, ai controlli abbinati alla fase.
In conclusione, il nostro metodo di doppia etichettatura del trapianto di tubi neurali e la pittura dei vasi DiI negli embrioni di pulcini vivi possono essere utilizzati per indagare le interrelazioni tra NCC e reti di vasi sanguigni durante l'organogenesi. Considerando che i meccanismi responsabili della corretta innervazione e vascolarizzazione bersaglio durante lo sviluppo degli organi sono ancora ampiamente sconosciuti, questa metodologia ha un potenziale per le scoperte future in questo campo.
Disclosures
Gli autori dichiarano di non avere interessi finanziari concorrenti.
Acknowledgments
Le uova di gallina GFP fecondate sono state fornite dalla prof.ssa Helen Sang, the Roslin Institute, e dall'Università di Edimburgo, Nel Regno Unito. Il Roslin Transgenic Chicken Facility è finanziato dal Wellcome Trust e dal Biotechnology and Biological Sciences Research Council (BBSRC). Il lavoro è stato in parte finanziato, e nt supportato, dalla Great Ormond Street Hospital Children's Charity, Londra, Regno Unito. Gli autori ringraziano Ben Jevans, UCL Institute of Child Health, per l'aiuto nella preparazione degli embrioni per l'innesto.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}