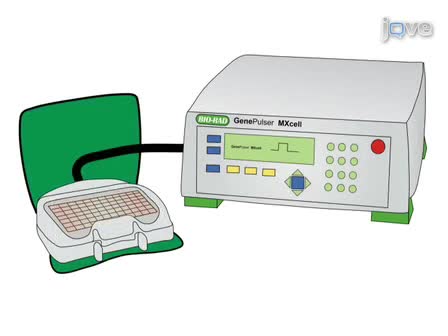
A phage display library was used to identify peptide sequences that target bone. The objective was to investigate the effect of these peptides on mesenchymal cell differentiation and to determine their effect on bone regeneration.