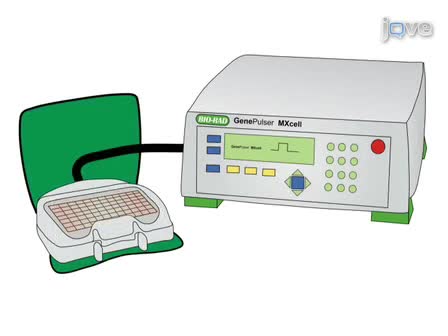
This procedure describes a method for the isolation and culture of the murine organ of Corti with or without the spiral limbus and spiral ganglion neurons. We also demonstrate a method for the expression of an exogenous reporter gene in the organ of Corti explant by electroporation.