

Summary
Myt tillåter känslig detektion av transienta och stabil interaktioner mellan proteiner som uttrycks i modellen organismen Saccharomyces cerevisiae. Det har med framgång använts för att studera exogena och jäst integrerade membranproteiner i syfte att identifiera sina samverkande partner i en hög genomströmning sätt.
Abstract
Den grundläggande biologiska och kliniska betydelsen av integrerad membranproteiner drev fram utvecklingen av en jäst-baserat system för hög genomströmning för identifiering av protein-protein interaktioner (PPI) för fullängds transmembrane proteiner. För detta ändamål utvecklat vårt labb split-ubiquitin baserade Membrane Jäst Två Hybrid (MYT) system. Denna teknik möjliggör känslig detektion av övergående och stabil protein interaktioner med
Protocol
1. Bakgrundsinformation
Protein-protein interaktioner (protonpumpshämmare) är de grundläggande byggstenarna inblandade i styr alla cellulära processer. Därför är det viktigt att alla interaktioner hårt regleras för att upprätthålla cellulär homeostas, som en förändring i denna biologiska jämvikt ofta spelar en roll i sjukdom och cancer cell transformation. Membran associerade proteiner är bland de biologiskt viktiga klass av proteiner som de kan initiera komplex signalering kaskader, och förmedlar både import och export av olika molekyler, inklusive läkemedel, som har varit de senaste betydelse inom sjukvården som problem relaterade till läkemedelsresistens har blivit allt vanligare. Att få insikt i komplexiteten av detta protein klass kräver identifiering av de samverkande partner. Att upptäcka sådana partners har visat utmanande, eftersom det ofta kräver hårda villkor som måste optimeras för varje membranbundna protein [1].
Den grundläggande biologiska och kliniska betydelsen av integrerad membranproteiner drev fram utvecklingen av en jäst-baserat system för hög genomströmning identifiering av PPI för full längd transmembrane proteiner. För detta ändamål har vi utvecklat split-ubiquitin baserade Membrane Jäst Två Hybrid (MYT) system [2-4]. Detta verktyg möjliggör känslig detektion av övergående och stabil proteininteraktioner. Det har med framgång använts för att studera exogena respektive endogena proteiner som uttrycks i modellen organismen Saccharomyces cerevisiae [3-7]. Myt utnyttjar iakttagelsen att ubiquitin kan delas upp i två beståndsdelarna:. C-terminal (C UB) och N-terminal (NubI) in vivo-studier har visat att dessa beståndsdelarna spontant rekonstruera på grund av deras höga affinitet till en annan (Figure. 1a). Men att införa en isoleucin 13 till glycin punktmutation i N-terminal hälften av ubiquitin (som producerar ett fragment kallas N UB G) förhindrar detta spontana re-föreningen [8] (Figure. 1b).
Vi använder denna princip på myten systemet (Figure. 1c och 2). Kortfattat är integrerad membran bete protein smält till en C UB fraktion som är kopplad till en konstgjord transkriptionsfaktor som består av Escherichia coli DNA-bindande protein Lexa och aktivering domän VP16 från herpes simplex virus. Bytesdjur genereras av fusion av cDNA eller genomiska DNA-deriverade fragment till NubG fraktion. En interaktion mellan bete och proteiner byte i en jäst värd leder till återupprättandet av en fullängds "pseudo-ubiquitin" molekyl, med efterföljande erkännande av cytosoliskt deubiquitinating enzymer (dubbar) och proteolytiska frisläppande av transkriptionsfaktor. Den transkriptionsfaktor kan ange sedan cellkärnan, och aktivera ett system reporter gen (i allmänhet innebär uttrycket för HIS3, lacZ och ADE2 gener) som möjliggör tillväxt av jäststammar på selektiva media, vilket tyder på bete och bytesdjur interaktion [ 2-4].
Identifiering och karakterisering av integrerad samverkan membranprotein kommer att ge information för att fullt ut förstå deras funktion. Som vi mer exakt förstå och dissekera de roller de proteiner som interagerar med integrerad membranproteiner, kan vi få inblick i det dynamiska samspelet involverade i regleringen av dessa proteiner och upptäck nya mål som kan ha terapeutiska potential.
2. Val av bete och lämplig MYTH System
- Innan genomföra MYTEN analys, kontrollera att din protein har sin N-och / eller C-terminus i cytosolen av cellen. Det är viktigt att C UB-Lexa-VP16 taggen smält till ditt protein vid en sådan terminus, eftersom deubiquitinating enzymer som behövs för transkriptionsfaktor release är beläget i cytosolen [4].
- Därefter avgöra vilket av de två stora varianter av myt är lämplig. För icke-infödda jäst proteiner, kan traditionella myten (tMYTH) användas, där beten är överuttryckt ectopically från en plasmid. För infödda jäst proteiner, är integrerad MYTEN (iMYTH) Metoden för val. I iMYTH beten är endogent taggade med C UB-Lexa-VP16 tag, lämnar dem under kontroll av sitt eget promotor. Detta är en fördel, eftersom vildtyp uttryck nivå beten hjälper till att eliminera problem som är förknippade med protein överuttryck, såsom ett ökat antal falskt positiva [3]. Med undantag av den initiala bete byggandet och den selektiva media som används, är båda formerna av myt som utförs i en i huvudsak identisk sätt. För tydlighetens skull kommer vi att fokusera främst på användningen av tMYTH i den här rapporten, eftersom denna variant kan i princip användas med membranproteiner från praktiskt taget alla organismer och ärdärmed mer allmänt tillämpliga.
3. Bete Generation och validering
- Krävs medier och lösningar
- Sterila DDH 2 O förberett genom autoklavering vid 121 ° C, 15 psi i 30 minuter.
- 3-Amino-1 ,2,4-triazol (3-AT) lösning som beretts enligt en 1M stamlösning i DDH 2 O. Sterilisera genom passage genom ett 0,2 ìm filter.
- YPAD Tillväxt Media bestående av 1% w / v jästextrakt, 2% w / v pepton, 2% w / v glukos och 100 mikroM adenin upprättats i DDH 2 O. Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- 10x aminosyror / Nucleotide Base Mix: Den kompletta mixen innehåller 1,0 mm adenin, 1,8 mm uracil 1,0 mm arginin, 1,0 mm Histidin, 2,3 mM Isoleucin, 7,6 mM Leucin, 1,6 mm Lysin, 10,1 mm metionin, 3,0 mm Fenylalanin, 16,8 mm treonin, 2,0 mm tryptofan, 1,7 mm Tyrosin och 12,8 mm Valin, upprättad i DDH 2 O. För avhopp utelämna nödvändig aminosyra (er) och / eller nukleotid bas (s). Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- Syntetiska Drop-Out (SD) Tillväxt Media består av 0,67% w / v jäst kväve bas (utan aminosyror, men med ammoniumsulfat), 2% w / v glukos, 2% w / v agar och 1x aminosyror / Nucleotide Mix, upprättats i DDH 2 O. Förbered både flytande och fasta (innehållande 2% agar) SD-Leucin media. Också förbereda solida SD-tryptofan-Leucin och SD-tryptofan-Leucin-Adenin-histidin media. Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter. Häll solida material i 100x15 mm petriskålar.
- Syntetiska Drop-Out (SD) Tillväxt media som innehåller 3-AT. Förbered SD-tryptofan-Leucin-Adenin-histidin media som beskrivs, men innehåller 3-AT i koncentrationer på 25, 50, 75 och 100 mm. Tillsätt lämplig mängd av 3-AT från 1M sterila stamlösning till media efter att den har autoklaveras och kylda (men ännu inte stelnat). Häll i 100x15 mm petriskålar.
- PEG / Litium Acetat Mix består av 40% w / v PEG-3350, 120 mm Litium acetat och 167 mikrogram / ml Lax spermiens DNA (typ III natriumsalt) upprättad i DDH 2 O. För att säkerställa steriliteten av denna blandning förbereda det från sterilt vatten och lösningar (dvs. autoklaveras 50% PEG-3350, autoklaveras 1M Litium Acetat och 2 mg / ml Lax spermiens DNA typ III natriumsalt upprättats i sterila DDH 2 O).
- Enzym och reagenser för att bedriva PCR.
- Kommersiella miniprep kit.
- Kalk glaspärlor (0,5 mm).
- Behöriga Escherichia coli-celler som lämpar sig för plasmid förökning (t.ex. DH5α, XL10 guld) och standard medier som lämpar sig för bakteriell förökning och plasmid urval.
- Särskilda jäststammar, plasmider och grundfärger som beskrivs i protokollet.
- Generering av tMYTH balter genom Gap Reparation
- Betet måste klonas in i en lämplig vektor för taggning och uttryck. En mängd olika tMYTH vektorer är för närvarande tillgängliga för användning i beten konstruktion. Vektorer som pCMBV4, pAMBV4 och pTMBV4 tillåter byggandet av C-terminalt taggade beten (bete-C UB-Lexa-VP16) under kontroll CYC1 (svag), ADH1 (stark) och TEF1 (mycket stark) initiativtagare respektive . N-terminalt taggade beten (Lexa-VP16-C UB-bete) kan genereras med vektorer som pTLB-1 och pBT3-N, under kontroll av de TEF1 och CYC1 initiativtagare, respektive. Vektorn Valet beror på bete och måste bestämmas empiriskt. I vissa fall högre bete uttryck är nödvändig för att upptäcka interaktioner, medan det i andra fall bete överuttryck faktiskt kan vara skadlig, vilket leder till ett ökat antal falskt positiva.
- Begränsning smälta den valda plasmiden på lämplig begränsning plats (er). Cleavage endast ske i omedelbar närhet av C UB-Lexa-VP16 tagg (före taggen för C-terminal märkning eller nedströms för N-terminal taggning). Till exempel när du använder pAMBV4 vektorn är SfiI ett perfekt val. Förvara rötas plasmiden vid -20 ° C tills produkten ska användas.
- Design primers för amplifiering och kloning av genen av intresse. Den 5 "slutet av din Framåt Primer måste matcha cirka 35-40 nukleotider uppströms begränsningen platsen, medan 3 'änden måste matcha de första 18-20 nukleotider i målgenen. Den 5 "slutet av omvänd Primer måste matcha omvända komplement cirka 35-40 nukleotider nedströms från begränsningen plats, med 3" slutet matchar omvända komplement till de sista 18-20 nukleotider i målgenen (utan stopp kodon om C UB-Lexa-VP16-tagg läggs vid C-terminus). Beroende på om N-eller C-terminal märkning utförs, välj 35-40 nukleotider för framåt eller bakåt primer så att målet genen är klonad i ram med C UB-Lexa-VP16 tag.
- Förstärk genen av intresse med PCR med hjälp av ovanstående grundfärger. PCR-parametrar kommer att bero på särskilt enzym och specifika används primers.
- Förvandla PCR-produkten tillsammans med de uppdelade plasmiden i ett lämpligt jäst lab stam med en leu2 mutation (t ex BY4741). En myt reporter stam (t.ex. THY.AP4 eller L40) kan användas men är inte nödvändigt, eftersom syftet med jästen på denna punkt är helt enkelt att fungera som en miljö där gapet reparation homolog rekombination kan förekomma. Omvandling bör ske enligt följande:
- Inokulera en enda koloni av valda jäst sila upp i 5 ml steril YPAD media och inkubera över natten vid 30 ° C med konstant skakning (200 rpm).
- Späd natten kultur i 50 ml rent YPAD media till en OD600 på ~ 0,15 och inkubera vid 30 ° C med skakningar (200 rpm). Växa under cirka 3-4 timmar tills en OD600 på ~ 0,6 uppnås.
- Centrifugera cellerna vid 700xg i 5 minuter och avlägsna supernatanten.
- Resuspendera cellpelleten i 25 ml sterila DDH 2 O och centrifugera vid 700xg i 5 minuter.
- Avlägsna supernatanten och resuspendera cellpelleten i 1 ml steril DDH 2 O.
- Tillsätt 100 mikroliter av cellerna, 300 mikroliter PEG / Litium Acetat mix, och smält plasmid (50 fmol) och PCR-produkten (250-500 fmol) till ett mikrofugrör.
- Inkubera vid 30 ° C i 30 minuter.
- Värme chock vid 42 ° C i 1 timme.
- Centrifugera vid 3000xg i 5 min och avlägsna supernatanten.
- Resuspendera cellpelleten i 200 mikroliter sterila DDH 2 O och platta hela volymen på solida, SD-Leucin selektiva medier. Växer vid 30 ° C i 2-4 dagar.
- Väx upp en enda koloni av det transformerade stam i 5 ml SD-Leucin flytande medier vid 30 ° C över natten.
- Centrifugera cellerna vid 700xg i 5 minuter och avlägsna supernatanten.
- Isolera bete plasmid DNA från cellens pelletsen med hjälp av något kommersiellt miniprep kit. Följ standardprotokoll med en modifiering. För att säkerställa tillräcklig lys jäst cell, tillsätt en liten mängd 0,5 kalk mm soda glaspärlor till pellets efter den första resuspension och vortexa kraftigt i 5 minuter. Fortsätt sedan med den kommersiella protokoll som vanligt.
- Förvandla isolerade jäst-DNA till en behörig E. coli stam som lämpar sig för plasmid förökning (t.ex. DH5α, XL10 guld) med en omvandlingseffektiviteten minst 1x10 7 celler / mikrogram DNA. Observera att bete plasmider kan väljas för att använda kanamycin.
- Harvest plasmid DNA från omvandlas E. coli med hjälp av en standardmetod för isolering av DNA eller kommersiella kit.
- Kontrollera tolkas betet plasmiden av sekvensering.
- Omvandla den verifierade bete bygga in en lämplig MYTEN reporter stam (t.ex. THY.AP4, L40). Jästen omvandlingen Protokollet just beskrivit kan användas, ersätta betet plasmid-DNA i stället för smält plasmiden och PCR-produkt.
- Bait Validering - Rätt lokalisering
- Före användning, är bete stammar analyseras för att säkerställa att de är korrekt lokaliserade till jästen membranet. När du använder iMYTH kommer denna lokalisering beror specifikt på de egenskaper taggade bete. För tMYTH, bete plasmider innehåller i allmänhet en signal sekvens (t.ex. Matα) styra uttryckt proteinet till plasmamembranet. Lokalisering bestäms med hjälp av fluorescensmikroskopi. Införande av en YFP molekyl i betet tagg sekvensen (dvs. C UB-YFP-Lexa-VP16) är den enklaste och mest direkta metoden, vilket direkt visualisering av levande celler, och är vanligt i iMYTH. Alternativt kan en standard immunofluorescens tillvägagångssätt med antikroppar mot Lexa eller VP16 komponenter taggen användas.
- Bait Validering - N UB G / I kontrolltest
- När korrekt lokalisering av betet har fastställts, är det nödvändigt att se till att betet inte aktivera reportern systemet ensam eller i närvaro av icke-samverkande bytesdjur (dvs kontrollera att betet inte är själv-aktivering). Detta sker med N UB G / I Test, där betet omvandlas med samverkande (positiv) och icke-samverkande (negativ) bytesorganismer kontroll, och tillväxten bedöms selektiva medier. En bete måste växa på selektiva medier in närvaro av den positiva kontrollen, och inte växa i närvaro av den negativa kontrollen, för att vara lämplig för användning i myt.
- Börja med att omvandla bete stam som skall undersökas med 100-200 ng kontroll bytesdjur plasmid. Den tidigare beskrivna jästen omvandling protokollet kan användas, ersätta SD-Leucin media i stället för YPAD och använda SD-tryptofan-Leucin media för det sista plätering steget. Följande kontroll byte konstruktioner är vanliga:
- pOST1-N ub I (positiv kontroll)
- pOST1-N ub G (negativ kontroll)
- pFUR4-N ub I (positiv kontroll)
- pFUR4-N ub G (negativ kontroll)
- OST1 är en del av oligosaccharyl komplexa och är lokaliserad till endoplasmatiska retiklet membranet [9] medan FUR4 är en uracil permease och är lokaliserad till plasmamembranet [10]. Även om dessa proteiner är generellt vårt första val för användning som icke-samverkande bytesorganismer, kommer deras lämplighet varierar från fall till fall. I den osannolika händelsen att din bete förutspås att verkligen interagera med båda dessa kontroller, kommer alternativa bytesdjur måste väljas. Minns att N UB jag är vildtyp form av N-terminalen av ubiquitin, och spontant interagerar med C UB oberoende av ett samspel mellan proteiner som C UB och N ub är smält. Således N ub jag bytesdjur utgör positiva kontroller, medan N UB G bytesdjur (bära Isoleucin 13 till Glycin mutation som hindrar spontan sammanslutning av N UB och C UB) tjäna som negativa kontroller.
- Resuspendera enstaka kolonier av varje omvandlas bete till 100 mikroliter sterila DDH 2 O.
- Seriellt späda den suspenderade celler i sterila DDH 2 O att producera utspädning på 1 / 10, 1 / 100 och 1 / 1000.
- Spot 5 mikroliter volymer före och efter utspädning celler på SD-tryptofan-Leucin och SD-tryptofan-Leucin-Adenin-histidin media med och utan 3-AT i en serie koncentrationer. 3-AT fungerar som en kompetitiv hämmare av HIS3 reporter genen, och tjänar till att öka stringensen i urvalsprocessen. Det kan vara bra i vissa fall för att hämma den icke-specifika tillväxt av svagt till måttligt själv aktivera beten.
- Låt fläckarna torka och sedan Inkubera plattorna vid 30 ° C i 2-4 dagar.
- Alla transformants ska växa på SD-tryptofan-Leucin plattor, vilket tyder på att de framgångsrikt har förvandlats med byte plasmid. Beten som inte själv aktivera kommer att växa på SD-tryptofan-Leucin-Adenin-histidin media först när omvandlas med N ub jag bytesdjur konstruerar, och inte med N ub G bytesorganismer. Notera vilken koncentration av 3-AT krävs i media (om någon) eftersom detta kommer att behöva användas vid screening.
4. Screening
- Krävs medier och lösningar
- Sterila DDH 2 O förberett genom autoklavering vid 121 ° C, 15 psi i 30 minuter.
- 0,9% NaCl-lösning upprättats i DDH 2 O och steriliseras genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- Natriumfosfat lösning som består av 493 mm Dinatriumvätefosfat och 250 mm natrium monobasiskt i DDH 2 O. Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- X-Gal (5-bromo-4-klor-3-indolyl-β-D-galaktopyranosid) lösning som beretts enligt en 100 mg / ml stamlösning i N, N-dimetylformamid.
- 2xYPAD Tillväxt Media innehållande 2% w / v jästextrakt, 4% w / v pepton, 4% w / v glukos och 100 mikroM adenin, upprättad i DDH 2 O. Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- Syntetiska Dropout (SD) Tillväxt Media, bereds enligt anvisningarna tidigare. Förbered flytande SD-Leucin och fasta SD-tryptofan-Leucin. Häll den fasta materialet i både 100x15 mm petriskålar. Förbered fast SD-tryptofan-Leucin-Adenin-histidin media i 150 mm runda tallrikar, 16 tallrikar för varje skärm, som innehåller 3-AT, om nödvändigt, vid koncentrationen bestämmas ur N UB G / I kontrolltest.
- Syntetiska Dropout (SD) media + 5-Bromo-4-klor-3-Indoyl-β-D-galaktopyranosid (X-Gal). Förbered SD-tryptofan-Leucin-Adenin-histidin media som innehåller agar som beskrivits tidigare. Efter autoklavering, låt svalna, tillsätt 3-AT (om nödvändigt) följt av 1:10 volym of sterila natriumfosfat Solution. Lägg sedan till X-Gal-lösning till en slutlig koncentration på 80 mikrogram / ml. Blanda väl och häll upp i 150 mm runda plattor.
- PEG / Lithium Acetat lösning II innehåller 40% PEG-3350, 100 mm Litium Acetat, 1 mM EDTA och 10 mM Tris pH 7,5. Bered denna lösning med hjälp av sterila DDH 2 O och lösningar (t.ex. autoklaveras 50% PEG-3350, 1 M Litium Acetat, 100 mM Tris pH 7,5 och 500 mM EDTA pH 8,0).
- Litium Acetat / Tris EDTA-lösning som innehåller 110 Acetate mM Litium, 11 mM Tris pH 7,5 och 1,1 mM EDTA. Bered denna lösning med hjälp av sterila DDH 2 O och lösningar (t.ex. autoklaveras 1M Litium Acetat, 100 mM Tris pH 7,5 och 500 mM EDTA pH 8,0).
- 10x Tris EDTA-lösning som består av 100 mM Tris pH 7,5 och 10 mM EDTA upprättats i DDH 2 O. Sterilisera genom autoklavering vid 121 ° C 15 psi i 30 minuter.
- Enkelsträngat Carrier DNA (ssDNA) Lösning innehållande 2 mg / ml Lax spermiens DNA Typ III natriumsalt, upprättad i sterila DDH 2 O.
- Kommersiella miniprep kit.
- Kalk glaspärlor (0,5 mm).
- Behöriga Escherichia coli-celler som lämpar sig för plasmid förökning (t.ex. DH5α, XL10 guld) och standard medier som lämpar sig för bakteriell förökning och plasmid urval.
- Särskilda jäststammar och plasmider som beskrivs i protokollet
- Large Scale Transformation
- Inokulera en enda koloni av myten reportern stam som innehåller ditt bete i 5 ml SD-Leucin media och inkubera över natten vid 30 ° C med skakningar (200 rpm).
- Späd natten kultur i 200 ml SD-Leucin media till en OD600 = 0,15 och inkubera vid 30 ° C med skakningar (200 rpm). Växa fram till OD600 = 0,6 till 0,7 (ca 4-5 timmar).
- Strax innan målet OD600 nås, tina en alikvot av ssDNA lösning. Koka i 100 ° C i 5 minuter och kyl sedan på is. Upprepa en gång.
- När målet OD600 har uppnåtts, skörda celler via centrifugering vid 700xg i 5 minuter (dela upp 200 ml kultur mellan 4x50 mL skruvkork centrifugrör).
- Tvätta varje pellet med 30 ml steril DDH 2 O och kort virvel provet. Centrifugera vid 700xg i 5 minuter.
- Kassera supernatanten och återsuspendera varje pelleten i 1 ml Litium Acetat / Tris EDTA-lösning. Överför till en steril 1,5 ml mikrofugrör och centrifugera vid 700xg i 5 minuter.
- Kassera supernatanten och återsuspendera varje pellet i 600 mikroliter av Lithium Acetat / Tris EDTA-lösning.
- Lägg till följande i 4x15 mL skruvkork centrifugrör:
- 2,5 ml PEG / Lithium Acetat lösning II
- 600 mikroliter återsuspenderade celler
- 100 mikroliter ssDNA lösning
- 7 mikrogram av bytesdjur bibliotek DNA
- Biblioteken innehåller bytesdjur taggade antingen N-eller C-terminus med N UB G, och framställs av en rad olika cDNA eller genomiska källor, finns kommersiellt tillgängligt ( www.dualsystems.com ). De specifika bibliotek som används måste avgöras från fall till fall, beroende på vald bete och experimentella mål.
- Vortexa rören i 1 minut för att säkerställa omsorgsfull blandning och sedan Inkubera i 30 ° C vattenbad under 45 minuter. Blanda kort var 15 minut.
- Tillsätt 160 mikroliter dimetylsulfoxid (DMSO) till varje rör och blanda genast genom att vända rören.
- Inkubera i 42 ° C vattenbad under 20 minuter.
- Samla transformants genom centrifugering vid 700xg i 5 minuter.
- Kassera supernatanten. Recover transformants genom resuspension av varje pellet i 3 ml 2xYPAD. Pool alla prover i ett enda 50 mL skruvkork centrifugrör.
- Inkubera vid 30 ° C i 90 minuter för cell återhämtning.
- Centrifugera vid 700xg i 5 minuter och kassera supernatanten.
- Resuspendera cellen pellets i 4,9 ml steril 0,9% NaCl.
- Använda 100 mikroliter återsuspenderad celler förbereda 10-faldig seriespädningar i steril 0,9% NaCl från 10x till 1000x.
- Plate 100 mikroliter av 100x och 1000x spädningar på SD-tryptofan-Leucin medier och inkubera vid 30 ° C i 2-3 dagar. Dessa skyltar fungerar som en kontroll och används för att beräkna effektiviteten i omvandlingen.
- Lika delar resterande 4,8 ml återsuspenderad celler och platta på stora (150 mm) SD-tryptofan-Leucin-Adenin-histidin plattor,innehåller den nödvändiga mängden av 3-AT som bestäms i N UB G / I-provning, och inkubera vid 30 ° C i 3-4 dagar.
- Resuspendera enstaka kolonier (med vardera celler som innehåller ett potentiellt interagerande bete-byte par) i 100 mikroliter 0,9% NaCl och platta 5 mikroliter alikvoter på SD-tryptofan-Leucin-Adenin-histidin + X-Gal medier (och inklusive 3-AT om krävs). Låt växa i 2-4 dagar. Detta steg fungerar som en andra omgång av selektiv undersökning, och hjälper till att avlägsna falska positiva erhållits i den första rundan. Endast kolonier som visar stark tillväxt och en blå färg väljs ut för vidare analys.
- Prey DNA Isolering och Sekvensering
- Isolera plasmid DNA från den blå jästen kolonier med hjälp av en miniprep protokoll med de ändringar som beskrivits tidigare. Var noga med att växa upp cellerna i SD-Tryptofan media endast välja för lagring av byten, men inte bete, plasmider. För skärmar som ger ett mycket stort antal träffar, kan ett kommersiellt tillgängligt hög genomströmning miniprep kit vara en fördel på denna punkt.
- Förvandla isolerade jäst plasmid-DNA till en behörig E. coli stam som lämpar sig för plasmid förökning (t.ex. DH5α, XL10 guld) med en omvandlingseffektiviteten minst 1x10 7 celler / mikrogram DNA. Observera att byte plasmider kan väljas för att använda ampicillin.
- Isolera plasmid DNA från omvandlas E. coli med hjälp av en standardmetod för isolering av DNA eller kommersiella kit. Återigen kan en hög genomströmning miniprep kit vara användbart om provet antalet är stort. Förstärkningen av DNA i E. coli ökar kraftigt plasmiden avkastning och säkerställer att en tillräcklig mängd DNA finns både för sekvensering och vidare analys.
- Sekvens de isolerade plasmider med en primer komplement till sekvens inom N ub G.
- Sammanställa och analysera alla sekvensering data för att montera din preliminära lista över interactmedlemmar. Detta kan ske manuellt eller på ett automatiserat sätt med hjälp av lämplig mjukvara.
- Bait Dependency Test
- Efter montering av den preliminära listan över interactmedlemmar är det viktigt att åter kontrollera interaktioner och eliminera promiskuösa bytesdjur som interagerar / aktivera reporter systemet på ett sätt som är oberoende av bete identitet. Detta sker med hjälp av Test Agn Dependency. I detta test är alla identifierade interactmedlemmar förvandlas tillbaka till den ursprungliga bete stammen, samt en stam hyser en kontroll konstgjorda beten består av ett enda transmembrane domän smält till C UB-Lexa-VP16 tag. Omvandling sker enligt de standardprotokoll som tidigare beskrivits, med hjälp av SD-Leucin media i stället för YPAD och SD-tryptofan-Leucin fasta medier för den sista plätering steg.
- Resuspendera enstaka kolonier från ovanstående förändringar i 100 mikroliter sterilt DDH 2 O och plats 5 mikroliter volymer på SD-tryptofan-Leucin-Adenin-histidin + X-Gal medier (och inklusive 3-AT om det behövs). Plattorna inkuberas därefter i 2-4 dagar vid 30 ° C. Helst flera transformants bör väljas för varje byte, och både den ursprungliga bete och konstgjorda beten bör upptäckas på samma tallrik.
- Jäst bär den konstgjorda beten och bytesdjur som orsakar aktivering av reportern systemet (dvs. tillväxt och blå färg) anses vara promiskuösa och att specifika bytesdjur tas bort från listan över interactmedlemmar.
- Bytesdjur som orsakar tillväxt och blåfärgning i jäst med agn-av-intresse, men inte den konstgjorda beten, bekräftar just detta samspel. Om däremot jäst hysa bytet och ditt bete-av-intresse växer inte, detta är byte bort från listan över interactmedlemmar.
- Resterande bytesdjur utgör den kompletta listan över interactmedlemmar identifierats i myten skärmen.
5. Ytterligare studier
När MYTEN har slutförts, måste ytterligare analyser göras för att validera och bestämma den biologiska betydelsen av de upptäckta interaktioner. De specifika undersökningar som behöver göras kommer att variera från fall till fall, och måste bestämmas av den enskilde forskaren. Några vanliga exempel på uppföljning omfattar kooperativa immunoprecipitation experiment och studier borttagning i den ursprungliga organismen. Dessutom kan datoriserad analys av de data som erhållits vara användbart för att upptäcka mönster och hjälpa till att identifiera potentiella relevansen och roll olika interaktioner kan spela. Således tjänar MYTEN tekniken som ett kraftfullt första steg "mot att identifiera och förstå de kritiska funktionella interaktioner av membranproteiner. Tillsammans med detaljerad uppföljning studier och andra nyutvecklade och nya technologies, det lovar att bli ett värdefullt verktyg för att låsa upp mysterier av cellen.

Figur 1. Principen om delad ubiquitin. A. Ubiquitin kan delas upp i två beståndsdelarna: C-terminal (C UB) och N-terminal (N ub I). Dessa beståndsdelarna spontant beredning på grund av deras höga affinitet till en annan. b. ETT ub Jag punktmutation i position 13 från en isoleucin till glycin (N ub G) förhindrar detta spontana re-förening. C. I myten systemet är C UB smält till bete-av-intresse (B) och bytet är smält till N UB G (A). AB proteininteraktioner återuppstår pseudo-ubiquitin.
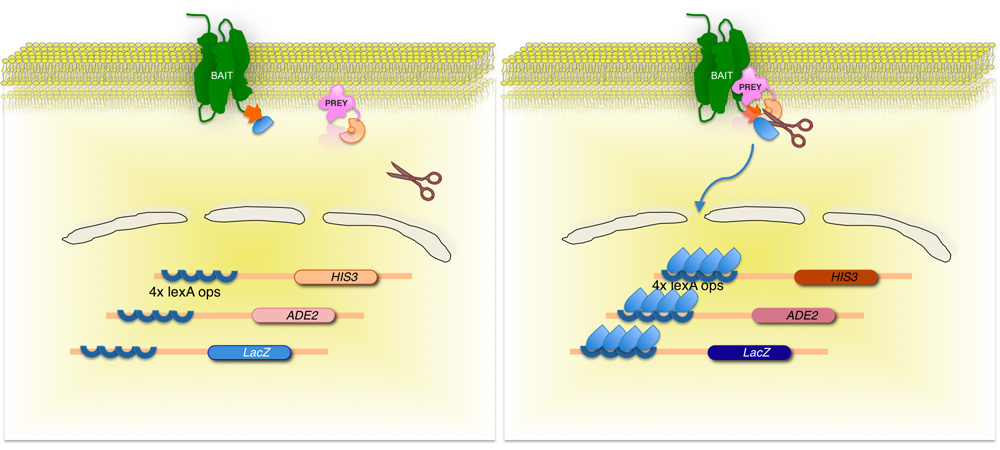
Figur 2. Split-ubiquitin baserade membran jäst två-hybrid (MYT) system.
Den membranprotein-av-intresse (bete) är smält till C-terminala av jäst ubiquitin (C UB), konjugerat till en transkriptionsfaktor. Använda ett cDNA eller gDNA biblioteket är varje protein som kodas av biblioteket (bytet) smält till motsvarande N-terminalen av ubiquitin fraktion (N Ub G). Om de två proteinerna inte samverkar förblir transkriptionsfaktor vid membran-gränssnittet (till vänster). Men om proteiner interagerar de två ubiquitin beståndsdelarna delta, vilket resulterar i klyvning av ubiquitin specifika proteaser. Klyvning släpper transkriptionsfaktor, vilket resulterar i uttryck av reportern gener (högra panelen)

Figur 3. MYT pipeline.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Myt är den första hög genomströmning system som möjliggör identifiering av interaktioner mellan full längd membranproteiner och cytosola eller membranbundna partner. Det har använts för att studera membranproteiner från en rad organismer [3-7]. Det finns dock vissa detaljer som kan behöva granskas för att säkerställa protein-av-intresse är mottaglig att studera med myt.
Många membranbundna proteiner är riktad till plasmamembranet via en signal sekvens som sedan klyvs att producera mogna proteinet. Denna sekvens är organism-specifika och det är möjligt att denna infödda signalsekvensen kommer att orsaka mis-lokalisering av protein-av-intresse när de uttrycks i jäst eftersom signalen sekvensen förblir oredovisade. För att kringgå detta problem, konstruerade vi dessa specifika proteiner som smält till jästen signalsekvensen, härrör från Parning faktor alfa (MATα). Denna peptid sekvens (sk MFα-SS) åter lokaliserar protein jästen plasmamembranet och viktigare, klyvs av peptidaser jäst signal. Denna peptid sekvens finns i plasmider pTMBV-MFα och pAMBV-MFα.
En annan viktig parameter som måste betonas är agn uttryck nivåer. Den projektansvariga som driver uttryck för betet reglerar denna parameter. Det kan krävas att optimera nivåerna bete uttryck med NubG / NubI test och mängden 3-AT nödvändigt att undanröja överuttryck artefakter, där betet är "self-aktivering" (dvs. den minst lovande interagerar med många icke-specifika byten proteiner) . Undersöka jäst proteiner producerar de mest fysiologiskt relevanta bete koncentrationer när iMYTH tillämpas. I detta fall är genen-av-intresse taggade med Cub-TF inom gDNA. Alternativt kan exogena proteiner uttryckas från pBT3-STE och pCMBV plasmider som bär CYC1 promotor vilket resulterar i låga bete uttryck. Den plasmider pTMBV och pTLB1 hamnen TEF1 arrangören medan pAMBV har ADH1 promotor, både som driver starka uttryck för bete protein. Om bete protein nivåer kräver ytterligare optimering kan det vara nödvändigt att använda den pTLB-en plasmid som bär TEF1 promotorn, är dock Lexa DNA-bindande domän muterat i R156G för minskning av de affinitet mot exogena initiativtagarna reporter gen, i slutändan minska sannolikheten för självaktivering [5].
En annan faktor som spelar en viktig roll för MYTEN framgång är biblioteket val används för screening process. Detta beror på den endogena profilerna bete uttryck. Till exempel kan betet uttryckas i specifika vävnader, och därför är det viktigt att använda ett bibliotek som byggs från detta specifika vävnad. Detta kommer att säkerställa fysiologiskt relevanta interaktioner har identifierats.
Det MYTEN Systemet är ett enkelt och snabbt verktyg som ger ett överflöd av information om en klass av proteiner som har varit svåra att studera. Dessa identifierade interaktioner kan stöd i att klarlägga hela biologiska funktionen av membranproteiner.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Igor Stagljar är en av grundarna av DualSystems Biotech, Schweiz.
Acknowledgments
Vi vill tacka Dawn Edmonds för en kritisk läsning av detta manuskript. Den Stagljar labbet stöds av medel från den kanadensiska institutet för innovation (CFI), den kanadensiska Institute for Health Research (CIHR), hjärtat och Stroke Foundation, den kanadensiska Cancerfonden, och Novartis.
Materials
| Name | Company | Catalog Number | Comments |
| Polyethenlene Glycol (PEG3350) | BioShop Canada | PEG335 | |
| Lithium Acetate Bihydrate | BioShop Canada | LIA001 | |
| X-Gal (5-Bromo-4-Chloro-3-Indolyl-b-D-galactopyranoside) | BioShop Canada | XGA001 | |
| N`,N-dimethyl formamide | BioShop Canada | DMF 451 | |
| 3-amino-1,2,4-triazole (3-AT) | BioShop Canada | ATT124 | |
| Sodium phosphate dibasic | BioShop Canada | SPD307 | |
| Sodium phosphate monobasic | Fisher Scientific | BP329-500 | |
| Salmon Sperm DNA | VWR international | CA80601-120 | |
| D-Glucose | BioShop Canada | GLU501 | |
| LB Broth LENOX | BioShop Canada | LBL405 | |
| Yeast Nitrogen Base | BioShop Canada | YNB406 | |
| Yeast Extract | BioShop Canada | YEX401 | |
| Peptone | BD Biosciences | 211677 | |
| Bio-Tryptone | BioShop Canada | TRP402 | |
| Adenine Sulphate | BioShop Canada | ADS201 | |
| L-Uracil | BioShop Canada | URA241 | |
| L-Threonine | BioShop Canada | THR002 | |
| L-Histidine | BioShop Canada | HIS200 | |
| L-Methionine | BioShop Canada | MET222 | |
| L-Valine | BioShop Canada | VAL201 | |
| L-Phenylalanine | BioShop Canada | PHA302 | |
| L-Isoleucine | BioShop Canada | ISO910 | |
| L-Tyrosine | BioShop Canada | TYR333 | |
| L-Leucine | BioShop Canada | LEU222 | |
| L-Arginine | BioShop Canada | ARG006 | |
| L-Tryptophane | Fisher Scientific | BP395-100 | |
| L-Lysine | BioShop Canada | LYS101 | |
| L-Alanine | Fisher Scientific | BP369-100 | |
| Agar | BioShop Canada | AGR001 | |
| Soda Lime Galss Beads | Biospec Products | 11079105 | |
| Sodium Chloride | BioShop Canada | SLD002 |
References
- Stagljar, I., Fields, S. Analysis of membrane protein interactions using yeast-based technologies. Trends Biochem Sci. 27 (11), 559-563 (2002).
- Iyer, K. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci STKE. 275, pl3-pl3 (2005).
- Paumi, C. M. Mapping protein-protein interactions for the yeast ABC transporter Ycf1p by integrated split-ubiquitin membrane yeast two-hybrid analysis. Mol Cell. 26 (1), 15-25 (2007).
- Stagljar, I. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc Natl Acad Sci U S A. 95 (9), 5187-5192 (1998).
- Gisler, S. M. Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol Cell Proteomics. 7 (7), 1362-1377 (2008).
- Scheper, W. Coordination of N-glycosylation and protein translocation across the endoplasmic reticulum membrane by Sss1 protein. J Biol Chem. 278 (39), 37998-38003 (2003).
- Thaminy, S. Identification of novel ErbB3-interacting factors using the split-ubiquitin membrane yeast two-hybrid system. Genome Res. 13 (7), 1744-1753 (2003).
- Johnsson, N., Varshavsky, A. Split ubiquitin as a sensor of protein interactions in vivo. Proc Natl Acad Sci U S A. 91 (22), 10340-10344 (1994).
- Kelleher, D. J., Gilmore, R. The Saccharomyces cerevisiae oligosaccharyltransferase is a protein complex composed of Wbp1p, Swp1p, and four additional polypeptides. J Biol Chem. 269 (17), 12908-12917 (1994).
- Chevallier, M. R. Cloning and transcriptional control of a eucaryotic permease gene. Mol Cell Biol. 2 (8), 977-984 (1982).
