PJ-34 is a stable water soluble phenanthridine1 (Figure 1). Our previous results revealed cell death and de-clustered extra-centrosomes in several types of fixed multi-centrosomal cancer cells that were treated with PJ-34. In contrast, normal proliferating cells were not impaired2,3. Centrosomes were identified by double labeling with antibodies directed against centrine1 and γ-tubulin in the fixed extra-centrosomal cells2.
Here, the cytotoxic activity of PJ-34 was documented in these live extra-centrosomal cells at real time by using live confocal microscopy. Live human breast cancer MDA-MB-231 cells, which have a high occurrence (>50%) of extra-centrosomes4,5, were scanned for at least 16 hr by confocal imaging focused on transfected cells with γ-tubulin-GFP (fluorescent labeling of γ-tubulin foci2) and with histone H2b-RED (fluorescent labeling of chromosomes). Six to ten live transfected cells were scanned in parallel in each experiment. Double immunolabeling of the γ-tubulin foci in the transfected cells with centrin1 was technically impossible.
Dispersed γ-tubulin foci and aberrant chromosomes arrangement were rarely detected in randomly selected untreated MDA-MB-231 cells in mitosis. Bifocal clustering of γ-tubulin foci, representing extra-centrosomes bifocal clustering, was documented in the majority of live untreated MDA-MB-231 cells (Figure 2), In contrast, un-clustered centrosomes and aberrant arrangement of chromosomes were detected in live transfected MDA-MB-231 cells incubated with PJ-34 (20 μM), and mitosis in these cells ended by cell death (Figure 3). These cells were incubated with PJ-34 for 18 – 24 hr before scanning and for additional 16 hr during scanning (Figure 3). The real-time documentation of cell death during mitosis strongly supports a previously defined positive correlation between the number of human malignant cells with multi-polar spindles in mitosis and the percentage of cell death in cells incubated with PJ-34 (20 μM)2.
PJ-34 acts as a potent PARP1 inhibitor1. We therefore examined the possibility of PARP1 inhibition causing cell death associated with mitotic failure. Unlike live imaging, imaging of fixed MDA-MB-231 cells enabled examination of a large population of cells in the cell cultures, thereby enabling statistical analysis of the effects of PARP1 inhibitors in a variety of human cancer cell lines. The activity of PJ-34 was compared to the activity of other potent, non-phenanthrene PARP1 inhibitors in normal or PARP1 deficient cells (i.e. normal and PARP1(-/-) mouse embryonic fibroblasts (MEF)) (Figure 4). PARP1 deficient MEF harbor multi-centrosomes in mitosis, but they are not tumor cells11. These cells were prepared by Dr. Francoise Dantzer, Strasbourg, France.
Fixed normal and PARP1(-/-) MEF were immunolabeled for α- and γ-tubulin that labeled their spindles and centrosomes, respectively, as reported before2. Some of the examined cell cultures were treated with PJ-34 or other potent, non-phenanthrene PARP1 inhibitors, including ABT-888 and AG01469, which inhibit the enzymatic activity of PARP1, and BSI-201, a compound that apparently attenuates PARP1 binding to nicked DNA12-14. None of the tested PARP1 inhibitors impaired normal MEF at concentrations inhibiting PARP1 activity (Figure 4). In contrast, PJ-34 dose-dependently caused un-clustering of γ-tubulin foci, distortion of spindles and cell death in PARP1(-/-) MEF (Figures 4A and B). This was not observed in normal MEF treated with PJ-34 (Figure 4B) or in PARP1(-/-) MEF treated with non-phenenthrene PARP1 inhibitors ABT-888 or AG014699 (Figure 4C). It should be noted that PJ-34 at concentrations exceeding 20 μM did impair normal MEF, although normal MEF were more resistant to PJ-34 activity than PARP1(-/-) MEF.
The fact that PJ-34 eradicated PARP1(-/-) MEF despite their PARP1 deficiency, and the correlation between the formation of multi-focal spindles and cell eradication in PARP1(-/-) MEF incubated with PJ-34 at concentrations higher than those required for PARP1 inhibition, were not consistent with a causal linkage between extra-centrosomes de-clustering in PARP1(-/-) MEF and PARP1 inhibition (Figure 4A). The cytotoxic activity of PJ-34 in PARP1(-/-) MEF could be better explained by its activity as an extra-centrosomes de-clustering agent in multi-centrosomal cells2 (Figure 3). Thus, the combination of live confocal imaging and immunocytochemistry methods was useful for identifying cytotoxic mechanisms impairing mitosis.

Figure 1. The phenanthridine PJ-34: N-(6-oxo-5,6-dihydro-phenanthridin-2-yl)-N,N-dimethyl-acetamide.

Figure 2. Bi-focal clustering of extra-centrosomes in a randomly selected live MDA-MB-231 cell in mitosis. A. Upper panel: Labeled centrosomes in a randomly selected live MDA-MB-231 cell transfected with γ-tubulin-GFP. Lower panel: Chromosome re-arrangements during mitosis in a randomly selected MDA-MB-231 cell transfected with histone H2b-RED. B. Bi-focal mitosis with clustered extra-centrosomes identified in a randomly selected cultured MDA-MB-231 cell. Cells were transfected by both γ-tubulin-GFP (labeling γ-tubulin foci; green) and histone H2b-RED (labeling chromosomes; red). 48 hr after transfection, cells were exposed to a live confocal imaging for 16 hr. Six cells were scanned in parallel in each experiment. Four different experiments were performed. See also Supplementary Information. Click here to view larger figure.

Figure 3. Extra-centrosomes de-clustering preceded cell death in live MDA-MB-231 cells treated with PJ-34. A randomly selected live MDA-MB-231 cell in mitosis with scattered centrosomes (1st frame on left) ended by cell death (2nd and 3rd frames). This cell was randomly selected in a cell culture incubated for 24 hr with PJ-34 (20 μM) applied 24 hr after transfection with vectors expressing γ-tubulin-GFP (labeling γ-tubulin foci including centrosomes; green) and histone H2b-RED (labeling chromosomes; red). The cell was scanned for 16 hr by live confocal imaging. Six cells were scanned in parallel in each experiment. Three different experiments were performed. See also Supplementary Information. Click here to view larger figure.

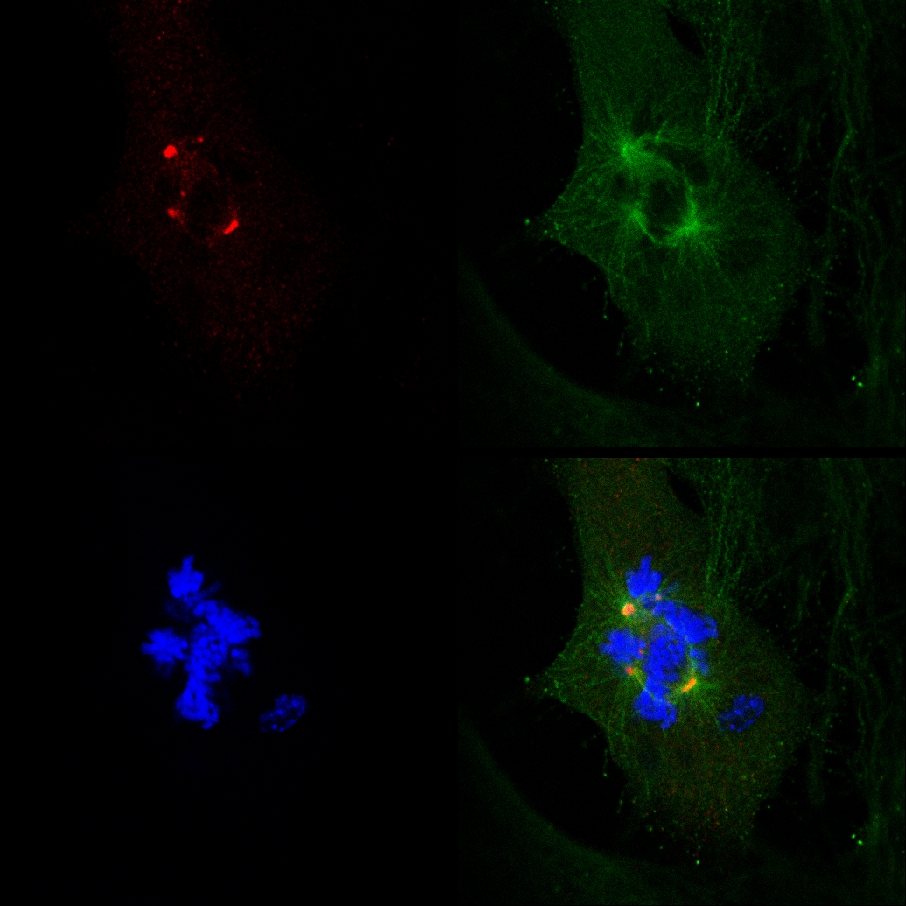
Figure 4. A cytotoxic activity of PJ-34 in PARP1(-/-) mouse embryonic fibroblasts. A. (Left) The percentage of multi-focal spindles calculated in normal (black line) and Parp1-/- (grey line) MEF, incubated for 48 hr with PJ-34 at the indicated concentrations. The percentage of multi-polar spindles was calculated out of 20 total spindles detected in 3 different experiments. (Right) Reduced cell survival detected in cell cultures incubated for 72 hr with PJ-34 (20 μM) relative to the survival of control untreated cells (normal (black line) and Parp1-/- (grey line) MEF). Cell survival was assayed by the cells’ ATP production (protocol 5). The mean values of 4 measurements for each cell line in 3 different experiments are presented. B. Spindles in randomly selected fixed normal and Parp1-/- MEF in mitosis, untreated (control) or incubated with PJ-34 for 48 hr at the indicated concentrations. PJ-34 caused multipolar spindles. Cells were fixed, permeabilized and immunolabeled for α- and γ-tubulin (green labeling of spindles and red labeling of centrosomes, respectively). Chromosomes were labeled with DAPI reagent (blue). Representative results of 3 different experiments. C. Potent non-phenanthrene PARP1 inhibitors did not affect centrosomes clustering in PARP1(-/-) MEF. Spindles of randomly selected normal and Parp1-/- MEF are presented; untreated MEF (control) or MEF treated for 48 hr with non-phenanthrene PARP inhibitors, AG01469 (20 μM) or ABT888 (20 μM). Chromosomes are labeled with DAPI reagent (blue). Similar results were obtained in 3 different experiments. Click here to view larger figure.
Supplementary Information
Click here to view larger figure.
Click here to view larger figure.
Figure 2B. Supplemental Figure. A randomly selected MDA-MB-231 cell in anaphase with bi-focal clustered γ-tubulin labeled extra-centrosomes was scanned for 16 hr by live confocal imaging, 48 hr after being transfected with vectors expressing γ-tubulin-GFP and H2b-RED (labeling γ-tubulin foci and centrosomes in the fixed cells (green) and H2b histone labeling of the chromosomes (red), respectively).
Figure 3. Click here to view movie. Supplemental Figure.
A live confocal imaging documentation of cell death in a randomly selected MDA-MB-231 cell in mitosis with un-clustered γ-tubulin labeled extra-centrosomes. MDA-MB-231 cells were incubated with PJ-34 for 24 hr before scanning and during the 16 hr of live confocal imaging. PJ-34 was applied 24 hr after transfection with vectors expressing γ-tubulin-GFP (green, γ-tubulin foci and centrosomes) and H2b-RED (red, chromosomes).
