NOTE: This protocol can be used for multiple applications such as quantification of phagocytosis and actin polymerization as used in our previously published work12. The protocol lends itself to a variety of modification, and Table 1 lists cell types and particles successfully utilized in past studies. Standard use of this protocol for quantification of phagocytosis or actin polymerization is illustrated in Diagram 1.

Diagram 1: Diagram of fluorometric assay for quantification of phagocytosis and actin polymerization. After the cells are plated, treated, and allowed to adhere, particles labeled with green fluorescence (FITC) are added to the cells for phagocytosis. Reaction is stopped by trypan or antibiotic wash (to eliminate non-internalized particles) and fixation is performed with 4% paraformaldehyde. Cells are subsequently stained with red fluorescent actin label (rhodamine phalloidin) and blue fluorescent label (DAPI). Indexes of phagocytosis and actin polymerization are quantified as a ratio of relative fluorescence units of green/blue (FITC/DAPI), or red/blue (rhodamine/DAPI) fluorescence.
1. Plating and Treating Cells
NOTE: Before beginning, consider running the treatments with at least 4 technical replicates, and include the following controls in the plate layout: cells only (unstained, with DAPI alone or rhodamine alone) and particle only.
- Conduct all addition of reagents in steps 1-4 in a sterile cell culture hood in order to protect the samples from contamination.
- Plate macrophages (murine J774 cell line) in a 96-well plate at 1-5 x 104 cells in 50 μl (per well) of supplemented preheated media. Ideal plates for this method are black 96-well plates with clear or black bottoms, depending on reader capabilities.
- For supplemented media, use 10% heat inactivated FBS to avoid interference of complement cascade, and 1% penicillin/streptomycin (if not using live bacteria for phagocytosis). If using adherent cells allow 1-2 hr for adhesion, and for non-adherent cells, spin down the plate with cells for 10 min at 500 x g.
- Add desired agonists/antagonists or vehicle control at 2x concentration (resuspended in PBS or more preferably, in supplemented media) in 50 μl for a final volume of 100 μl (1x) and incubate for a desired amount of time. Multichannel pipette and reagent reservoirs will facilitate fastest processing.
2. Opsonization of Fluorescent Particles

Table 1: Tested cell types and fluorescent particles in fluorometric analysis of phagocytosis and actin polymerization. Table 1 illustrates combinations of cell types (cell lines, and primary cells) that our group has used via the fluorometric method. Other than looking at phagocytosis and actin polymerization by wild type cells, we have also utilized some of these particles for examination of phagocytosis by transfected cells expressing a GFP (green fluorescent protein). Fluorescence of the cells transfected with plasmids containing fluorescent reporters can also be used as a cellular marker in addition to DAPI. Table legend: A – actin polymerization; P – phagocytosis; PT – phagocytosis by GFP labeled transfected cells; OPDex – opsonized dextran bead; HKOP – heat killed opsonized.
- Ensure that fluorescent particles and labels are protected from light at all times, during processing, washing, as well as during the incubation in order to prevent photobleaching
- For dry particles, weigh out fluorescent heat killed opsonized (HKOP) bacteria (E2861: E. coli K-12 strain), using manufacturer’s specifications. Take care to make sure the mass accounts for 10:1 – 20:1 bacteria:cell ratio (or at least a 10:1 ratio – commonly used in quantification of phagocytosis)13,14. Number of particles can be provided by the manufacturer or determined via couting serial dilutions of 1 mg/ml stock solution.
- Resuspend in 50 μl PBS and vortex for 1 minute at highest setting. Ensure proper vortexing (here and throughout the protocol) as the particles tend to aggregate which may affect efficacy of opsonization.
- For particles in suspension such as fluorescently labeled dextran beads (DEX beads), vortex the stock solution for 1 min, take 50 μl of suspension or depending on the bead concentration in the volume determine mount of beads to facilitate 10:1 bead:cell ratio.
- Add an equal volume of the opsonizing reagent to the particles and vigorously vortex for another minute.
- Incubate the particles with opsonizing reagent at 37 °C for 1 hr.
- Following the incubation, centrifuge the particles for 5 min at 500 x g, and remove the supernant containing excess opsonizing reagent. Wash the particles by adding 100 μl of PBS, vortex to resuspend the pellet, and centrifugate for 5 min at 500 x g.
- Repeat these steps two times to assure proper opsonization and removal of unbound antibody.
- Prepare the working solution of opsonized particles by resuspending them in 5 ml of media (sufficient for one 96-well plate) and store away from light until addition to the cells.
3. Phagocytosis
- Place the working solution of particles in a sterile reagent reservoir. From here, add 50 μl of opsonized particles to the cells prepared in step 1 (for a final concentration of 50 ng/ml) and allow phagocytosis to occur at standard cell culture conditions. Illustrated below are two methods of phagocytosis which can be assessed using this method (Diagram 2).

Diagram 2: Comparison of continuous versus synchronize phagocytosis. Continuous phagocytosis indicates continuous internalization of particles over time as they slowly reach the cells on the bottom of the well. A contrasting synchronization step (centrifugation) forces the particles to sink to the bottom, enhancing the particle contact with the cell, and leading to immediate internalization by the cells. Synchronized phagocytosis is a quicker process which more rapidly internalizes particles due to the increased cell:particle contact.
-
- For the continuous phagocytosis method (Diagram 2) that allow cells to continuously phagocytose opsonized bacteria as they slowly sink to the bottom and come in contact with cells, utilize longer time frames to allow for the bacteria to reach the bottom of the well and get in touch with the cells. If conducting a timecourse experiment, add bacteria at different time intervals and stop the entire plate of reactions at the same time to enhance the speed of processing.
- For the synchronized phagocytosis method (Diagram 2) that enhances particle: cell contact via centrifugation, spin down the entire plate containing cells and particles (5 min at 500 x g) as soon as the particles are added, to facilitate synchronization of all time points and to expedite the particle: cell interaction.
- Measure the time course starting right after the completion of the centrifugation step. If conducting a time course experiment, stop each time point independently at different time intervals as described below.
NOTE: The synchronized phagocytosis method is somewhat more laborious from the previous one, but provides higher phagocytic index in a shorter time.
4. Stopping Phagocytosis
- For adherent cells, remove the supernatant and add 50 μl of diluted trypan blue with PBS (1:2 dilution), in order to extinguish fluorescence of non-internalized bacteria.
- For non-adherent cells, spin down the plate and remove supernatant. Add 50 μl of diluted trypan blue. Ensure to centrifuge the plate at (5-10 min at 500 x g) between each wash in order to minimize the cell loss and enable adequate washing.
- Following trypan incubation, wash cells two times with PBS to remove any residual trypan (until the PBS removed from the well turns clear). Trypan has been shown to quench fluorescence15,16 of non-internalized fluorescently labeled, heat killed, bacterial particles, but not for live bacteria or OPDex beads. For live bacteria, include a wash with antibiotics such as penicillin, streptomycin, or gentamicin instead of the trypan wash to avoid confounding effects of fluorescence from non-internalized particles.
- Fix cells with 100 μl of 4% paraformaldehyde, and incubate for 15 min at room temperature, then wash with PBS (at 100 μl per well for each wash). At this step, save the cells at 4 °C for up to a week, or proceed directly to staining and quantification.
NOTE: All the subsequent steps form this point foreword can be done outside of the cell culture hood. - Remove all liquid and add 50 μl of DAPI at 5 ng/ml reconstituted in PBS. Allow 5 min for staining, and then wash twice with PBS. Following the wash step, add 50 μl of PBS to each well and record phagocytosis.
5. Actin Staining
NOTE: In addition to phagocytosis, fluorometric method will enable quantification of actin polymerization by measuring intensity of actin, which is reduced during inhibition of lamellipodia, pseudopodiae, and membrane ruffling as a result of treatment with an inhibitor (Figure 1A). This is an optional step if interested in quantification of actin polymerization in addition to phagocytosis. If actin polymerization is the final goal, using fluorescent particles for phagocytosis is optional.
- Following paraformaldehyde fixation (step 4.3), wash the plate twice with PBS at 100 μl per well for each wash. Add 50 μl of 0.1% Triton X-100 in PBS for permeabilization and incubate for -5 min at RT.
- Wash out 0.1% Triton X-100 2 times with PBS (as above), and block with 1% bovine serum albumin (BSA) for 20 min to avoid non-specific binding of the label.
- Following blocking, remove the BSA solution and immediately add rhodamine phalloidin. To prepare the working solution of rhodamine stain, use 100 μl of methanoic solution (provided by the manufacturer) in 5 ml of PBS (sufficient for one plate). From the working solution add 50 μl per well and incubate for 20 min at room temperature.
NOTE: Following the washing steps and DAPI staining as in step 4.4 cells will be ready for quantification.
6. Quantification
- To quantify the phagocytosis using a fluorescent plate reader, record green fluorescence using excitation filter at λ = 488 nm and emission filter at λ = 518 nm, and blue fluorescence using excitation filter at λ = 355 nm and emission filter at λ = 460 nm.
- To quantify the actin polymerization using a fluorescent plate reader, record blue fluorescence via λ = 355 nm and λ = 460 nm (as above), and red fluorescence via excitation filter at λ = 584 nm and emission filter λ = 620 nm.
NOTE: Orbital or central detection is acceptable for recording. Some cell types gather more to the edge of the well than the center, in which case orbital recording can be of advantage. In particular, average of three points around the orbit could also be used to obtain more accurate results. - Divide the total RFU of green or red fluorescence by blue fluorescence readout (from DAPI), depending on the particle label or phagocytic vs. actin quantification (Diagram 1). Due to high plate-to-plate variance, use relative indexes (such as relative to t = 0 control), to best illustrate trends observed and to facilitate more reliable statistical analysis.
Two major ways of quantifying phagocytosis and the subsequent actin polymerization through the use of this protocol is to observe continuous or synchronized phagocytosis.
Microscopic analysis (Figure 1B) illustrates fluorescent images comparable to what the fluorometer is recording (also see Diagram 1). In Figure 1B are red fluorescence of actin, green 268 fluorescence of FITC labels HKOP E. coli, nuclear DAPI stain, and the merged image of all three 269 fluorophores together. This image indicates effective phagocytosis, actin polymerization, and 270 effective trypan quenching of non-internalized particles.
Continuous phagocytosis of opsonized and unopsonized particles is quite comparable as seen in Figure 2A and C respectively, where J774 cells are internalizing dextran beads. Interestingly, actin polymerization following opsonophagocytosis increases, slowing down at later time points (Figure 2B), while the actin polymerization following internalization of unopsonized particles does not increase (Figure 2D). This is important to consider for future studies examining these processes since opsonophagocytosis leads to much greater actin polymerization than internalization of particles that are not opsonized. Neither of the phagocytic methods using dextran particles result in changes in cell viability as was confirmed via the MTT analysis (Figure 2E).
Unlike continuously increasing trends observed during continuous phagocytosis (Figure 2), synchronized phagocytosis of opsonized dextran beads has a bell shaped trend (Figure 3A), showing reduction in internalization following 30 min. This is to be expected considering processing time and nature of the protocol. This trend is also confirmed by the actin polymerization data (Figure 3B) of these cells where the highest polymerization time point is early on in the timecourse, while at 30 min polymerization returns to baseline. Although more labor intensive, this method has a value of examining phagocytosis right after the synchronization, therefore examining immediate phagocytic effects.
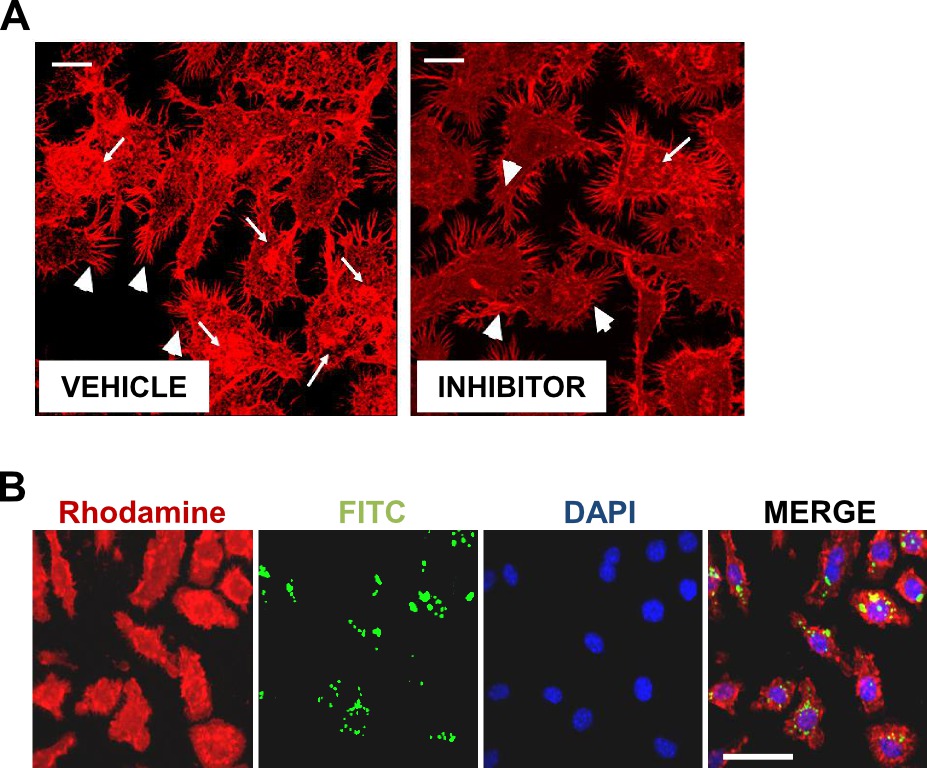
Figure 1: Confocal microscopy of phagocytosis and actin polymerization. Cells were allowed to internalize FITC conjugated OPDex bead (green) at 20:1 bead:cell ratio for 60 min, then cells were washed with trypan and PBS, fixed with paraforlmaldehyde, permeabilised with acetone, and stained for f-actin using rhodamine phalloidin (red), and DAPI (blue). Images illustrate confocal microscopic analysis at 60X with additional digital magnification; (A) white bar = 10 µm (B) white bar = 50 µm. (A) Illustrates changes in actin polymerization following addition of inhibitor. Arrowheads indicate pseudopodia formation, and arrows indicate changes in membrane ruffling. Figure 1A has been modified from Ninkovic and Roy12. Please click here to view a larger version of this figure.

Figure 2: Fluorometric analysis following continuous phagocytosis. J774 murine macrophages were plated onto a 96-well plate and exposed to dextran beads at varying amounts of time, starting with 90, 60, 45, 30, and 15 min. Beads were added at different times and the phagocytic processes for all the reactions were stopped at the same time. Cells were washed with trypan, fixed, stained, and analyzed for phagocytosis of opsonized (A) or unopsonized (C) dextran particles and actin polymerization associated with each (B, D respectively). (E) MTT assay was conducted immediately following phagocytosis at different time points. This figure has been modified from Ninkovic and Roy12.

Figure 3: Fluorometric analysis following synchronized phagocytosis. FITC labeled OPDex beads were added to J774 murine macrophage cells plated in 96-well plates at a cell density of 104 cell/well. After a 30 min incubation the plate was centrifuged at 500 x g for 5 min. Phagocytosis was stopped one at a time, at time points stated in the x-axis by trypan wash, followed by PBS washes, paraformaldehyde fixation and actin staining. This figure has been modified from Ninkovic and Roy12.
