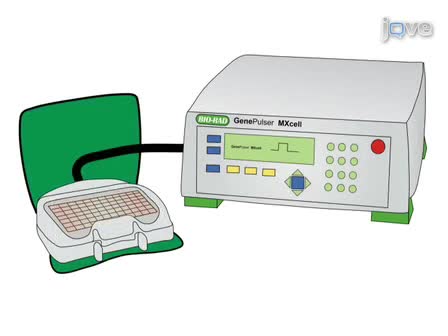
A method of gene transfer into chicken embryos at later incubation stages (older than Hamburger and Hamilton stage (HH) 22) is described. This method overcomes disadvantages of in ovo electroporation applied to older chicken embryos and is a useful technique to study gene function and regulation at older developmental stages.