

Summary
这是一种使用宿主范围选择和重组病毒的可视化识别来生成"无疤痕"重组病毒的方法。
Abstract
Vaccinia病毒(VACV)有助于从自然中根除天花的病原体变异病毒(VARV)。VACV自首次作为疫苗使用以来,一直作为治疗性疫苗的载体和溶血性病毒而开发。这些应用利用 VACV 易于操作的基因组和广泛的主机范围,作为一个出色的平台,以生成具有各种治疗应用的重组病毒。已经开发了几种方法来生成重组VACV,包括标记选择方法和瞬态显性选择。在这里,我们介绍了主机范围选择方法的改进,以及重组病毒的可视化识别。我们的方法利用宿主抗病毒蛋白激酶R(PKR)产生的选择性压力,加上荧光融合基因,表达mCherry标记的E3L,这是两个VACV PKR拮抗剂之一。盒式磁带,包括感兴趣的基因和mCherry-E3L融合,由从VACV基因组派生的序列组成。感兴趣的基因和mCherry-E3L是一个较小的区域,与3'臂的前150个核苷酸相同,促进mCherry-E3L基因在选择后的同源重组和丢失。我们证明,这种方法允许在各种细胞类型中高效、无缝地生成 rVACV,而无需选择药物或广泛筛选突变病毒。
Introduction
Vaccinia病毒(VACV)有助于首次成功地从自然界中消灭人类病原体变异病毒(VARV)。自从病毒灭绝以来,包括VACV在内的痘病毒一直为人类和动物医学提供有用的治疗病毒。例如,基于VACV的狂犬病病毒疫苗在防止神经病性狂犬病在欧洲1和美国2的传播方面非常有效。最近,反应各种抗肿瘤分子(如单链抗体或人类红细胞生成素)的重组痘病毒作为溶血剂33、4、54,5取得了令人鼓舞的成功。VACV作为载体特别有吸引力,因为它容易受遗传操作,具有广泛的宿主范围,在各种条件下保持稳定,便于运输和疫苗在6、77领域的生存能力。虽然已经开发出多种技术来生成用于实验室实验和疫苗生成的重组VACV,但目前产生这些病毒的策略有显著的局限性。
由于VACV的效用,开发了多种生成重组病毒的策略。第一种策略采用同源重组来引入一种盒式磁带,包括转基因和可选标记基因,如抗生素耐药性基因。盒式磁带的两个+500核苷酸(nt)或更大的手臂将基因定向到病毒基因组中的特定部位,然后通过双交叉事件88、9、109,10进行稳稳地集成。这一战略是快速和高效的;然而,它产生额外的遗传物质的形式标记基因,可能产生意想不到的效果。此外,转基因数量有实际的上限,可以引入的转基因标记数量受可用独特可选择标记的数量限制。瞬时主导选择(TDS)策略通过促进"无痕"重组病毒的生成解决了这一问题。使用这种策略,包含突变VACV基因和可选择标记基因的质粒被集成到病毒基因组中,但没有额外的侧翼VACVDNA。这种方法导致整个质粒的暂时整合和VACV基因的重复,由于单个交叉事件的整合。这种中间体是稳定的,只要它在选择压力下保持,允许此构造的丰富。移除选择后,VACV 复制启用第二个交叉事件,导致以大约 50:50 的比例去除质粒并随后形成野生类型 (wt) 或重组病毒。虽然TDS生成重组病毒而无需稳定引入外来DNA,但多个病毒克隆必须通过测序分析筛选出预期的突变,这一步骤可能耗时且成本高昂。
在这里,我们提出了一种生成重组痘病毒的方法,结合每种方法的最佳方面,类似于为复制无能的、经过修改的安卡拉12、13、14,14所述的方法。12,此策略结合了视觉和宿主范围选择,通过双交叉事件快速生成重组病毒,然后通过同源重组消除可选标记基因。这种方法允许由同源重组促进的突变体的快速生成,具有TDS方法的"无痕"性质,同时无需随后的筛选步骤来区分野生病毒和突变病毒。我们的方法还采用宿主范围选择代替抗生素选择,消除了细胞系中化学诱导的型比变化的风险。对于这种方法,我们选择使用宿主抗病毒蛋白激酶R(PKR)作为选择性剂,以产生重组VACV。在大多数单元格类型15中,PKR 表示为非活动单体。在N-终端dsRNA结合域中结合双链RNA(dsRNA)时,PKR变暗,并自磷化16。这种活性形式的PKR磷酸酶酶启动因子2(eIF2)的α亚单位,最终抑制启动器二甲酰-tRNA的传递,从而防止细胞内翻译,并广泛抑制许多病毒家族17,18,18的复制。
为了应对PKR广泛而有效的抗病毒活性,许多病毒至少进化了一种防止PKR激活的策略。大多数痘病毒表达两个PKR拮抗剂,由VACV中的E3L和K3L基因编码,通过两种不同的机制19对抗PKR。E3通过结合双链RNA20,2121来防止20PKR消增,而K3则通过直接结合到激活的PKR,从而抑制与其基质eIF2+22的相互作用,作为伪22基质抑制剂。重要的是,这两个PKR拮抗剂不一定能抑制所有物种的PKR。例如,羊痘病毒的K3同源性强抑制羊的PKR,而羊痘E3同源性没有表现出相当大的PKR抑制23,24。23,在这项研究中,我们提出了一种使用PKR介导选择性压力与荧光选择相结合的方法,为E3L和K3L(VC-R4)生成一种VACV重组,这种重组不能在从不同物种衍生的PKR合格细胞中复制。这种重组病毒为快速生成重组病毒提供了极好的背景,这些病毒表达由原生E3L启动器控制的基因。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 生成重组矢量
- 设计引漆以生成选择盒。使用多个在线引物设计工具,使用多个在线引物设计工具中的任何一种具有重叠序列和矢量设计每个具有重叠序列的单个放大器,以方便 DNA 分子的等温酶组装,也称为 Gibson 组装。
注:此协议也可以使用传统的基于限制内核酶的克隆方法完成。在这种情况下,使用适当的限制位点而不是重叠序列设计引素。 - 使用步骤 1.1 中设计的引素,PCR 按 5' 到 3' 的顺序放大以下元素(图 1): E3L (5' 臂)的 VACV 基因组区域 5' 的 +500 核苷酸、 EGFP 或感兴趣的基因, #150核苷酸从VACV基因组区域立即3'E3L(短3'手臂),合成早期/晚痘病毒启动剂25,mCherry-E3L融合基因,和#500核苷酸从VACV基因组区域3'E3L包括短3'手臂(长3'手臂)。25
- 在 PCR 管中,按以下顺序添加试剂,用于每种安培:17 μL 的 DNase 自由水, 每个底漆的1.2μL(初始浓度=10μM,最终浓度=0.5μM),5μL 5x PCR反应缓冲液的5μL,模板DNA(从质粒放大的安培体10纳克:EGFP和E/L促进剂-mCherry-E3L盒;100纳克,从病毒基因组DNA扩增的amplicons:5'和3'手臂),以及0.5L的DNA聚合体。调整最终反应体积为 50 μL 的加入水量。
注:模板DNA的浓度应该根据经验确定,但我们一般从10纳克/反应开始。 - 将管放入热循环器中,在98°C下熔化DNA30秒,然后使用25轮三步PCR协议:98°C为5s,55°C为10s,72°C为1分钟。
注:根据制造商建议的 Tm 为每个底漆集确定熔融温度。根据每个安培的长度(1 分钟/kb)确定适当的延长时间。
- 在 PCR 管中,按以下顺序添加试剂,用于每种安培:17 μL 的 DNase 自由水, 每个底漆的1.2μL(初始浓度=10μM,最终浓度=0.5μM),5μL 5x PCR反应缓冲液的5μL,模板DNA(从质粒放大的安培体10纳克:EGFP和E/L促进剂-mCherry-E3L盒;100纳克,从病毒基因组DNA扩增的amplicons:5'和3'手臂),以及0.5L的DNA聚合体。调整最终反应体积为 50 μL 的加入水量。
- 在 1% 的琼脂胶凝胶上可视化扩增产物。在每个井中加入每个DNA产物的10μL和2 μL的加载缓冲液,以8 V/cm的速度运行1小时。
- 凝胶使用DNA凝胶提取试剂盒和制造商协议纯化每个安培。通过添加 50 μL 的 DNase 自由水并立即离心,从柱中释放安培。
- 使用EcoRI内酶酶消化线性化 pUC19 克隆载体。在管中,加入1μg的pUC19,将水加入17μL的体积,2升反应缓冲液,1 μL(20个单位)的EcoRI。在37°C孵育1小时。
- 在 1% 的琼脂胶凝胶上可视化放大产品,在 8 V/cm 下运行 1 小时。从凝胶中去除带,并使用第 1.4 步所述的 DNA 凝胶提取试剂盒纯化产品。
- 使用主混合套件,将所有个体、凝胶纯化的放大器和线性矢量进行装。
- 到 PCR 管中,加入 0.2 pmol 的线性化 pUC19 和每个安培(5' 臂,EGFP,短 3' 臂,E/L 促进器-mCherry-E3L 盒,3'臂)。将无D纳塞游水加入到10μL的最终体积中,然后加入10μL的DNA组装主混合物。在50°C下孵育样品1小时。
- 从步骤1.6开始,将化学合产大肠杆菌从步骤1.6中转化,如前所述的26、27。,27含有100微克/mL安培霉素的LB琼脂糖板上转化细胞的板100 μL。在37°C下孵化板过夜。
- 选择分离良好的菌落,并将单个菌落转移到含有100微克/mL安培霉素的含有Luria肉汤的管子上。在37°C下孵育管过夜,同时在225 rpm时摇动。
- 使用质粒迷你制备试剂盒将质粒与过夜培养物分离。使用分光光度计检查 DNA 的浓度和纯度。A260/A280 比介于 1.8 和 2.0 之间是可以接受的。
- 提交桑格测序的质粒,以确定所需的克隆产品是否正确。将DNA储存在-20°C。
2. 生成重组病毒
- 感染适合细胞的汇合单层与病毒,在6孔板的1.0(MOI = 1.0)的多重感染中重新组合。在37°C和5%CO2下孵育受感染的细胞1小时。然后吸气介质并将其替换为新鲜的 DMEM。在37°C和5%CO2孵育受感染的2细胞。
注:对于复制能力病毒,如缺乏K3L22的气喘病毒,细胞系,如欧洲兔肾细胞系RK13(ATCC#CCL-37)或BSC-40是合适的。然而,对于复制缺陷病毒,如本文中描述的病毒,缺乏PKR拮抗器E3L和K3L,需要一种补充细胞系,在跨或PKR敲除或敲出细胞中表达这两个基因。 - 使用制造商协议在步骤 1.10 中生成和验证的 500 ng 载体的受感染细胞。在37°C和5%CO2孵育细胞48小时。
注:如果使用缺乏E3L和K3L的真空病毒,PKR介导的选择性压力将推动重新组合病毒的选择,并保持在这些细胞中表达mCherry-E3L融合蛋白。如果需要,也可以PCR放大仅用于转染的插入物,而不是整个质粒。 - 感染后48小时,收获感染的单层。在某些情况下,细胞可以通过移液来收获,但如果它们仍然紧密粘附,则用细胞刮刀收获它们。冷冻细胞三次,然后以50%的振幅将水相声15s。将这种酸乳酸储存在-80°C,直到随时使用。
- 连续10倍稀释步骤2.3中收获的水化水,从10-1到10-6,将120μL的水合物加入到1080μL的DMEM(10-1),然后将-1120μL的稀释物加到1080μL的DMEM(10-2),并重复这个过程4-2次。将每个稀释物的1 mL添加到PKR合格细胞系的单个汇合孔中,在这种情况下为RK13细胞。
- 在37°C和5%CO2下孵育受感染的细胞1小时。然后吸气介质,用新鲜的DMEM在37°C和5%CO2孵育受感染的细胞。
- 感染后24至48小时,通过荧光显微镜识别重组病毒。重组病毒的斑块由于mCherry-E3L融合基因的整合而表示红色荧光(图2)。如果最初使用不含PKR抑制剂的病毒,所有斑块将含有重组病毒。
- 斑块在RK13细胞上三次纯化重组病毒。最后一轮斑块净化后,所有斑块应表达红色荧光。
- 感染从步骤2.6开始用斑块纯化的红色荧光病毒感染RK13细胞的流6孔板,表达VACV PKR抑制剂E3L和K3L(RK13_E3L+K3L细胞28)。每口井可达到约50-100块斑块。
注:这些细胞提供VACV PKR拮抗剂在跨和缓解PKR介导的选择性压力,以保持mCherry-E3L融合基因,从而促进重组病毒的"无疤痕"生成。 - 使用 EVOS2 显微镜和 GFP 滤立方(激发:470/22,发射:525/50)和 RFP 滤波器立方体(激发:531/40,发射:593/40)通过荧光显微镜识别已崩溃的病毒。
注:mCherry-E3L融合基因丢失的频率约为2.5%(表2)。如果不将EGFP作为标记基因包括在内,失去mCherry-E3L融合基因的突变病毒的斑块将是无色的。 - 在 RK13_E3L+K3L 细胞上,斑块纯化绿色 (VC-R4) 或无色斑块 (E3L) 三次。确保没有斑块荧红。
- 确认 mCherry-E3L 的丢失以及 PCR 和桑格测序的预期突变的存在。
注:如果感兴趣的基因或突变没有PKR抑制活性,重组病毒必须在RK13_E3L+K3L细胞或等效的PKR抑制或PKR缺陷细胞系上生长(图3)。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
我们使用图 1中所示的过程生成了一个 VACV,该 VACV 缺少 PKR 拮抗器 E3L 和 K3L,在已为 K3L (vP872) 删除的病毒中用 EGFP 替换 E3L。图 2显示了 PKR 合格 RK13 细胞中指示 mCherry-E3L 病毒表达的红色荧光斑块,以及 RK13_E3L_K3L 细胞中的 EGFP,确认 E3L 的丢失和 mCherry-E3L 选择标记的崩溃。图3证实,这种重组病毒VC-R4,缺少两个PKR拮抗剂不能复制在PKR胜任的RK13细胞,而父病毒,vP872表示E3L,是复制能力。为了确认在RK13细胞中无法复制,只是因为E3L的丢失,我们用E3L替换了VC-R4中的EGFP,以便使用相同的选择协议生成还原病毒。图 3还验证此还原病毒复制的效率是否与 RK13 细胞中的 vP872 一样有效。有趣的是,在选择RK13_E3_K3细胞之前,与mCherry-E3L选择标记的折叠一致的无色斑块被识别,这些细胞通常需要选择"无疤痕"重组剂,这可能是因为mCherry-E3L重组盒和E3L基因插入VC-R4之间的序列识别扩展。因此,为了确定重组的效率,以及崩溃的速度,我们选择产生病毒表达痘病毒PKR拮抗剂K3L,以避免早期崩溃的问题23。图 4表示 RK13_E3L_K3L 细胞感染后无色斑块(箭头)的外观。表1显示了三个独立实验的结果,其中平均12.6%的祖生病毒与转染质粒进行了重组,类似于先前报告的频率29、30、31。29,30,31表 2详细介绍了 RK13_E3L_K3L 细胞中无色斑块相对于总斑块的频率,表明 mCherry-E3L 选择标记的折叠和损耗频率约为 1.8%。
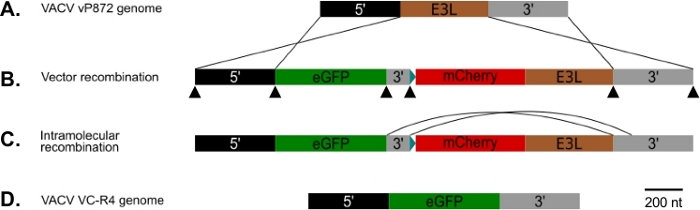
图 1:p837-GOI-mCherry-E3L 以及主机范围和视觉重组策略的图。(A) 5' 臂 (黑色) 和 3' 臂(灰色) 侧翼 E3L 轨迹(棕色) VACV 中。(B) 在p837-GOI-mCherry-E3L中,这些手臂侧翼一盒含有感兴趣的基因 (GOI),在本例中,EGFP(绿色)由一个短的 3' 手臂(灰色)从 mCherry-E3L(红色)融合基因的控制之下分离。25这些外部臂驱动 VACV 和 p837-GOI-mCherry-E3L 之间的同源重组。黑色箭头表示吉布森克隆用来生成这种质粒的重叠引素的位点。(C) 当去除PKR选择性压力时,可以选择在短臂和长3'臂之间进行分子内重组的病毒。(D) 导致病毒 (VC-R4) 只包含 E3L 位点感兴趣的基因。请点击此处查看此图形的较大版本。

图2:在重组病毒斑块重组24小时后(顶部)的荧光显微图,在RK13细胞中表达mCherry(左)和EGFP(右)。(下图)在RK13+细胞中去除PKR介导的选择性压力48小时后,重组病毒斑块的显微图,表示EGFP(右),但不表示mCherry(左)。刻度条表示所有面板的 650 μm。请点击此处查看此图形的较大版本。
图3:VC-R4不能在PKR能力单元中复制。指示的细胞系在MOI = 0.1感染了vP872(蓝色)、VC-R4(绿色)或VC-R4+E3L(洋红色)。感染后48小时,通过RK13_E3L+K3L细胞的连续稀释,采集和进行发泡。Titers 以 PFU/mL 报告,误差条表示三个复制实验的标准偏差。请点击此处查看此图形的较大版本。

图 4:RK13_E3L_K3L 细胞中 mCherry-E3L 表达式丢失。VC-R4_K3L-mCherry-E3L感染RK13_E3L+K3L细胞的荧光和相位对比显微图的叠加。由于产生 VC-R4_K3L 的选择盒的折叠,三块斑块不再表示 mCherry(圆圈)。请点击此处查看此图形的较大版本。
| 实验 1 | 实验2 | 实验3 | |
| 红色斑块 (RK13) | 30 | 11 | 18 |
| 牙菌斑总数(RK13_E3L+K3L) | 225 | 64 | 249 |
| 重组速率 | 13.30% | 17.20% | 7.20% |
表1:VACV与p837-K3L-mCherry-E3L质粒的重组频率。
| 实验 1 | 实验2 | 实验3 | |
| 牙菌斑总数(RK13_E3L+K3L) | 115 | 44 | 210 |
| 无色斑块(RK13_E3L+K3L) | 3 | 1 | 1 |
| 重组速率 | 2.60% | 2.30% | 0.50% |
表2:RK13_E3_K3电池中VC-R4_K3L-mCherry-E3L的mCherry-E3L损耗频率。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
在这里,我们提出了一个瞬态标记选择策略32的变化,以生成重组性气喘病毒,而不保留外来DNA在最后重组病毒。我们的策略使用由宿主抗病毒蛋白PKR介导的选择性压力,而不是其他形式的选择性压力,如抗生素。使用宿主抗病毒基因消除了细胞中化学诱导的型物变化的可能性,或由于选择药物而增加突变的风险。此外,与药物选择不同,我们的方法没有滞后阶段,因为PKR在所有细胞中都是有组织地表达的。基于 mCherry 表达的次视觉选择还提高了此方法的特异性,确保在第一阶段只选择表达转基因的斑块,在选择失去 mCherry-E3L 基因的成熟重组病毒时,作为负选择性标记同样有效。
此重组策略最关键的步骤是生成适当的重组载体,并适当进行斑块纯化,以确保所选病毒是克隆病毒。在本文中,我们建议使用"吉布森装配"来生成重组向量。此策略非常有效,允许在一天内组装包含重组向量的所有片段。然而,由于短3'臂和长3'手臂具有相同的序列,这些片段有可能在克隆反应期间连接在一起,有些载体可能不包含mCherry-E3L盒。根据我们的经验,这种情况很少见,但有必要确认克隆后载体的结构。我们还使用传统的内核酶和连接法为此策略生成了重组向量。此策略避免了上述问题,但可能更耗费人力。斑块纯化通常很简单,主要依赖于使用适当的放任细胞进行初始重组,PKR能力细胞进行初始斑块纯化,以确保只有重组病毒才能复制,然后再次放任细胞,以促进分子内重组和可选择标记的丢失。因此,密切注意细胞系对于成功实施和有效应用这一策略至关重要。
在这项研究中,我们演示了使用这种方法生成为PKR拮抗器E3L和K3L删除的VACV重组,并在E3L启动器的控制下表达EGFP。今后,这种病毒将作为未来重组病毒的有效背景,因为它无法在PKR能力细胞中复制。因此,将有强大的PKR介导的选择性压力,驱动mCherry-E3L重组盒成后代病毒,同时基本上防止复制非重组病毒。此外,通过接受重组盒而损失EGFP是一个有用的二次选择标记,以确保采摘的斑块不会与非重组病毒共同感染。我们观察到的重组速率与之前报告的 VACV 速率一致,但视觉荧光标记通过确保增加选择适当重组病毒的可能性来提高生成重组病毒的效率。我们在PKR能力细胞上经过两轮选择后,对无色斑块的观察,大概是由于E3L和mCherry-E3L标记基因之间相同序列长度的增加,表明mCherry-E3L的损耗率可以通过增加或减少3'短臂的长度来"调整"。该技术的主要局限性是使用PKR作为重组剂的选择性压力。这种重组策略的最有效用法是在缺乏PKR对抗因素的背景下生成这些病毒。然而,色度选择标记允许这种重组策略的使用,即使没有PKR的介导选择,只需通过斑块纯化mCherry表达斑块。虽然缺乏PKR介导的选择性压力将降低第一个筛选步骤的效率,但mCherry表达斑块的百分比仍然很高,以至于基于颜色的选择是可行的。因此,这种方法可用于将几乎任何基因插入痘病毒基因组中。
正如EGFP的插入所证明的那样,通过这种方法,任何基因都可以迅速插入由原生启动子控制的E3L位点中,前提是如果转基因不是PKR拮抗剂,则PKR空细胞或补充细胞系用于下游实验。这一策略,结合我们在这里报告的VC-R4病毒,增加了一种新的和强有力的方法,使用宿主介导的选择性压力和早期重组剂的视觉识别,快速可靠地生成重组性空腹病毒。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者声明没有相互竞争的财务利益。
Acknowledgments
该项目由国家卫生研究院(AI114851)资助至SR。
Materials
| Name | Company | Catalog Number | Comments |
| 2X-Q5 Master Mix | NEB | M0492L | High-fidelity polymerase used in PCR |
| Ampicillin | ThermoFisher Scientific | 11593027 | Bacterial selective agent |
| Disposable Cell Scrapers | ThermoFisher Scientific | 08-100-242 | Cell scraper to harvest infected cells |
| EVOS FL Auto 2 Cell imaging system | ThermoFisher Scientific | AMAFD2000 | Fluorescent microscope |
| EVOS Light Cube, GFP | ThermoFisher | AMEP4651 | GFP Cube |
| EVOS Light Cube, RFP | ThermoFisher | AMEP4652 | RFP Cube |
| GenJet | SignaGen Laboratories | SL100489 | Transfection reagent |
| Luria Bertani (LB) Broth | Gibco | 10855021 | Bacterial growth medium |
| Monarch DNA gel extraction kit | NEB | T1020L | Gel purification kit used to purify amplicons and linearized vectors |
| Monarch Plasmid Miniprep kit | NEB | T1010L | Miniprep kit ussed to purify plasmids |
| NanoDrop One | ThermoFisher Scientific | ND-ONE-W | Spectrophotometer used to measure RNA and DNA concentration |
| NEBuilder Master Mix | NEB | E2621L | Isothermal enzymatic assembly kit used to generate the recombination vector |
| Q500 Sonicator | Qsonica | Q500-110 | Sonicator for virus lysates |
| RK13 cells | ATCC | CCL-37 | Rabbit kidney cells |
| VWR Multiwell Cell Culture plates | VWR | 10062-892 | Cell culture plates |
References
- Brochier, B., et al. Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Nature. 354 (6354), 520-522 (1991).
- Pastoret, P. P., Brochier, B. The development and use of a vaccinia-rabies recombinant oral vaccine for the control of wildlife rabies; a link between Jenner and Pasteur. Epidemiology and Infection. 116 (3), 235-240 (1996).
- Chan, W. M., McFadden, G.
- Nguyen, D. H., et al. Vaccinia virus-mediated expression of human erythropoietin in tumors enhances virotherapy and alleviates cancer-related anemia in mice. Molecular Therapy. 21 (11), 2054-2062 (2013).
- Frentzen, A., et al. Anti-VEGF single-chain antibody GLAF-1 encoded by oncolytic vaccinia virus significantly enhances antitumor therapy. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12915-12920 (2009).
- Pastoret, P. P., Vanderplasschen, A.
- COLLIER, L. H. The development of a stable smallpox vaccine. The Journal of Hygiene. 53 (1), 76-101 (1955).
- Weir, J. P., Bajszár, G., Moss, B. Mapping of the vaccinia virus thymidine kinase gene by marker rescue and by cell-free translation of selected mRNA. Proceedings of the National Academy of Sciences of the United States of America. 79 (4), 1210-1214 (1982).
- Mackett, M., Smith, G. L., Moss, B. Vaccinia virus: a selectable eukaryotic cloning and expression vector. Proceedings of the National Academy of Sciences of the United States of America. 79 (23), 7415-7419 (1982).
- Nakano, E., Panicali, D., Paoletti, E. Molecular genetics of vaccinia virus: demonstration of marker rescue. Proceedings of the National Academy of Sciences of the United States of America. 79 (5), 1593-1596 (1982).
- Falkner, F. G., Moss, B. Transient dominant selection of recombinant vaccinia viruses. Journal of Virology. 64 (6), 3108-3111 (1990).
- Staib, C., Drexler, I., Ohlmann, M., Wintersperger, S., Erfle, V., Sutter, G. Transient Host Range Selection for Genetic Engineering of Modified Vaccinia Virus Ankara. BioTechniques. 28 (6), 1137-1148 (2000).
- Staib, C., Drexler, I., Sutter, G. Construction and Isolation of Recombinant MVA. Vaccinia Virus and Poxvirology. , 77-99 (2004).
- Di Lullo, G., et al. Marker gene swapping facilitates recombinant Modified Vaccinia Virus Ankara production by host-range selection. Journal of Virological Methods. 156 (1-2), 37-43 (2009).
- Pfaller, C. K., Li, Z., George, C. X., Samuel, C. E. Protein kinase PKR and RNA adenosine deaminase ADAR1: New roles for old players as modulators of the interferon response. Current Opinion in Immunology. 23 (5), 573-582 (2011).
- Bevilacqua, P. C., George, C. X., Samuel, C. E., Cech, T. R. Binding of the protein kinase PKR to RNAs with secondary structure defects: Role of the tandem A - G mismatch and noncontigous helixes. Biochemistry. 37 (18), 6303-6316 (1998).
- Krishnamoorthy, T., Pavitt, G. D., Zhang, F., Dever, T. E., Hinnebusch, A. G. Tight Binding of the Phosphorylated Subunit of Initiation Factor 2 (eIF2) to the Regulatory Subunits of Guanine Nucleotide Exchange Factor eIF2B Is Required for Inhibition of Translation Initiation. Molecular and Cellular Biology. 21 (15), 5018-5030 (2001).
- Rothenburg, S., Georgiadis, M. M., Wek, R. C. Evolution of eIF2α kinases: Adapting translational control to diverse stresses. Evolution of the Protein Synthesis Machinery and Its Regulation. , 235-260 (2016).
- Bratke, K. A., McLysaght, A., Rothenburg, S. A survey of host range genes in poxvirus genomes. Infection, Genetics and Evolution. 14, 406-425 (2013).
- Chang, H. W., Watson, J. C., Jacobs, B. L. The E3L gene of vaccinia virus encodes an inhibitor of the interferon-induced, double-stranded RNA-dependent protein kinase. Proceedings of the National Academy of Sciences. 89 (11), 4825-4829 (1992).
- Romano, P. R., et al. Inhibition of double-stranded RNA-dependent protein kinase PKR by vaccinia virus E3: role of complex formation and the E3 N-terminal domain. Molecular and Cellular Biology. 18 (12), 7304-7316 (1998).
- Beattie, E., Tartaglia, J., Paoletti, E. Vaccinia virus-encoded eIF-2 alpha homolog abrogates the antiviral effect of interferon. Virology. 183 (1), 419-422 (1991).
- Park, C., Peng, C., Brennan, G., Rothenburg, S. Species-specific inhibition of antiviral protein kinase R by capripoxviruses and vaccinia virus. Annals of the New York Academy of Sciences. 1438 (1), 18-29 (2019).
- Rothenburg, S., Brennan, G. Species-Specific Host-Virus Interactions: Implications for Viral Host Range and Virulence. Trends in Microbiology. , (2019).
- Chakrabarti, S., Sisler, J. R., Moss, B. Compact, synthetic, vaccinia virus early/late promoter for protein expression. BioTechniques. 23 (6), 1094-1097 (1997).
- Chung, C. T., Niemela, S. L., Miller, R. H. One-step preparation of competent Escherichia coli: Transformation and storage of bacterial cells in the same solution (recombinant DNA). Biochemistry. 86, 2172-2175 (1989).
- Chung, C. T., Miller, R. H. Preparation and storage of competent Escherichia coli cells. Methods in Enzymology. 218, 621-627 (1993).
- Rahman, M. M., Liu, J., Chan, W. M., Rothenburg, S., McFadden, G. Myxoma Virus Protein M029 Is a Dual Function Immunomodulator that Inhibits PKR and Also Conscripts RHA/DHX9 to Promote Expanded Host Tropism and Viral Replication. PLOS Pathogens. 9 (7), 1003465 (2013).
- Evans, D. H., Stuart, D., McFadden, G. High levels of genetic recombination among cotransfected plasmid DNAs in poxvirus-infected mammalian cells. Journal of Virology. 62 (2), 367-375 (1988).
- Ball, L. A. High-frequency homologous recombination in vaccinia virus DNA. Journal of Virology. 61 (6), 1788-1795 (1987).
- Spyropoulos, D. D., Roberts, B. E., Panicali, D. L., Cohen, L. K. Delineation of the viral products of recombination in vaccinia virus-infected cells. Journal of Virology. 62 (3), 1046-1054 (1988).
- Liu, L., et al. Transient dominant host-range selection using Chinese hamster ovary cells to generate marker-free recombinant viral vectors from vaccinia virus. BioTechniques. 62 (4), 183-187 (2017).
