A figure outlining the procedural steps in the isolation of mitochondria using tissue dissociation and differential filtration is shown in Figure 1. Total procedural time is less than 30 min.
Tissue samples were obtained using a 6 mm biopsy punch. Tissue weight was 0.18 ± 0.04 g (wet weight). The number of mitochondria isolated as determined by particle size counting was 2.4 x 1010 ± 0.1 x 1010 mitochondria for skeletal muscle and 2.75 x 1010 ± 0.1 x 1010 mitochondria for liver preparations (Figure 2A). To allow for comparison mitochondrial number was also determined by hemocytometer. Mitochondrial number was underestimated as determined by hemocytometer as 0.11 x 1010 ± 0.04 x 1010 mitochondria for skeletal muscle and 0.34 x 1010 ± 0.09 x 1010 mitochondria for liver preparations (Figure 2A). Mitochondrial diameter as determined by size based particle counter is shown in Figure 2B. The representative tracing shows the isolated mitochondria are localized under one peak with mean diameter of 0.38 ± 0.17 µm in agreement with previous reports7.
Mitochondrial protein/g (wet weight) starting tissue as determined by Bicinchoninic Acid (BCA) assay was 4.8 ± 2.9 mg/g (wet weight) and 7.3 ± 3.5 mg/g (wet weight) for skeletal muscle and liver samples respectively (Figure 2C).
Mitochondrial purity was determined by transmission electron microscopy and is shown in Figure 2D. Mitochondria are shown to be electron dense with less than 0.01% being fractured or damaged. Contamination by non-mitochondrial particles is less than 0.001%.
Mitochondrial viability was determined by MitoTracker Red as previously described17,18. Our results show that the isolated mitochondria maintain membrane potential (Figures 3A – C).
ATP was determined using a luminescent assay kit. A plate map for the ATP assay is shown in Figure 4. ATP standards were plated in duplicate. Mitochondrial samples and negative controls were plated in triplicate. ATP content was 10.67± 4.38 nmol/mg mitochondrial protein and 14.83 ± 4.36 nmol/mg mitochondrial protein for skeletal muscle and liver samples respectively (Figure 3D).
Mitochondrial respiration was assessed using a Clark type electrode as previously described17,18. Mitochondrial oxygen consumption rate was 178 ± 17 nM O2/min/mg mitochondrial protein for skeletal muscle and 176 ± 23 nM O2/min/mg mitochondrial protein for liver preparations. Respiratory control index (RCI) values were 2.45 ± 0.34 and 2.67 ± 0.17 for skeletal muscle and liver sample preparations respectively (Figure 3E). These results are similar to those reported in our previous studies using manual homogenization and differential centrifugation to isolate mitochondria17-18.
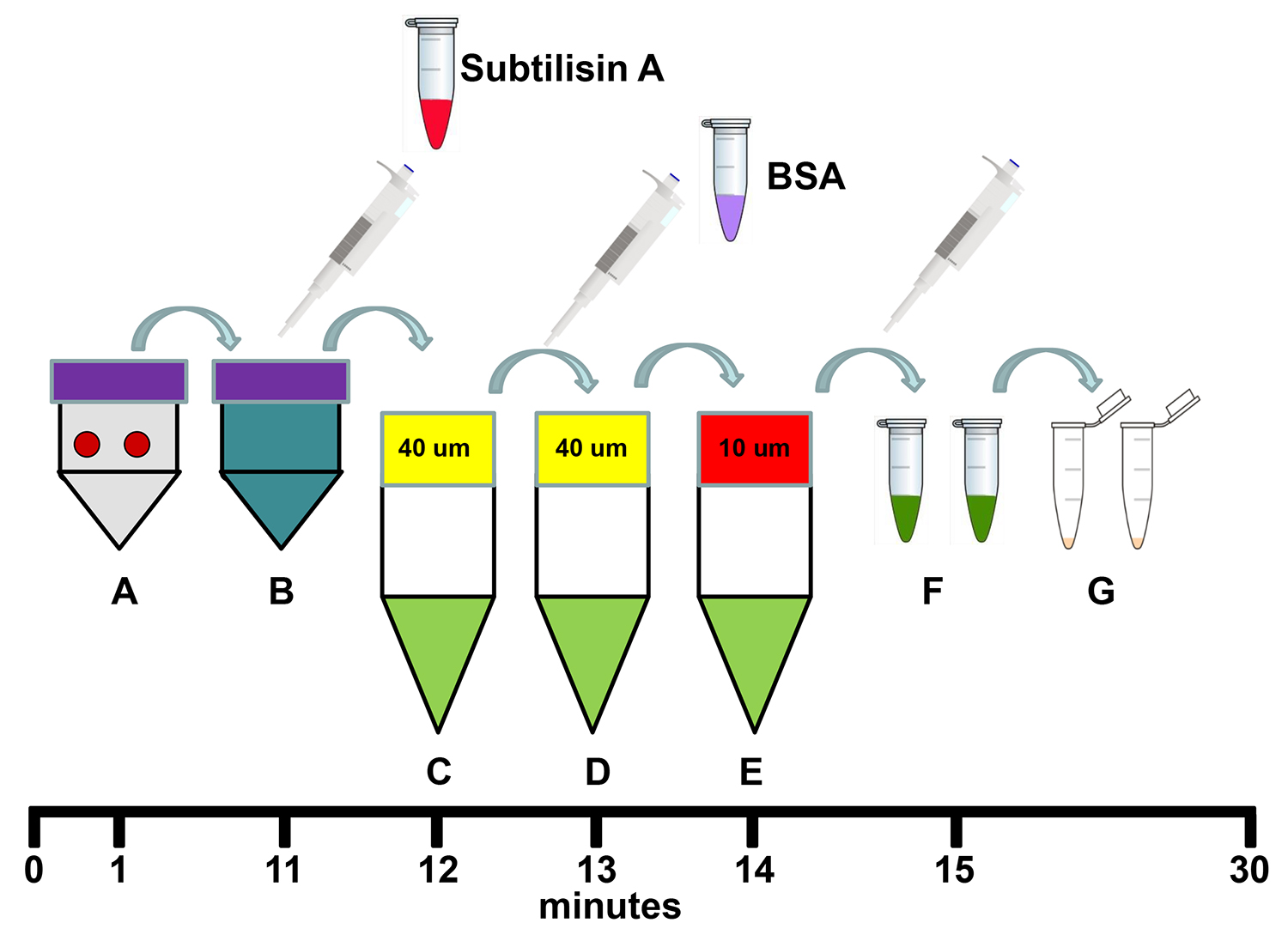
Figure 1. Schema for the isolation of mitochondria using tissue dissociation and differential filtration. (A) Transfer two 6 mm biopsy sample punches to 5 ml of Homogenizing Buffer in a dissociation C tube and homogenize the samples using the tissue dissociator's 1 min homogenization program. (B) Add 250 µl Subtilisin A stock solution to the homogenate in the dissociation C tube and incubate on ice for 10 min. (C) Filter the homogenate through a pre-wetted 40 µm mesh filter in a 50 ml conical centrifuge tube on ice and then add 250 µl of BSA stock solution to the filtrate. (D) Re-filter the filtrate through a new pre-wetted 40 µm mesh filter in a 50 ml conical centrifuge on ice. (E) Re-filter the filtrate through a new pre-wetted 10 µm mesh filter in a 50 ml conical centrifuge tube on ice. (F) Transfer the filtrate to 1.5 ml microfuge tubes and centrifuge at 9,000 x g for 10 min at 4 °C. (G) Remove the supernatant and re-suspend and combine the mitochondrial pellets in 1 ml Respiration Buffer. Total procedure time is less than 30 min. Please click here to view a larger version of this figure.
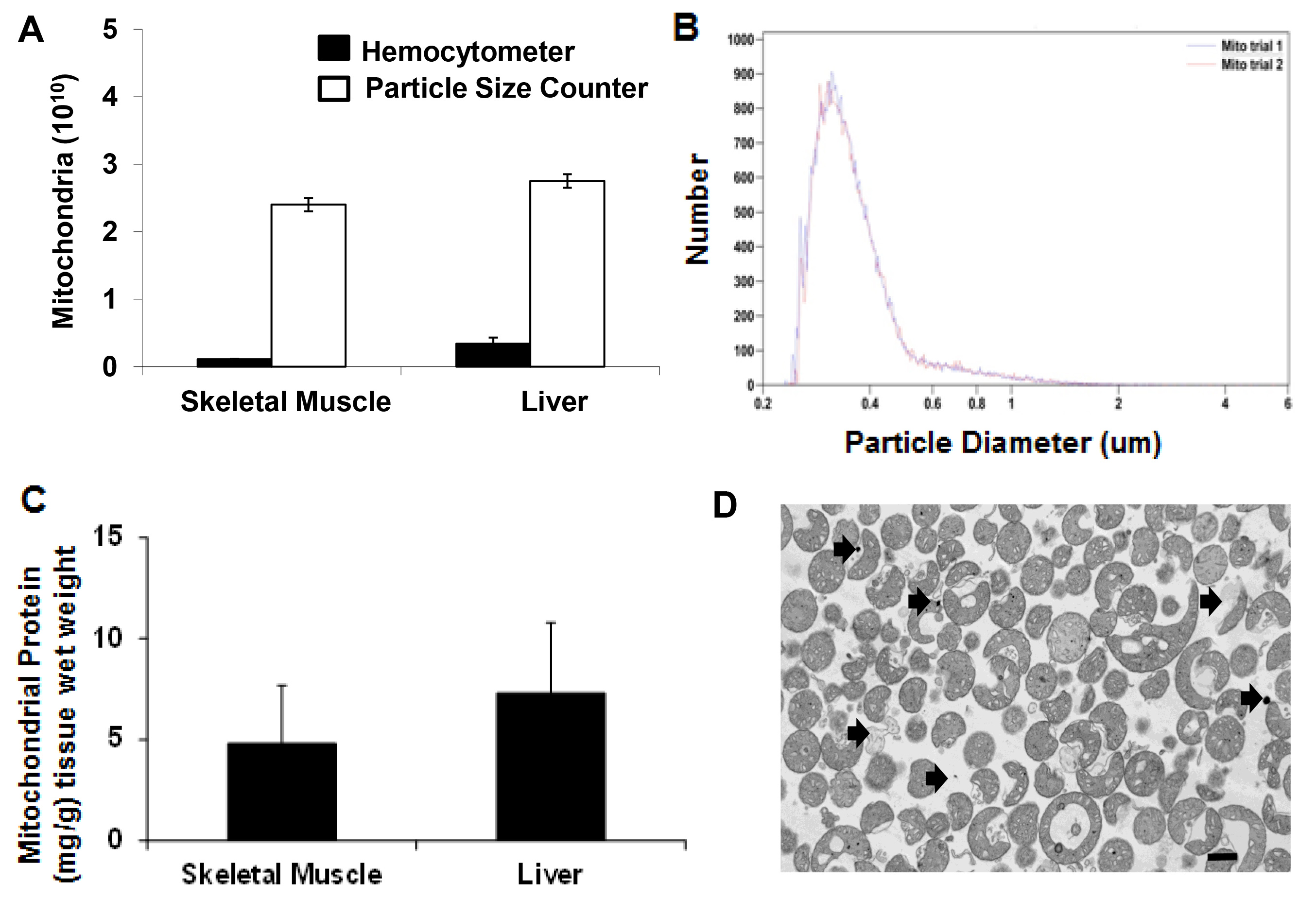
Figure 2. Mitochondrial yield and purity. (A) Hemocytometer and particle size counter mitochondria number isolated from 0.18 ± 0.04 g tissue (wet weight) for skeletal muscle and liver. (B) Mitochondrial size (mg/g) distribution as detected by particle size counter. (C) Mitochondrial protein mg/g tissue wet weight for skeletal muscle and liver. (D) Transmission electron microscopy image of isolated mitochondria. Scale bar is 100 nm. Arrows indicate possible contamination by non mitochondrial particles and damaged mitochondria. Please click here to view a larger version of this figure.
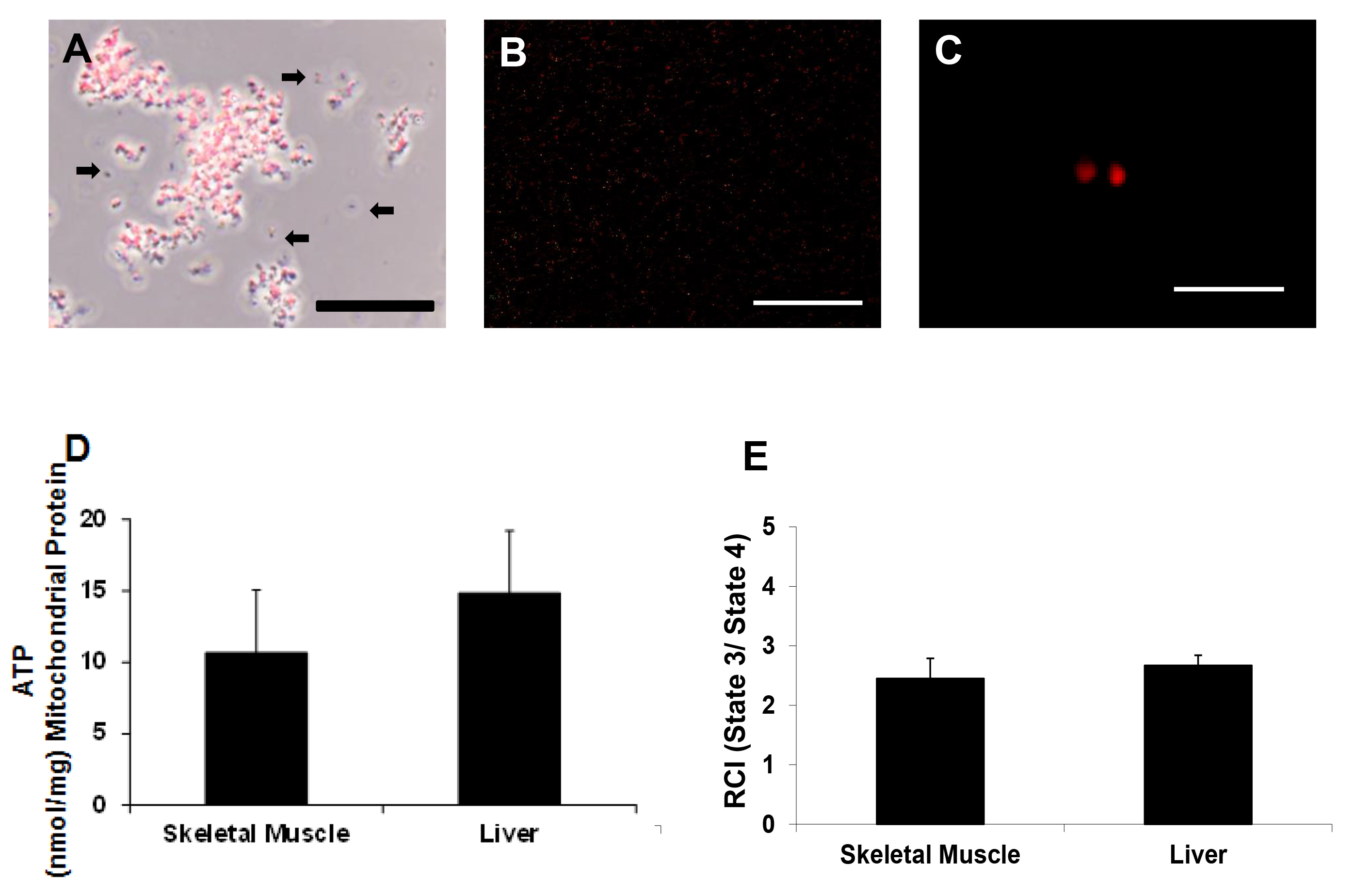
Figure 3. Mitochondrial viability. Representative photomicrographs of isolated mitochondria (A) under phase contrast illumination and (B and C) under fluorescence, with mitochondria labeled with MitoTracker Red CMXRos. Scale bars are 25 μm (A, B) and 5 μm (C). These images indicate that mitochondria maintained membrane potential. Arrows indicate mitochondria lacking membrane potential or debris (D) ATP content nmol/mg mitochondrial protein as determined by ATP assay and (E) RCI (state 3/state 4) as determined by Clark electrode. Please click here to view a larger version of this figure.

Figure 4. Plate map for ATP assay. This plate map illustrates how to set up standards (A1 – A12), mitochondria samples (B1 – C6), and negative controls (C7 – C9) for the ATP assay. During the assay, 100 µl of Respiration Buffer, 50 µl of mammalian cell lysis solution and 50 µl of reconstituted substrate solution are added to all wells (A1 – C9).
