

Summary
Her rapporterer vi dobbel merking av nevrale kamceller og blodårer ved hjelp av kyllingGFP neural tube intraspecies podning kombinert med intra-vaskulær DiI injeksjon. Denne eksperimentelle teknikken lar oss samtidig visualisere og studere utviklingen av det NCC-avledede (enteriske) nervesystemet og det vaskulære systemet, under organogenese.
Abstract
Alle utviklende organer må kobles til både nervesystemet (for sensorisk og motorisk kontroll) samt det vaskulære systemet (for gassutveksling, væske- og næringsforsyning). Følgelig utvikler både de nervøse og vaskulære systemene seg sammen og deler slående likheter i forgreningsarkitekturen. Her rapporterer vi embryonale manipulasjoner som lar oss studere samtidig utvikling av nevralt kam-avledet nervevev (i dette tilfellet det enteriske nervesystemet) og det vaskulære systemet. Dette oppnås ved å generere kylling chimeras via transplantasjon av diskrete segmenter av nevralrøret, og tilhørende nevral kam, kombinert med vaskulær DiI-injeksjon i samme embryo. Vår metode bruker transgen kyllingGFP embryoer for intraspecies podning, noe som gjør transplantasjonsteknikken kraftigere enn den klassiske vaktel-chick interspecies podningsprotokollen som brukes med stor effekt siden 1970-tallet. ChickGFP-chick intraspecies podning letter avbildning av transplanterte celler og deres projeksjoner i intakt vev, og eliminerer eventuelle skjevheter i celleutvikling knyttet til artsforskjeller. Denne metoden drar full nytte av enkel tilgang til fugleembryoet (sammenlignet med andre virveldyrembryoer) for å studere samutviklingen av det enteriske nervesystemet og det vaskulære systemet.
Introduction
Kyllingembryoet er en uvurderlig modellorganisme i virveldyrutviklingsbiologi, ikke minst fordi utviklingen i ovo tillater eksperimentelle manipulasjoner som ellers er umulige å utføre i vertebrater som utvikler seg i utero. Denne tilgjengeligheten og enkel manipulasjon har ført til at kyllingembryoet spiller nøkkelroller i mange seminalfunn innen utviklingsbiologi. Blant de kraftigste teknikkene har vært bruk av vaktelkylling chimeric embryoer for å studere celle skjebne, en metode pioner av professor Nicole Le Douarin i 1970-tallet1-3. Spesielt har vaktel-chick chimeras vært spesielt nyttig for å genetisk markere og følge svært trekkende nevrale kamcellepopulasjoner (NCC) under tidlig utvikling. NCC er en multipotent populasjon av trekkceller, som oppstår i dorsal ektoderm ved marginene i nevralrøret, som gir opphav til et bredt spekter av celletyper gjennom vertebratembryoet. Disse inkluderer kraniofaciale strukturer (brusk, bein, muskler), nevroner og glia (i sensoriske og autonome nervesystemer), melanocytter og en subpopulasjon av celler i det endokrine systemet2,4,5. En av de viktigste faktorene som påvirker NCC-skjebnen er deres opprinnelige plassering langs den fremre bakre aksen til nevralrøret. For eksempel oppstår enterisk NCC, som gir opphav til nevronene og glia i det enteriske nervesystemet (ENS), fra to diskrete underpopulasjoner: den første som ligger i vagal (kaudal hindbrain) -regionen, og den andre i sakralområdet i nevralrøret6-13. Inter eller intra-arter podning av de tilsvarende områdene i nevralrøret har vært de valgte teknikkene for å permanent merke disse cellene og deretter tillate sporing, fra fødselen på marginene i nevralrøret, til deres endelige destinasjoner i fordøyelseskanalen6,7,10.
En annen embryonal manipulasjon lettere å utføre hos kyllinger, sammenlignet med andre dyremodeller, er den vitale merkingen av det vaskulære systemet. Faktisk, som kyllingembryoet utvikler seg, ligger det på toppen av et ekstra embryonalt vaskulært nettverk som sirkulerer oksygen og næringsstoffer fra eggeplommen. Dette tilgjengelige vaskulære nettverket, som ligger på overflaten av eggeplommen, kan brukes som en inngangsport for å merke embryoets utviklende vaskulære system under organogenese12,14-17. Intravaskulær injeksjon av ulike fargestoffer, som lipofil fargestoff diI, gjør det mulig å avgrense / flekke alle de luminiserte karene i det nascent vaskulære nettverket.
Fordi utviklende organer må kobles til både nervesystemet (for sensorisk og motorisk kontroll) samt det vaskulære systemet (for gassutveksling, væske- og næringsforsyning), utvikler de to nettverkene sammen og deler slående likheter i forgreningsarkitekturen18-20. Her rapporterer vi embryonale manipulasjoner som gjør at vi kan studere samtidig utvikling av den NCC-avledede ENS, sammen med det vaskulære systemet, under organogenese. Dette oppnås ved å generere kylling chimeras via transplantasjon av diskrete segmenter av nevralrøret, inkludert nevral kam, kombinert med vaskulær DiI injeksjon. Som et fremskritt fra vaktel-kylling chimeras, bruker vår metode transgene GFP chick embryoer for intraspecies podning, noe som gjør transplantasjonsteknikken kraftigere, når det gjelder bildeceller og deres projeksjoner, og eliminerer potensielle skjevheter knyttet til artsforskjeller.
Protocol
1. Tilberedning av mikro-skalpell for Neural Tube Ablations
- Form en mikro-skalpell fra en kommersielt tilgjengelig stålsynål.
- Flat først nålen på begge sider ved hjelp av et slipeskive montert på en drevet benkkvern.
- Begynn å forme skalpellen, først på en grov gradert Arkansas-stein ved hjelp av en kontrollert sirkulær bevegelse, i alternative retninger, på begge sider av nålen.
- Fortsett de samme slipebevegelsene på en ekstra fin klasse Arkansas-stein for å forme en ultrafin mikroskapell, med en veldefinert skjærekant (Figur 1A, B).
MERK: Alternativer til mikro-skalpellen kan være elektrolittisk skjerpede nåler, kommersielt tilgjengelige wolframnåler eller trukket glassnåler.
2. Inkuber wild type og GFP egg til ønsket stadium
- Oppbevar befruktede kyllingegg og transgene GFP kyllingegg i en avkjølt inkubator ved 14 - 15 °C før inkubasjon, da utviklingen stoppes ved denne temperaturen. Oppbevar egg i noen dager, opptil en uke.
- For å starte utviklingen, plasser vill type og GFP egg på et brett horisontalt og inkuber samtidig i en fuktet (58 - 60%) inkubator ved 37,5 ºC, slik at embryoer er i matchende stadier for nevral rørtransplantasjon.
- For å oppnå embryoer på 10 - 12 somite utviklingsstadium for å utføre vagal nevral tube podning, inkubere egg i 1,5 dager (33 - 38 timer) og stadium embryoer i henhold til utviklingstabellene hamburger og Hamilton21.
3. Forbered egg til vindus- og podning
- Flytt ett egg om gangen til en skreddersydd eggholder for vindu. Lag et lite hull i eggeskallet ved å trykke gjentatte ganger, med rett saks, på den øvre overflaten av den spisse enden av egget.
- Fjern 2 - 3 ml albumin fra egget med en hypodermisk nål på 181/2 G og 5 ml sprøyte. Fjerning av albumin senker eggeplommen i egget og letter etterfølgende vinduer uten å forårsake skade på embryoet.
- Forkast albuminet. Forsegle hullet med en liten stripe med klart tape kuttet i størrelse med fin saks.
- Bruk buet saks, trykk på et annet hull i eggeskallets øvre overflate. Sett spissen av saksen inn i hullet, og hold saksen parallelt med benken, arbeid i en sirkulær bevegelse for å kutte et vindu med ~ 2 cm diameter på toppen av skallet.
- Hold saksen i stasjonær stilling og roter egget. Kast den fjernede disken med eggeskall. På E1.5 er embryoet gjenkjennelig som en mørkere gul disk på toppen av eggeplommen.
- Fjern eventuelle skallrester som har falt i egget ved hjelp av pinsett. Kast eventuelle unfertilized egg (identifisert av en liten hvit flekk på toppen på den ellers lyse gule eggeplommen).
4. Forbered vertsembryoet for å motta podet vev
- Juster stereomikroskopet til øyenivå og optimaliser retningen til gooseneck lyskilde for å tilstrekkelig belyse embryoet uten å forårsake refleksjoner.
- For å visualisere embryoet riktig, injiser en liten mengde indisk blekk under midten av den mørkere gule disken, ved hjelp av et munnrør og en trukket glassmikropipette (Figur 1C, Oii).
- Forbered blekket 50:50 med PBS som inneholder Penicillin/Streptomycin ved 100 μg/ml endelig konsentrasjon. Sett mikropipetten gjennom eggeplommemembranen utenfor blastodermens omkrets, og vinkle spissen forsiktig rett under embryoet.
- Lever blekk under embryoet ved å blåse på munnrøret. Hvis munnpipettering ikke er tillatt, bruk en 1 ml sprøyte i stedet. Pass på at du ikke introduserer noen luftbobler under embryoet, noe som kan føre til forurensning, og fjern deretter glassmikropipetten forsiktig. Dette er et delikat skritt som kan føre til embryoets død hvis det ikke gjøres med presisjon.
- Stage embryoet ved referanse til Hamburger og Hamilton21 og registrere scenen i en lab bok.
- Ved hjelp av en skreddersydd mikro-skalpell (eller en fin wolfram nål) montert på en nålholder, lage en svært liten gash i vitelline membranen, ved siden av området der mikrokirurgi vil bli utført.
- Påfør forsiktig 2 - 3 dråper PBS over membrantåren (ved hjelp av en glassmikropipette og munnrøret) for å skape plass mellom embryoet og membranen. Klipp et større vindu i membranen for å eksponere hele regionen der mikrooperasjonen vil finne sted.
- Fjern det nevrale rørområdet av interesse ved hjelp av mikro-skalpell, som starter med rostrale og kaudale tverrsnitt over hele dorsale nevrale røret (på nivået av somitt 1 til 7 i videoen).
- Klipp bilateralt mellom nevralrøret og somittene for å skille nevralrøret fra omkringliggende vev, uten å skade somittene.
- Svært forsiktig skille nevralrøret fra den underliggende notokord, som skal forbli intakt. Vær oppmerksom på at vellykket nevral røreksisjon vil etterlate alle omkringliggende vev helt intakte (Figur 2).
- Fjern det utskilte nevrale røret ved å aspirere det inn i en mikropipette i glass, og kast det deretter.
- Registrer nivået av nevral tube ablasjon i en lab bok. Vertsembryoet er nå klart til å motta donor nevralrøret.
5. Forbered donortransplantvevet
- Velg et vindusbasert, scenetilpasset GFP-embryo ved å se under et fluorescerende stereomikroskop med FITC-filter. GFP-fluorescensen gjør det veldig enkelt å visualisere somittene og iscenesette embryoet.
- Når et stadium-matchet embryo er identifisert, fjern embryoet fra egget ved å lage 4 snitt, med Pascheff-Wolff vårsaks (Figur 1C, l) i rektangelform rundt embryoet og plukk det forsiktig opp med en embryoskje.
- Plasser embryoet i et firkantet klokkeglass med en sylgardpolymerbase. Rist forsiktig embryoet med Dumont #5 pinsett for å fjerne eventuelle vedlagte eggeplommer. Fjern vitellinemembranen og fest embryoet på polymerbasen ved hjelp av rustfrie minutienpinner (Figur 1C).
- Bruk vårsaksen til å lage 4 snitt i rektangulær form rundt nevralrøret og omkringliggende somitter, i samme region som er fjernet fra vertsembryoet.
- Bruk en plastoverføringspipette, overfør nevralrøret og somittvevet fra donor GFP-embryoet til et klokkeglass som inneholder 0,2% pankreatin i Pen / Strep PBS.
- La enzymatisk fordøyelse fortsette i 10 minutter ved RT for å skille vevet. Etter inkubasjon i enzym, bruk rustfrie minutien pinner montert på et håndtak for å manuelt skille nevralrøret fra alle tilstøtende vev.
- Bruk en mikropipette i glass, overfør det dissosierte nevrale røret til et annet klokkeglass som inneholder DMEM + 10% serum (f.eks.geit, hest eller fosterkalv) på is, for å skylle overflødig bukspyttkjertel og stoppe enzymatisk fordøyelse. Etter 5 min er det dissekerte nevrale røret klar til å bli podet ortopisk inn i kyllingverten (Figur 2 og S1).
6. Graft vevet
- Bruk en mikropipette i glass, og overfør forsiktig det dissekerte nevrale røret fra klokkeglasset til vertsembryoet. Plasser nevralrøret i riktig fremre bakre orientering og skyv forsiktig utvisningen ved siden av den utskilte regionen til kyllingverten ved hjelp av mikro-skalpellen. La et lite fragment av ektoderm festet til, eller ved å kutte en liten nick i, den dorsale overflaten for å identifisere orienteringen av nevralrøret.
- Bruk om nødvendig mikro-skalpellen til å trimme utvisningen til den nøyaktige størrelsen på det utskilte området.
- Før nevralrøret forsiktig inn i det ablerte området og plasser det slik at dorsalsiden er riktig orientert. Bruk en mikropipette i glass, montert på et munnrør, for å fjerne PBS og/eller væske rundt transplantatet. Dette hjelper donoren og vertsvevet til å holde seg og graftet til å bli etablert.
- Forsegle hele vinduet med 24 mm bredt klart tape for å forhindre dehydrering og forurensning.
- Merk det chimeriske embryoet ved å merke med en blyant på eggeskallet og registrere nummeret i labboken. Returner egget til inkubatoren for videre utvikling.
7. Injiser diI i blodårene til vertsembryo
- På ønsket eksperimentelt tidspunkt (her, 3 - 10 dager senere), hent det chimeriske embryoet fra inkubatoren og fjern det klare båndet ved hjelp av rett saks for å få tilgang til embryoet i egget.
- Om nødvendig forstørrer du vinduet i skallet ved hjelp av saksen. Vær forsiktig så du ikke skader den chorioallantoiske membranen hvis den er festet til skallet, noe som vil føre til blødning og sette blodkarmerkingen i fare.
- Velg en tilgjengelig vene på eggeplommen, og sørg for at blodstrømmen er rettet mot embryoet. Velg et forgreningspunkt for en av vitelline-venene (Figur 3B, C).
MERK: På E6,5 - E7,5 kan det hende at den chorioallantoiske membranen må flyttes forsiktig til side med pinsett for å få tilgang til eggeplommeårene. Etter E8.5 er det eneste alternativet å injisere i en av de chorioallantoiske membranårene siden, på dette stadiet, dekker den chorioallantoiske membranen fullt ut embryoet. - Fjern vitellinemembranen over det valgte injeksjonspunktet ved hjelp av to Dumont-#5 pinsett ved å rive i motsatte retninger.
- Bryt en trukket glassnål ved hjelp av en Dumont #5 og juster diameteren til den omtrentlige størrelsen på venen før lasting med CellTracker CM-DiI. Lag DiI-lagerløsningen ved 40 μg/μl i DMSO og oppbevar den ved -20 °C. Forbered arbeidsløsningen i 0,3 M sukrose/PBS i en konsentrasjon på 4 μg/μl.
- Aspirer mellom 5 - 10 μl DiI i 0,3 M sukrose/PBS inn i nålen ved hjelp av sug med et munnrør. Eldre embryoer kan kreve opptil 25 μl eller mer. Fra E8.5 har embryoer større, mer muskuløse årer, som kanskje må holdes på plass med en Dumont-#5 før de stikker med dii-lastet glassnål.
- Sett nålen raskt inn i venen og blås jevnt med munnrøret slik at diI kan bli med blodstrømmen sakte uten å danne en blodpropp. Alternativt kan du bruke en trykkinjektor for DiI-levering.
8. Høst embryoer for seksjonering eller full monteringsundersøkelse
- For å beholde så mye DiI i embryoet som mulig, høst embryoet umiddelbart etter injeksjonen ved å øse det på en perforert skje og kutte blodkarene og bindevevet med et par rette saks, for å frigjøre embryoet fra eggeplommen.
- Fjern eventuelle løse membraner og disseker ut organene av interesse (dvs.lungene og fordøyelseskanalen i denne opplæringen), og pass på at du ikke komprimerer vevet, noe som skaper diffusjon av DiI. Fest umiddelbart vevet ved nedsenking i 4% PFA i 1 - 2 timer ved RT.
- Skyll vevet i 5 min i PBS, deretter 15 min i PBS som inneholder 5 μg/ml DAPI. Monter prøvene på et brokoblet mikroskopsklie for undersøkelse av hele braketten eller bygg dem inn for kryo-seksjonering.
Representative Results
Figur 1 viser typiske instrumenter som kreves for å utføre mikrokirurgisk isolasjon og transplantasjon av nevralrøret. Figur 2 viser transplantasjonsprosedyren. Etter transplantasjon blir embryoer screenet for transplantasjonssuksess. Dette innebærer å undersøke embryoet under et stereofluorescensmikroskop, vanligvis morgenen etter mikrokirurgi, for tilstedeværelse av graft-avledet (GFP +) NCC. Hvis transplantasjon har vært en suksess, kan GFP + NCC observeres i nærheten av nevralrøret og i tidlige migrasjonsveier som fører mot foregut. Hvis prosedyren ikke har vært vellykket, vil GFP + NCC ikke bli observert utenfor nevralrøret, eller hvis de er til stede i verten, kan de være i mindre antall. Disse mislykkede embryoene kastes. Vanligvis utføres 5-8 nevrale rørtransplantasjoner på en dag, og av disse er 80% vellykkede. Årsaker til mislykket nevral rørtransplantasjon inkluderer død av embryoet på grunn av vevsskade som oppstår under mikrokirurgi, eller svikt i nevralrøret for å integrere seg i vertsembryoet. Sistnevnte kan skyldes dårlig plassering av nevralrøret i verten eller fra et nevralrør av dårlig kvalitet på grunn av dårlig avhandlingsteknikk eller fra overdreven eksponering for dissosiasjonsenzym. Det første screeningtrinnet, samt lignende senere undersøkelser for GFP+-celler, er nyttig, da det betyr at tid og ressurser ikke kastes bort ved å utføre eksperimenter på embryoer som ikke har GFP-merket NCC i tarmen.
Figur 3 viser prosedyren for DiI-injeksjon av blodårene. Effektiviteten /suksessen til DiI-injeksjonsteknikken avhenger av: Først kutte injeksjonsnålen til optimal diameter for den målrettede venen, andre en presis gest når du setter nålen i venen (for ikke å trenge gjennom den andre siden), og for det tredje unngå at nålen blir plugget under injeksjonen ved å blåse med konstant hastighet. Hvis noen av disse tre parametrene gjøres feil, vil embryoet blø ut eller trenger flere timer for å gjenopprette før et nytt forsøk blir gjort, da blødningen gjør det nesten umulig å injisere umiddelbart. Vellykkede embryoer bør velges umiddelbart ved å se under et stereofluorescensmikroskop og må dissekeres raskt. I vellykkede embryoer er DiI-merkede blodkar til stede i hele embryoet (Figur 3C, D) inkludert kapillære senger (Figur 3D).
Ved høsting av embryoer og undersøkelse av vevsseksjoner eller fullkomne mage-tarmkanaler, typiske resultater avslører GFP+ NCC innenfor den primitive ENS og den fine strukturen til DiI-merkede tarmblodkarnettverk (Figur 4) Wholemount-preparater kan undersøkes ved hjelp av konfokal mikroskopi der bildestakker produserer tredimensjonale (3D) rekonstruksjoner som viser sammenhengene mellom de fine projeksjonene av GFP + ENS-celler og DiI-farget vaskulært system (Figur 4 A-C; G-I; Videoer 1 og 2).

Figur 1. Anbefalte mikrokirurgiinstrumenter. (A) mikro-skalpell formet fra en synål. (B) fin Arkansas stein for å forme en mikro-skalpell. (C) a) rett saks, b) buet saks, c) 5 ml sprøyte med 181/2 G hypodermisk nål, d) plastpipette, e) spesiallaget eggholder, f) svart blekk, g) firkantet klokkeglass, h) firkantet klokkeglass med svart sylgard base, i) mikro-skalpell på nåleholder, j) minutien pinner, k) minutien eller wolfsten nål på nålholder, l) Pascheff-Wolff vår saks, m) Dumont #5 pinsett, n) perforert skje, oi) kort brann-trukket overføring nål, oii) lang brann-trukket inking, p) munn. Klikk her for å se en større versjon av denne figuren.

Figur 2. Intraspecies nevral rørtransplantasjon. Chick embryo / GFP neural tube bilder har blitt modifisert fra Delalande et al. 12. Vaskularisering er ikke nødvendig for tarmkolonisering ved enteriske nevrale kamceller. Klikk her for å se en større versjon av denne figuren.

Figur 3. Intravenøs DiI-injeksjon. (A) Anbefalte instrumenter: a) CellTracker CM-DiI-dråpe på parafilm, b) trukket glassinjeksjonsnål, c) munnrør. (B) Skjematisk diagram over intravenøs DiI-injeksjon i E4 chimerisk kyllingembryo. (C) i ovo DiI intravenøs injeksjon som viser fin glassnål som inneholder DiI satt inn i venen (pil). (D) E4 chimerisk embryo post DiI injeksjon (rød) med GFP + nevralrør (pil). (E) DiI farget fint blodkar nettverk i et levende embryo, 24 timers etter injeksjon. Br: hjerne; H: hjerte; LB: lem knopp; A: allantois. Bilder i (C) og (D) er endret fra Delalande et al. 12 Vaskularisering er ikke nødvendig for tarmkolonisering av enteriske nevrale kamceller. Klikk her for å se en større versjon av denne figuren.

Figur 4: Representative resultater i magen og caecum av en E5.5 chick embryo. (A-C) 3 dimensjonal (3D) rekonstruksjon av en konfokal bildestabel i mageområdet som viser (D) GFP + enteriske nevrale kamceller (ENCC) (E) DiI-farget vaskulært system og (F)) et sammenslått bilde av begge nettverkene D-F Histologiske seksjoner på nivået av magen som viser (G) GFP + ENCC (H) DiI farget vaskulært system og (I) et fusjonert bilde av begge nettverkene. Nuclei er farget med DAPI (cyan). (G-H) 3D-rekonstruksjon av en konfokal bildestakk i caecum-regionen som viser (A) GFP+ ENCC-migreringsfronten i grønt, (B) DiI-farget vaskulært system i rødt, og (C) et sammenslått bilde av begge nettverkene. Bilder (A-F) er endret fra Delalande et al. 12 Vaskularisering er ikke nødvendig for tarmkolonisering av enteriske nevrale kamceller. Klikk her for å se en større versjon av denne figuren.

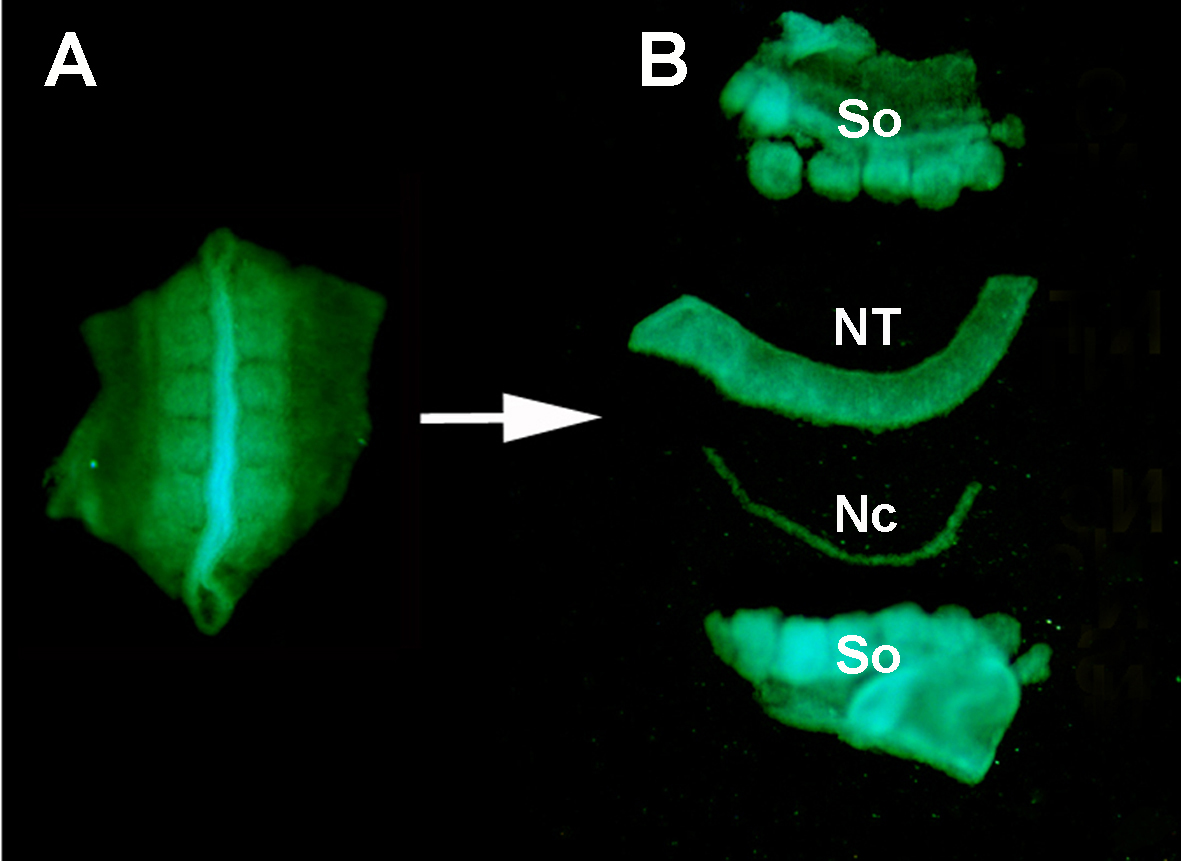
Figur S1. Isolering av en donor GFP + nevralrør fra det omkringliggende vevet ved enzymatisk fordøyelse og mikro-disseksjon. (A) GFP+ nevralrør og tilstøtende somitter dissekert fra donorembryoet. (B) Isolert nevralt rør etter pankreatin fordøyelse og mikro-disseksjon ved hjelp av rustfrie minutien pinner. Så: somitter; NT: nevralt rør; Nc: Notokord.

Video 1. 3-dimensjonal 360° rotasjon av bildet i figur 4C, som viser det vaskulære systemet og ENCC i magen på E5,5 (HH27-28). Klikk her for å se denne videoen.

Video 2. 3-dimensjonal 360° rotasjon av bildet i figur 4I, som viser det vaskulære systemet og ENCC-migrasjonsfronten i caecumområdet på E5.5 (HH27-28). Klikk her for å se denne videoen.
Discussion
Metoden for intraspecies neural tube podning, kombinert med blodkarmerking beskrevet her, utnytter den enkle tilgangen til fugleembryoet i egget (sammenlignet med andre virveldyrembryoer) for å studere samutviklingen av et element i det autonome nervesystemet (ENS) og det vaskulære systemet.
For merking av NCC-derivater har kyllingenGFP-chick intraspecies podningsmetoden vi beskriver en rekke fordeler i forhold til den klassiske vaktelkylling chimera-metoden som ble etablert for over 40 år siden1-3. For det første, under FITC-lys, er GFP-fluorescens ekstremt lys, i den grad at GFP + celler er lett merkbare i levende chimeriske embryoer. Dette gjør at podatets suksess kan kontrolleres i ovo, mens vaktel-kylling podning krever at embryoet blir drept, behandlet og immunostert ved hjelp av QCPN, før graftets suksess kan fastslås2. For det andre er GFP-uttrykket i den transgene kyllingenGFP cytoplasmisk, derfor merker det ikke bare cellelegemer, men gjør det også mulig å visualisere fremspringene til de transplanterte cellene22. Dette tillater at intrikate nevronnettverk observeres ved høy oppløsning (merk at fine projeksjoner best visualiseres når prøven er immunostained med anti-GFP antistoff). Siden QCPN-merking er begrenset til vaktelcellekjernen, avsløres ikke slike nettverk ved hjelp av vaktel-kylling chimeras. For det tredje eliminerer intraspecies podning eventuelle artsforskjeller mellom celler i det chimeriske embryoet. Siden vaktelegg embryoer har en kortere inkubasjonsperiode enn kylling (19 dager versus 21 dager) har det blitt antydet at vaktelceller har en høyere spredningshastighet enn kyllingceller, noe som potensielt kan påvirke utviklingen av det chimeriske vevet23. Interessant nok har det også blitt vist i planter at interspecies podning kan produsere omfattende endringer i DNA-metyleringsmønstre i verten 24. For det fjerde legger chickGFP til rette for ryggtransplantasjonseksperimenter for å ta opp emner som NCC-skjebne og celleforpliktelse25. For det femte er den transgenekyllingen GFP også nyttig for mange andre teknikker, inkludert FACS-sortering av GFP + celle subpopulations, organotypisk kultur av organer som inneholder GFP + celler, genetisk manipulering av GFP + podet vev via elektroporasjon av uttrykk plasmider26, og andre bildeteknologier som optisk projeksjon tomografi27.
Neural tube transplantasjon tilnærming kan endres ved mikrokirurgisk erstatte kortere mengder neural tube. Ved å bruke mindre segmenter av nevralrør er mikrokirurgien potensielt mindre skadelig for embryoet, og overlevelsen kan forbedres. Ulempen med å transplantere mindre nevralt rør er imidlertid at antallet GFP + NCC i verten vil bli redusert. Brukere kan prøve å oppnå en balanse mellom mengden nevralrør transplantert for å gi optimal overlevelse av embryoer, og antall GFP + NCC innen vertstarmen tilstrekkelig til å gi informative resultater.
For fartøymaling har DiI fordelen at fluorescensen er veldig lys og robust. Den har også kapasitet til å spre seg under fiksering som forsikrer farging av de fineste åpnede kapillærene. Siden det er et viktig fargestoff, kan embryoer overleve injeksjonsprosedyren og fortsette å utvikle seg med et farget vaskulært system (opptil 24 timer i våre hender, selv om fargingen blir mer punktlig over tid, se figur 3E). Kombinasjonen av kyllingGFP-podning med DiI vaskulært maleri er derfor kompatibel med levende avbildning. Foruten alle disse fordelene, er det viktig å merke seg at vaskulær injeksjon bare merker luminiserte kar og derfor ikke identifiserer uåpnede kapillærer, endotelceller eller isolerte endotelceller. Videre fremgang i fugletransgenese kan imidlertid gi nye måter å omgå slike problemer på, som eksemplifisert ved eksperimenter ved hjelp av Tg(tie1:H2B-eYFP) vaktelembryoer for å studere vaskulær morfogenese28. En annen begrensning ved denne teknikken er at for effektiv fartøymerking i embryoer på E7,5 og utover, må større mengder fargestoff injiseres, noe som kan gjøre eksperimenter dyre. En modifikasjon av teknikken kan imidlertid omfatte lavpris blodkar merkingusing highlighter blekk14, selv om denne tilnærmingen ikke har blitt prøvd i våre hender.
Kritiske trinn i prosedyrene inkluderer prosessen med å visualisere embryoet ved å injisere blekk under blastodisc. Hvis membranen som dekker eggeplommen blir revet av den blekkfylte nålen på dette stadiet, blir embryooverlevelse alvorlig kompromittert. Det er også viktig, når du forbereder en donor nevral tube, at vevet ikke er igjen i altfor lang tid i bukspyttkjertelen (vurder ca. 10 min som maksimum). Langvarig eksponering for pankreatin skader vevet og nevralrøret er da vanskelig å håndtere, og det vil ikke innlemme godt i verten. Å få erfaring med DiI-injeksjonsteknikken på embryoer av vill type er viktig før injisering av chimeriske embryoer, da bare ett forsøk på injeksjon generelt er mulig for hvert embryo. DiI-volum og nåleediameter er kritiske parametere for hvert embryo og bør vurderes på vill type, trinntilpassede kontroller.
Til slutt kan vår doble merkingsmetode for nevral rørtransplantasjon og DiI-karmaling i levende kyllingembryoer brukes til å undersøke sammenhengene mellom NCC og blodkarnettverk under organogenese. Tatt i betraktning mekanismene som er ansvarlige for å etablere riktig mål innervering og vaskularisering under organutvikling, er fortsatt stort sett ukjent, har denne metodikken potensial for fremtidige funn på dette feltet.
Disclosures
Forfatterne erklærer at de ikke har konkurrerende økonomiske interesser.
Acknowledgments
Befruktede GFP kyllingegg ble levert av prof. Helen Sang, The Roslin Institute og University of Edinburgh, UK. Roslin Transgenic Chicken Facility er finansiert av Wellcome Trust og av Biotechnology and Biological Sciences Research Council (BBSRC). Arbeidet ble delvis finansiert, og NT støttet, av Great Ormond Street Hospital Children's Charity, London, UK. Forfatterne takker Ben Jevans, UCL Institute of Child Health, for hjelp til å forberede embryoer for podning.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}