

Summary
Bu konak aralığı seçimi ve rekombinant virüslerin görsel kimlik kullanarak "scarless" rekombinant vaccinia virüsler oluşturmak için bir yöntemdir.
Abstract
Vaccinia virüsü (VACV) variola virüsünün (VARV) kökünün kazınmasına yardımcı oldu. Vacv, aşı olarak ilk kullanımından bu yana terapötik aşılar için vektör ve onkolitik virüs olarak geliştirilmiştir. Bu uygulamalar, VACV'un kolayca manipüle edilen genomundan ve çeşitli terapötik uygulamalarla rekombinant virüsler oluşturmak için olağanüstü bir platform olarak geniş ana bilgisayar aralığından yararlanır. Marker seçimi yöntemleri ve geçici baskın seçki dahil olmak üzere rekombinant VACV oluşturmak için çeşitli yöntemler geliştirilmiştir. Burada, rekombinant virüslerin görsel tanımlaması ile birleştiğinde bir ana dizi seçim yöntemi nin bir iyileştirmesi salıyoruz. Yöntemimiz, iki VACV PKR antagonistinden biri olan mCherry etiketli E3L'yi ifade eden floresan füzyon geni ile birleşen konak antiviral protein kiazaz R (PKR) tarafından oluşturulan seçici basınçtan yararlanır. Kaset, ilgi geni ve mCherry-E3L füzyon uvacv genomundan elde edilen diziler ile çevrilidir. İlgi geni ve mCherry-E3L arasında homolog rekombinasyon ve seçim den sonra mCherry-E3L geni kaybıteşvik etmek için, 3 'kol ilk ~ 150 nükleotitler aynı daha küçük bir bölgedir. Bu yöntemin, mutant virüsler için ilaç seçimi veya kapsamlı tarama gerektirmeden çeşitli hücre tiplerinde verimli ve sorunsuz rVACV üretimine izin verdiğini gösteriyoruz.
Introduction
Vaccinia virüsü (VACV) bir insan patojeni ilk başarılı eradikasyon için aracı oldu, variola virüsü (VARV), doğadan. Variola virüsünün yok edilmesinden bu yana, VACV dahil olmak üzere poxviruses hem insan hem de hayvan tıbbı için yararlı terapötik virüsler olmaya devam edilmiştir. Örneğin, bir VACV tabanlı kuduz virüsü aşısıAvrupa'da sylvatic kuduz bulaşmasını önlemede çok etkili olmuştur 1 ve Amerika Birleşik Devletleri2. Daha yakın zamanda, anti-tümör molekülleri çeşitli ifade rekombinant poxviruses (örneğin, tek zincirli antikorlar veya insan eritropoietin) onkolitik ajanlar olarak teşvik başarı gördük3,4,5. VACV genetik manipülasyon için kolayca münasip olduğu için bir vektör olarak özellikle çekici, geniş bir konak aralığına sahiptir, ve çeşitli koşullar altında istikrarlı, alanında kolay ulaşım ve aşı canlılığı sağlayan6,7. Laboratuvar deneyleri ve aşı üretimi için rekombinant VACV üretmek için birden fazla teknik geliştirilmiş olsa da, bu virüsleri üretmek için mevcut stratejiler önemli sınırlamalar vardır.
VACV'un yararı nedeniyle, rekombinant virüsler oluşturmak için birden fazla strateji geliştirilmiştir. İlk strateji, transgen ve antibiyotik direnç geni gibi seçilebilir marker geni içeren bir kaset tanıtmak için homolog rekombinasyon kullanır. Kaset iki ~ 500 nükleotit (nt) veya daha büyük kollar viral genom, daha sonra stably çift crossover olaylar8,9,10ile entegre belirli bir siteye gen yönlendiren çevrilidir. Bu strateji hızlı ve verimlidir; ancak, beklenmeyen etkilere neden olabilir marker geni şeklinde ekstra genetik malzeme ile sonuçlanır. Ayrıca, mevcut benzersiz seçilebilir belirteçlerin sayısı ile sınırlı olarak tanıtılabilir transgen lerin sayısı için pratik bir üst sınır vardır. Geçici baskın seçilim (TDS) stratejileri "scarless" rekombinant virüslerin oluşumunu kolaylaştırarak bu sorunu ele almış11. Bu strateji kullanılarak, mutant VACV geni ve seçilebilir marker geni içeren bir plazmid viral genoma entegre edilmiştir, ancak vacv DNA'sını ekleyen bir kanaat olmadan. Bu yaklaşım, tek bir crossover olayı ile bütünleşme sonucunda VACV geninin tamamının geçici entegrasyonu ve çoğaltılması ile sonuçlanır. Bu ara, seçim baskısı altında muhafaza edildiği sürece stabildir ve bu yapının zenginleşmesine izin verilir. Seçim kaldırıldığında, VACV çoğaltma yaklaşık 50:50 oranında yabani tip (wt) veya rekombinant virüsün plazmid ve sonraki oluşumunun kaldırılması ile sonuçlanan ikinci bir crossover olay sağlar. TDS yabancı DNA'nın istikrarlı bir şekilde piyasaya sürülmesini gerektirmeden rekombinant virüsler üretirken, birden fazla virüs klonu, potansiyel olarak zaman alıcı ve maliyetli bir adım olan dizianalizi ile beklenen mutasyon için taranmalıdır.
Burada, bu yaklaşımların her birinin en iyi yönlerini birleştiren rekombinant poxviruses üreten bir yaklaşım savuruyoruz, replikasyon yetersiz modifiye vaccinia Ankara12için açıklanan bir yaklaşım a benzer12 ,13,14. Bu strateji, çift crossover olaylarla hızla rekombinant virüsler oluşturmak ve daha sonra homolog rekombinasyon ile seçilebilir marker genini ortadan kaldırmak için görsel ve ana bilgisayar aralığı seçimini birleştirir. Bu yaklaşım, homolog rekombinasyon aracılığı ile mutantların hızlı nesil izin verir, TDS yaklaşımlarının "scarless" doğası ile, vahşi tip ve mutant virüsleri ayırt etmek için bir sonraki tarama adımı gerektirmez iken. Yöntemimiz aynı zamanda antibiyotik seçimi yerine konak aralığı seçimi kullanır, hücre hattında kimyasal olarak indüklenen phenotik değişiklikler riskini ortadan kaldırarak. Bu yaklaşım için, rekombinant VACV oluşturmak için selektif ajan olarak konak antiviral protein kiril R (PKR) kullanmayı seçtik. PKR çoğu hücre türünde etkin olmayan bir monomer olarak ifade edilir15. N-terminal dsRNA bağlayıcı etki alanlarında çift iplikli RNA (dsRNA) bağlanan üzerine, PKR dimerizes ve otofosforile16. PKR fosforilatbu aktif formu ökaryotik inisiyasyon faktörü 2 (eIF2) alfa alt birimi, sonuçta ribozom için başlatıcı metiyonil-tRNA teslim inhibe, böylece hücre içi çeviri önlenmesi ve geniş birçok virüs ailelerinin çoğaltılmasını inhibe17,18.
PKR geniş ve güçlü antiviral aktiviteyanıt olarak, birçok virüs PKR aktivasyonu önlemek için en az bir strateji gelişti. Çoğu poxviruses iki PKR antagonistleri ifade, VACV E3L ve K3L genleri tarafından kodlanmış, hangi iki farklı mekanizmalar aracılığıyla PKR antagonize19. E3 çift iplikli RNA20bağlayarak PKR homodimerization önler,21, K3 doğrudan aktive PKR bağlanarak bir psödosubstrat inhibitörü olarak hareket ederken ve bu nedenle substrat eIF2α22ile etkileşimi inhibe . Daha da önemlisi, bu iki PKR antagonisti tüm türlerden PKR'yi mutlaka engellemez. Örneğin, koyun çiçeği virüsüK 3 homolog kuvvetle koyun PKR inhibe ederken, koyun E3 homolog önemli PKR inhibisyonu23,,24. Bu çalışmada, e3L ve K3L (VC-R4) için silinen bir VACV rekombinant oluşturmak için floresan seçimi ile birlikte PKR aracılı selektif basınç kullanmak için bir yöntem sürünüldü, hangi PKR yetkili hücrelerde farklı türlerden elde edilen çoğalamaz. Bu rekombinant virüs yerli E3L organizatörü kontrolü altında genleri ifade rekombinant virüslerin hızlı nesil için mükemmel bir arka plan sağlar.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Rekombinasyon vektörü oluşturma
- Seçim kasetini oluşturmak için astarlar tasarlayın. Birkaç online astar tasarım araçları herhangi birini kullanarak, aynı zamanda Gibson montaj olarak adlandırılan DNA moleküllerinin izomal enzimatik montaj kolaylaştırmak için komşu amplicons ve vektör ile örtüşen dizileri ile her bir amplicon tasarım.
NOT: Bu protokol geleneksel kısıtlama endonükleaz bazlı klonlama yöntemleri kullanılarak da tamamlanabilir. Bu durumda, çakışan diziler yerine uygun kısıtlama siteleri ile tasarım astarları. - 1.1 adımda tasarlanan astarları kullanarak, PCR aşağıdaki elementleri 5' ila 3'(Şekil 1):~500 e3L (5' kol), EGFP veya ilgi geninin VACV genomik bölgesi5' nükleotitleri sırasına göre yükseltin, ~150 nükleotit VACV genomik bölgeden hemen 3' E3L (kısa 3' kol), sentetik bir erken / geç poxvirüs organizatörü25,mCherry-E3L füzyon geni, ve ~ 500 nükleotitler VACV genomik bölge 3' kısa 3'kol (uzun 3' kol dahil olmak üzere VACV genomik bölge 3'.
- Bir PCR tüpünde, reaktifleri her amplicon için aşağıdaki sıraya ekleyin: 17 μL DNase serbest su, her astar1,2 μL (başlangıç konsantrasyonu = 10 μM, son konsantrasyon = 0,5 μM), 5 μL 5x PCR reaksiyon tamponu, şablon DNA (plazmidlerden güçlendirilmiş ampliconlar için 10 ng: EGFP ve E/L promotör-mCherry-E3L kaset; viral genomik DNA'dan güçlendirilmiş ampliconlar için 100 ng: 5' ve 3' kollar) ve 0,5 μL DNA polimeraz. Son reaksiyon hacmi 50°L olarak eklenen suyun hacmini ayarlayın.
NOT: Şablon DNA konsantrasyonu ampirik olarak belirlenmelidir, ancak genellikle 10 ng/reaksiyon la başlarız. - Tüpü bir termocycler yerleştirin ve DNA'yı 98 °C'de 30 s'de eritin ve ardından üç adımlı PCR protokolünün 25 mermisini kullanın: 5 s için 98 °C, 10 s için 55 °C ve 1 dk için 72 °C.
NOT: Her astar seti için üreticinin önerdiği Tm'ye göre erime sıcaklığını belirleyin. Her amplicon (1 dakika/kb) uzunluğuna göre uygun uzatma süresini belirleyin.
- Bir PCR tüpünde, reaktifleri her amplicon için aşağıdaki sıraya ekleyin: 17 μL DNase serbest su, her astar1,2 μL (başlangıç konsantrasyonu = 10 μM, son konsantrasyon = 0,5 μM), 5 μL 5x PCR reaksiyon tamponu, şablon DNA (plazmidlerden güçlendirilmiş ampliconlar için 10 ng: EGFP ve E/L promotör-mCherry-E3L kaset; viral genomik DNA'dan güçlendirilmiş ampliconlar için 100 ng: 5' ve 3' kollar) ve 0,5 μL DNA polimeraz. Son reaksiyon hacmi 50°L olarak eklenen suyun hacmini ayarlayın.
- Amplifikasyon ürünlerini %1 agarose jel üzerinde görselleştirin. Her bir DNA ürününden 10 μL ve her kuyuya 2 μL yükleme tamponu ekleyin ve 8 V/cm'de 1 saat çalıştırın.
- Jel bir DNA jeli çıkarma kiti ve üreticiprotokolü kullanarak her amplicon arındırMak. 50 μL DNase serbest su ve hemen santrifüj ekleyerek sütundan amplicons elute.
- EcoRI enonukleaz sindirimi kullanarak pUC19 klonlama vektörü doğrusallaştırın. Bir tüpe 1 μg pUC19, 17 μL hacimde su, 2 L reaksiyon tamponu ve 1 μL (20 birim) EcoRIekleyin. 37 °C'de 1 saat kuluçkaya yatırın.
- Amplifikasyon ürünlerini 8 V/cm'de %1 agarose jel üzerinde görselleştirin.
- Bir ana karışım kiti kullanarak tüm bireysel, jel saflaştırılmış amplicons ve doğrusalvektör ligate.
- Bir PCR tüpüne 0,2 pmol doğrusallaştırılmış pUC19 ve her amplicon (5' kol, EGFP, kısa 3' kol, E/L promotör-mCherry-E3L kaset, 3'kol) ekleyin. DNase serbest suyu 10 μL'lik son bir hacme ekleyin ve ardından 10 μL DNA montaj ana karışımı ekleyin. 50 °C'de 1 saat boyunca kuluçka örnekleri.
- Daha önce açıklandığı gibi adım 1.6 dan monte ürünün 2 μL ile kimyasal yetkili E. coli dönüştürün26,27. 100 μg/mL ampisilin içeren LB agarose plakaları üzerindeki dönüştürülmüş hücrelerin 100 μL plakası. Plakaları bir gecede 37 °C'de kuluçkaya yatırın.
- İyi izole edilmiş kolonileri seçin ve 100 μg/mL ampisilin içeren Luria suyu içeren tüplere ayrı kolonileri aktarın. Tüpleri gece boyunca 37 °C'de kuluçkaya yatırın ve 225 rpm'de titreyin.
- Plazmidleri plazmid miniprep kiti kullanarak gece kültüründen izole edin. Spektrofotometre kullanarak DNA'nın konsantrasyonu ve saflığını kontrol edin. 1.8 ile 2.0 arasında a260/A280 oranı kabul edilebilir.
- İstenilen klonlama üründoğru olup olmadığını belirlemek için Sanger sıralama için plazmidler gönderin. DNA'yı -20 °C'de saklayın.
2. Rekombinant virüs oluşturma
- 6 kuyulu bir plakada 1.0 (MOI = 1.0) enfeksiyonun çokluğuyla yeniden biraraya getirilecek virüsile uygun hücrelerden oluşan bir monotabakası enfekte edin. Enfekte hücreleri 37 °C'de ve %5 CO2'de 1 saat kuluçkaya yatırın. Daha sonra orta aspire ve taze DMEM ile değiştirin. Enfekte hücreleri 37 °C ve %5 CO2'dekuluçkaya yatırın.
NOT: K3L22yoksun bir vaccinia virüs gibi çoğaltma yetkili virüsler için, Avrupa tavşan böbrek hücre hattı RK13 (ATCC #CCL-37) veya BSC-40 gibi bir hücre hattı uygundur. Ancak, çoğaltma eksik virüsler için, bu makalede hem PKR antagonistleri E3L ve K3L yoksun virüs gibi, trans veya PKR knock-down veya knock-out hücreleri bu iki gen ifade tamamlayıcı bir hücre hattı gereklidir. - Üreticinin protokolünü izleyerek ticari olarak kullanılabilen bir transfeksiyon reaktifi kullanarak 1.10 adımda oluşturulan ve doğrulanmış vektörün 500 ng'si ile enfekte hücreleri dönüştürün. Hücreleri 37 °C'de ve %5 CO2'de 48 saat kuluçkaya yatırın.
NOT: Hem E3L hem de K3L'den yoksun bir vaccinia virüsü kullanıyorsanız, PKR aracılı selektif basınç yeniden birleştirilmiş virüslerin seçilmesini sağlayacak ve bu hücrelerde mCherry-E3L füzyon proteininin ekspresyonunu koruyacaktır. İstenirse, tüm plazmid yerine transfeksiyon için kullanılacak sadece kesici uç pcr yükseltmek de mümkün olmalıdır. - Enfeksiyondan 48 saat sonra, enfekte monolayer'ı hasat edin. Bazı durumlarda, hücreler pipetleme ile hasat edilebilir, ancak hala sıkıca yapıştırılır, bir hücre kazıyıcı ile hasat. Hücreleri üç kez dondurun ve %50 genlikte 15 s'ye sonicate edin. Kullanıma hazır olana kadar -80 °C'de saklayın.
- Seri olarak 10 kat seyreltin adım 2.3 10-10 -6 -6 1080 μL DMEM (10-1)için lysate 120 μL ekleyerek ve daha sonra dmem 1080 μL (10-2), ve bu dört kez daha tekrarlayarak bu seyreltme. Bu durumda RK13 hücrelerinin bir PKR yetkili hücre hattının bir birey, confluent iyi her seyreltme 1 mL ekleyin.
- Enfekte hücreleri 37 °C'de ve %5 CO2'de 1 saat kuluçkaya yatırın. Daha sonra ortamı aspire edin ve enfekte hücreleri 37 °C ve %5 CO2'detaze DMEM Kuluçkaya yatırın.
- Enfeksiyon sonrası 24 ila 48 saat, floresan mikroskopi ile rekombinant virüsleri tanımlayın. Rekombinant virüslerden gelen plaklar mCherry-E3L füzyon geninin entegrasyonu nedeniyle kırmızı floresanifade eder (Şekil 2). Başlangıçta PKR inhibitörlerinden yoksun bir virüs kullanılmışsa, tüm plaklar rekombinant virüs içerecektir.
- Plak RK13 hücrelerinde rekombinant virüsleri üç kez arındırın. Plak arınması son turdan sonra, tüm plaklar kırmızı floresan ifade etmelidir.
- VACV PKR inhibitörleri E3L ve K3L (RK13+E3L+K3L hücreleri28)ifade eden rk13 hücrelerinin bir confluent 6-iyi plaka enfekte adım 2.6 plak saflaştırılmış kırmızı floresan virüs ile. Her kuyuda yaklaşık 50-100 plak hedefleyin.
NOT: Bu hücreler trans VACV PKR antagonistleri sağlamak ve mCherry-E3L füzyon geni korumak için PKR aracılı seçici basınç hafifletmek, böylece rekombinant virüsün "scarless" nesil teşvik. - EVOS2 mikroskobu ve GFP filtre küpü (Uyarma: 470/22, Emisyon: 525/50) ve RFP filtre küpü (Ekscb: 531/40, Emisyon: 593/40) kullanarak çökmüş virüsleri tanımlayın.
NOT: MCherry-E3L füzyon geninin kaybolma sıklığı yaklaşık %2.5 'dir(Tablo 2). EGFP bir marker geni olarak dahil edilmezse, mCherry-E3L füzyon genini kaybetmiş mutant virüslerden gelen plaklar renksiz olacaktır. - Plak, RK13+E3L+K3L hücrelerinde üç kez sadece yeşil (VC-R4) veya renksiz plakları (E3L) arındırır. Hiçbir plak floresan kırmızı emin olun.
- MCherry-E3L kaybını ve PCR ve Sanger dizilimi ile beklenen mutasyonun varlığını doğrulayın.
NOT: İlgide pkr inhibitör aktivitesi yoksa RK13+E3L+K3L hücrelerinde veya eşdeğer PKR inhibitlü veya PKR eksik hücre hattında rekombinant virüsler yetiştirilmelidir(Şekil 3).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Şekil 1'de diyagramlanan yordamı, Hem PKR antagonistleri E3L hem de K3L'den yoksun bir VACV oluşturmak için, E3L'yi K3L (vP872) için silinmiş bir virüste EGFP ile değiştirerek kullandık. Şekil 2 PKR yetkili RK13 hücrelerinde kırmızı floresan plaklar mCherry-E3L viral ekspresyonu gösteren yanı sıra RK13 +E3L+K3L hücrelerinde e3L kaybını ve mCherry-E3L seçim belirteci çöküşünü doğrulayan EGFP'yi göstermektedir. Şekil 3 bu rekombinant virüs, VC-R4, her iki PKR antagonistleri yoksun PKR yetkili RK13 hücrelerinde çoğaltmak değil, ana virüs ise, vP872 E3L ifade, çoğaltma yetkili olduğunu doğrular. RK13 hücrelerinde çoğalamamasının sadece E3L kaybından kaynaklandığını doğrulamak için VC-R4'teki EGFP'yi E3L ile değiştirdik ve aynı seçim protokolünü kullanarak geri dönen bir virüs oluşturduk. Şekil 3 ayrıca bu geri dönen virüsün RK13 hücrelerinde vP872 kadar verimli bir şekilde çoğaldığını da doğrular. İlginçtir ki, mCherry-E3L seçim markerinin çöküşüile tutarlı renksiz plaklar, genellikle "scarless" rekombinantları seçmek için gerekli olan RK13+E3+K3 hücrelerinde seçimden önce tanımlanmıştır, muhtemelen mCherry-E3L rekombinasyon kaseti ile VC-R4'e eklenen E3L geni arasındaki genişletilmiş sırakimliği nedeniyle. Bu nedenle, rekombinasyon verimliliğini belirlemek ve erken çökme sorunu önlemek için poxvirus PKR antagonist K3L ifade virüsler üretmek için seçti çöküş oranını23. Şekil 4, RK13+E3L+K3L hücrelerinin enfeksiyonundan sonra renksiz plakların (ok uçları) görünümünü gösterir. Tablo 1 üç bağımsız deneylerin sonuçlarını gösterir, burada nesilvirions ortalama% 12.6 transfected plazmid ile rekombinasyon geçirmişti, daha önce bildirilen frekansları benzer29,30,31. Tablo 2, RK13+E3L+K3L hücrelerindeki toplam plaklara göre renksiz plakların sıklığını ayrıntılı olarak belirterek, mCherry-E3L seçim belirtecinin çökme ve kayıp oranını yaklaşık %1,8'lik bir frekansta gösterir.
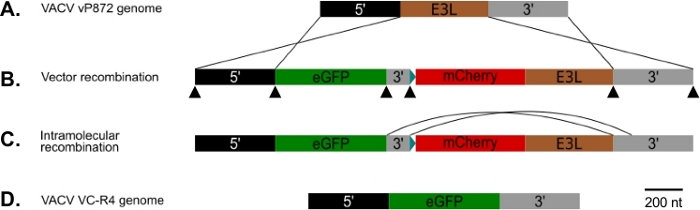
Şekil 1: P837-GOI-mCherry-E3L diyagramı ve ana bilgisayar aralığı ve görsel rekombinasyon stratejisi. (A) 5' kol (siyah) ve 3' kol (gri) VACV'deki E3L locus (kahverengi) kanadı. (B) P837-GOI-mCherry-E3L'de, bu kollar ilgi genini içeren bir kaseti (GOI), bu durumda EGFP, (yeşil) sentetik erken/geç poxvirus promotörü25 mavinin kontrolü altında bulunan bir mCherry-E3L (kırmızı) füzyon geninden ayrılır) kısa bir 3' kol (gri) ile böler. Bu dış kollar VACV ve p837-GOI-mCherry-E3L arasında homolog rekombinasyon sağlar. Siyah ok uçları Gibson klonlama tarafından bu plazmid oluşturmak için kullanılan örtüşen astar siteleri gösterir. (C) PKR selektif basınç çıkarıldığında, kısa ve uzun 3' kollar arasında intramoleküler rekombinasyon geçiren virüsler seçilebilir. (D) E3L lokussadece ilgi geni içeren bir virüs (VC-R4) ile sonuçlanan. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 2: RK13 hücrelerinde hem mCherry (sol) hem de EGFP (sağda) ifade eden p837-GOI-mCherry-E3L ile rekombinasyondan 24 saat sonra rekombinant bir virüs plağı (üstte) floresan mikrograflar. (Alt) RK13++ hücrelerinde PKR aracılı selektif basınç çıkarıldıktan 48 saat sonra rekombinant virüs plak mikrografı, EGFP 'yi (sağda) ifade eder, ancak mCherry'yi (solda) ifade etmez. Ölçek çubuğu tüm paneller için 650 μm'yi gösterir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Şekil 3: VC-R4 PKR yetkili hücrelerde çoğalamaz. Belirtilen hücre hatları nai = 0.1'de vP872 (mavi), VC-R4 (yeşil) veya VC-R4+E3L (macenta) ile enfekte oldu. 48 saat enfeksiyon sonrası enfekte hücreler rk13+E3L+K3L hücrelerinde seri seyreltme ile hasat edildi ve titretedildi. Titreler PFU/mL olarak rapor edilir, hata çubukları üç çoğaltma deneyinin standart sapmasını temsil edilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 4: RK13+E3L+K3L hücrelerinde mCherry-E3L ekspresyonunun kaybı. VC-R4+K3L-mCherry-E3L enfekte RK13+E3L+K3L hücrelerinin floresan ve faz kontrast mikrograflarının yer kaplaması. Vc-R4+K3L ile oluşan seçilim kasetinin çökmesi nedeniyle üç plak artık mCherry'yi (daireleri) ifade etmez. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
| Deney 1 | Deney 2 | Deney 3 | |
| Kırmızı plaklar (RK13) | 30 | 11 | 18 |
| Toplam plaklar (RK13+E3L+K3L) | 225 | 64 | 249 |
| Rekombinasyon Oranı | 13.30% | 17.20% | 7.20% |
Tablo 1: VACV'nin p837-K3L-mCherry-E3L plazmidile rekombinasyon frekansı.
| Deney 1 | Deney 2 | Deney 3 | |
| Toplam plaklar (RK13+E3L+K3L) | 115 | 44 | 210 |
| Renksiz plaklar (RK13+E3L+K3L) | 3 | 1 | 1 |
| Rekombinasyon Oranı | 2.60% | 2.30% | 0.50% |
Tablo 2: RK13+E3+K3 hücrelerinde VC-R4+K3L-mCherry-E3L'den mCherry-E3L kaybı sıklığı.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Burada son rekombinant virüs yabancı DNA tutmadan rekombinant vaccinia virüsleri oluşturmak için geçici bir marker seçim stratejisi 32 bir varyasyonu sıyoruz. Stratejimiz, antibiyotikler gibi diğer seçici basınç biçimleri yerine ev sahibi antiviral protein PKR aracılığındaki seçici basıncı kullanır. Konak antiviral genlerin kullanımı hücrelerde kimyasal kaynaklı fenotipik değişiklikler olasılığını ortadan kaldırır, ya da seçim ilaçları nedeniyle mutasyon riskinin artması. Ayrıca, ilaç seçiminden farklı olarak, yaklaşımımız için bir gecikme fazı yoktur, çünkü PKR tüm hücrelerde kurucu olarak ifade edilir. mCherry ekspresyonuna dayalı ikincil görsel seçilim, sadece transgeni ifade eden plakların ilk aşamada seçilmesini sağlayarak bu yöntemin özgüllüğünü artırır ve mCherry-E3L genini kaybetmiş olgun rekombinant virüsleri seçerken negatif selektif belirteç olarak eşit derecede etkilidir.
Bu rekombinasyon stratejisi için en kritik adımlar uygun rekombinasyon vektörü üretimi ve seçilen virüsün klonal olmasını sağlamak için uygun plak arınmasıdır. Bu yazıda rekombinasyon vektörü oluşturmak için "Gibson montaj" öneririz. Bu strateji son derece verimlidir ve rekombinasyon vektöründen oluşan tüm parçaların tek bir günde birleştirilmesine olanak sağlar. Ancak, kısa 3' kol ve uzun 3' kol aynı dizileri paylaştığından, bu parçaların klonlama reaksiyonu sırasında biraraya geleme potansiyeli vardır ve bazı vektörler mCherry-E3L kasetini içermeyebilir. Deneyimlerimize göre bu nadirdir ancak klonlama dan sonra vektörün yapısını onaylamak gereklidir. Ayrıca geleneksel endokleaz ve ligaz yöntemleri ni kullanarak bu strateji için rekombinasyon vektörleri oluşturduk. Bu strateji, yukarıda açıklanan sorunu önler, ancak daha fazla emek yoğun olabilir. Plak arınması genellikle basittir ve öncelikle ilk rekombinasyon için uygun izin hücreleri kullanarak bağlıdır, PKR-sadece rekombinant virüsler çoğalabilir sağlamak için ilk plak saflaştırma için pkr yetkili hücreleri, ve daha sonra tekrar intramoleküler rekombinasyon ve seçilebilir belirteç kaybını kolaylaştırmak için izin hücreleri. Bu nedenle hücre hatlarına yakın ilgi, bu stratejinin başarılı ve verimli bir şekilde uygulanması için çok önemlidir.
Bu çalışmada, hem PKR antagonistleri E3L hem de K3L için silinmiş bir VACV rekombinant oluşturmak ve EGFP'yi E3L organizatörünün kontrolü altında ifade etmek için bu yöntemin kullanımını gösteriyoruz. İleriye dönük olarak, bu virüs gelecekteki rekombinant virüsler için etkili bir arka plan olarak hizmet verecek, PKR yetkili hücrelerde çoğaltma aciz olduğu gibi. Bu nedenle, aynı zamanda aslında non-rekombinant virüsün replikasyonunu önlerken, mCherry-E3L rekombinasyon kasetini nesilvirions içine sürmek için güçlü PKR aracılı seçici basınç olacaktır. Ayrıca, rekombinasyon kaset alımı ile EGFP kaybı, seçilmiş plakların rekombinant olmayan bir virüsle birlikte enfekte olmadığından emin olmak için yararlı bir ikincil seçim belirtecidir. VACV için daha önce bildirilen oranlar ile tutarlı rekombinasyon oranları gözlemledik, ancak görsel floresan belirteçler uygun rekombinant virüslerin seçilme olasılığını artırarak rekombinant virüsler imal etme verimliliğini artırır. PKR-yetkili hücrelerde iki tur seçimden sonra renksiz plaklar gözlemimiz, muhtemelen E3L ve mCherry-E3L marker geni arasındaki aynı dizinin uzunluğunun artması nedeniyle, 3' kısa kolun uzunluğunu artırarak veya azaltarak mCherry-E3L kaybı oranının "ayarlanabileceğini" düşündürmektedir. Bu tekniğin birincil sınırlamarekler için seçici basınç olarak PKR kullanımıdır. Bu rekombinasyon stratejisinin en verimli kullanımı PKR antagonistleri eksik bir arka planda bu virüsleri üreten. Ancak, kolorimetrik seçim marker bu rekombinasyon stratejisi pkr aracılı seçim olmadan bile kullanılmasını sağlar, sadece plak mCherry ifade plaket leri arındırıcı tarafından. PKR aracılı seçici basınç eksikliği ilk tarama adımının verimliliğini azaltacak olsa da, mCherry ifade plakları yüzdesi hala renk tabanlı seçim uygulanabilir yeterince yüksektir. Böylece, bu yöntem poxvirus genomuna hemen hemen herhangi bir gen eklemek için kullanılabilir.
EGFP'nin eklenmesiyle gösterildiği gibi, bu yaklaşımla, transgen PKR antagonisti değilse, PKR null hücreleri veya iltifat hücre hatlarının alt akım deneyleri için kullanılması koşuluyla, herhangi bir gen hızla yerli organizatörün kontrolü altında E3L çekirgesine yerleştirilmesi mümkündür. Bu strateji, burada rapor VC-R4 virüsü ile birlikte, hızlı ve güvenilir bir şekilde ev sahibi aracılı seçici basınç ve sürecin başlarında rekombinantgörsel tanımlama kullanarak rekombinant vaccinia virüsleri oluşturmak için yeni ve güçlü bir yöntem ekler.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarlar hiçbir rakip mali çıkarları beyan.
Acknowledgments
Bu proje Ulusal Sağlık Enstitüleri (AI114851) tarafından SR'ye finanse edilmiştir.
Materials
| Name | Company | Catalog Number | Comments |
| 2X-Q5 Master Mix | NEB | M0492L | High-fidelity polymerase used in PCR |
| Ampicillin | ThermoFisher Scientific | 11593027 | Bacterial selective agent |
| Disposable Cell Scrapers | ThermoFisher Scientific | 08-100-242 | Cell scraper to harvest infected cells |
| EVOS FL Auto 2 Cell imaging system | ThermoFisher Scientific | AMAFD2000 | Fluorescent microscope |
| EVOS Light Cube, GFP | ThermoFisher | AMEP4651 | GFP Cube |
| EVOS Light Cube, RFP | ThermoFisher | AMEP4652 | RFP Cube |
| GenJet | SignaGen Laboratories | SL100489 | Transfection reagent |
| Luria Bertani (LB) Broth | Gibco | 10855021 | Bacterial growth medium |
| Monarch DNA gel extraction kit | NEB | T1020L | Gel purification kit used to purify amplicons and linearized vectors |
| Monarch Plasmid Miniprep kit | NEB | T1010L | Miniprep kit ussed to purify plasmids |
| NanoDrop One | ThermoFisher Scientific | ND-ONE-W | Spectrophotometer used to measure RNA and DNA concentration |
| NEBuilder Master Mix | NEB | E2621L | Isothermal enzymatic assembly kit used to generate the recombination vector |
| Q500 Sonicator | Qsonica | Q500-110 | Sonicator for virus lysates |
| RK13 cells | ATCC | CCL-37 | Rabbit kidney cells |
| VWR Multiwell Cell Culture plates | VWR | 10062-892 | Cell culture plates |
References
- Brochier, B., et al. Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Nature. 354 (6354), 520-522 (1991).
- Pastoret, P. P., Brochier, B. The development and use of a vaccinia-rabies recombinant oral vaccine for the control of wildlife rabies; a link between Jenner and Pasteur. Epidemiology and Infection. 116 (3), 235-240 (1996).
- Chan, W. M., McFadden, G.
- Nguyen, D. H., et al. Vaccinia virus-mediated expression of human erythropoietin in tumors enhances virotherapy and alleviates cancer-related anemia in mice. Molecular Therapy. 21 (11), 2054-2062 (2013).
- Frentzen, A., et al. Anti-VEGF single-chain antibody GLAF-1 encoded by oncolytic vaccinia virus significantly enhances antitumor therapy. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12915-12920 (2009).
- Pastoret, P. P., Vanderplasschen, A.
- COLLIER, L. H. The development of a stable smallpox vaccine. The Journal of Hygiene. 53 (1), 76-101 (1955).
- Weir, J. P., Bajszár, G., Moss, B. Mapping of the vaccinia virus thymidine kinase gene by marker rescue and by cell-free translation of selected mRNA. Proceedings of the National Academy of Sciences of the United States of America. 79 (4), 1210-1214 (1982).
- Mackett, M., Smith, G. L., Moss, B. Vaccinia virus: a selectable eukaryotic cloning and expression vector. Proceedings of the National Academy of Sciences of the United States of America. 79 (23), 7415-7419 (1982).
- Nakano, E., Panicali, D., Paoletti, E. Molecular genetics of vaccinia virus: demonstration of marker rescue. Proceedings of the National Academy of Sciences of the United States of America. 79 (5), 1593-1596 (1982).
- Falkner, F. G., Moss, B. Transient dominant selection of recombinant vaccinia viruses. Journal of Virology. 64 (6), 3108-3111 (1990).
- Staib, C., Drexler, I., Ohlmann, M., Wintersperger, S., Erfle, V., Sutter, G. Transient Host Range Selection for Genetic Engineering of Modified Vaccinia Virus Ankara. BioTechniques. 28 (6), 1137-1148 (2000).
- Staib, C., Drexler, I., Sutter, G. Construction and Isolation of Recombinant MVA. Vaccinia Virus and Poxvirology. , 77-99 (2004).
- Di Lullo, G., et al. Marker gene swapping facilitates recombinant Modified Vaccinia Virus Ankara production by host-range selection. Journal of Virological Methods. 156 (1-2), 37-43 (2009).
- Pfaller, C. K., Li, Z., George, C. X., Samuel, C. E. Protein kinase PKR and RNA adenosine deaminase ADAR1: New roles for old players as modulators of the interferon response. Current Opinion in Immunology. 23 (5), 573-582 (2011).
- Bevilacqua, P. C., George, C. X., Samuel, C. E., Cech, T. R. Binding of the protein kinase PKR to RNAs with secondary structure defects: Role of the tandem A - G mismatch and noncontigous helixes. Biochemistry. 37 (18), 6303-6316 (1998).
- Krishnamoorthy, T., Pavitt, G. D., Zhang, F., Dever, T. E., Hinnebusch, A. G. Tight Binding of the Phosphorylated Subunit of Initiation Factor 2 (eIF2) to the Regulatory Subunits of Guanine Nucleotide Exchange Factor eIF2B Is Required for Inhibition of Translation Initiation. Molecular and Cellular Biology. 21 (15), 5018-5030 (2001).
- Rothenburg, S., Georgiadis, M. M., Wek, R. C. Evolution of eIF2α kinases: Adapting translational control to diverse stresses. Evolution of the Protein Synthesis Machinery and Its Regulation. , 235-260 (2016).
- Bratke, K. A., McLysaght, A., Rothenburg, S. A survey of host range genes in poxvirus genomes. Infection, Genetics and Evolution. 14, 406-425 (2013).
- Chang, H. W., Watson, J. C., Jacobs, B. L. The E3L gene of vaccinia virus encodes an inhibitor of the interferon-induced, double-stranded RNA-dependent protein kinase. Proceedings of the National Academy of Sciences. 89 (11), 4825-4829 (1992).
- Romano, P. R., et al. Inhibition of double-stranded RNA-dependent protein kinase PKR by vaccinia virus E3: role of complex formation and the E3 N-terminal domain. Molecular and Cellular Biology. 18 (12), 7304-7316 (1998).
- Beattie, E., Tartaglia, J., Paoletti, E. Vaccinia virus-encoded eIF-2 alpha homolog abrogates the antiviral effect of interferon. Virology. 183 (1), 419-422 (1991).
- Park, C., Peng, C., Brennan, G., Rothenburg, S. Species-specific inhibition of antiviral protein kinase R by capripoxviruses and vaccinia virus. Annals of the New York Academy of Sciences. 1438 (1), 18-29 (2019).
- Rothenburg, S., Brennan, G. Species-Specific Host-Virus Interactions: Implications for Viral Host Range and Virulence. Trends in Microbiology. , (2019).
- Chakrabarti, S., Sisler, J. R., Moss, B. Compact, synthetic, vaccinia virus early/late promoter for protein expression. BioTechniques. 23 (6), 1094-1097 (1997).
- Chung, C. T., Niemela, S. L., Miller, R. H. One-step preparation of competent Escherichia coli: Transformation and storage of bacterial cells in the same solution (recombinant DNA). Biochemistry. 86, 2172-2175 (1989).
- Chung, C. T., Miller, R. H. Preparation and storage of competent Escherichia coli cells. Methods in Enzymology. 218, 621-627 (1993).
- Rahman, M. M., Liu, J., Chan, W. M., Rothenburg, S., McFadden, G. Myxoma Virus Protein M029 Is a Dual Function Immunomodulator that Inhibits PKR and Also Conscripts RHA/DHX9 to Promote Expanded Host Tropism and Viral Replication. PLOS Pathogens. 9 (7), 1003465 (2013).
- Evans, D. H., Stuart, D., McFadden, G. High levels of genetic recombination among cotransfected plasmid DNAs in poxvirus-infected mammalian cells. Journal of Virology. 62 (2), 367-375 (1988).
- Ball, L. A. High-frequency homologous recombination in vaccinia virus DNA. Journal of Virology. 61 (6), 1788-1795 (1987).
- Spyropoulos, D. D., Roberts, B. E., Panicali, D. L., Cohen, L. K. Delineation of the viral products of recombination in vaccinia virus-infected cells. Journal of Virology. 62 (3), 1046-1054 (1988).
- Liu, L., et al. Transient dominant host-range selection using Chinese hamster ovary cells to generate marker-free recombinant viral vectors from vaccinia virus. BioTechniques. 62 (4), 183-187 (2017).
