

Summary
Миф позволяет чувствительны обнаружения переходных и стабильного взаимодействия между белками, которые выражаются в модели CEREVISIAE организме Saccharomyces. Она была успешно применена для изучения экзогенных и дрожжи интегральные мембранные белки с целью выявления их взаимодействия партнеров в высоком образом пропускную способность.
Abstract
Фундаментальные биологические и клинические важность интегральные мембранные белки стимулировали развитие дрожжей-системы для высокой пропускной способности идентификации белок-белковых взаимодействий (PPI) за полнометражный белками трансмембранного. С этой целью в нашей лаборатории разработаны сплит-убиквитин основан дрожжей Мембрана два-Hybrid (МИФ) системы. Эта технология позволяет чувствительны обнаружения переходных и стабильных белковых взаимодействий использованием
Protocol
1. Исходная информация
Белок-белковые взаимодействия (ИПП) являются основными строительными блоками участие в управлении всех клеточных процессах. Следовательно, очень важно, чтобы все взаимодействия жестко регулируется в целях поддержания клеточного гомеостаза, так как сдвиг в этом биологического равновесия обычно играет роль в болезни и преобразования раковых клеток. Мембранные белки связаны одни из самых биологически важных класс белков, так как они могут инициировать комплекс сигнальных каскадов, а также посредником и импорта и экспорта различных молекул, в том числе наркотиков, которая была последние значения в области здравоохранения, как проблемы, связанные с лекарственной устойчивости становятся все более распространенным явлением. Получение понимание сложности этого белка класс требует идентификации их взаимодействия партнеров. Обнаружение таких партнеров оказался непростым, как это часто требует суровых условиях, которые должны быть оптимизированы для каждого мембраной белков [1].
Фундаментальные биологические и клинические важность интегральные мембранные белки стимулировали развитие дрожжей-системы для высокой пропускной способности идентификации ИЦП полнометражный белками трансмембранного. С этой целью мы разработали сплит-убиквитин основан дрожжей Мембрана два-Hybrid (МИФ) системы [2-4]. Этот инструмент позволяет чувствительны обнаружения переходных и стабильного взаимодействия белка. Она была успешно применена для изучения экзогенных и эндогенных белков выражается в модели CEREVISIAE организме Saccharomyces [3-7]. Миф использует наблюдение, что убиквитин могут быть разделены на две фрагменты. С-концевой половине (C иь) и N-терминальной половины (Нуби) в естественных условиях исследования показали, что эти фрагменты спонтанно воссоздать в связи с их высоким сродством к одна другую (Figure. 1а). Однако, представляя изолейцин 13 до глицин точечной мутации в N-концевую часть убиквитин (производство фрагмент называют N иЬ G) предотвращает это спонтанное повторное объединение [8] (Figure. 1б).
Мы используем этот принцип в МИФ системы (Figure. 1c и 2). Короче говоря, интегральный белок мембраны приманка сливается с фрагментом C иь, который связан с искусственным фактором транскрипции, состоящий из кишечной палочки ДНК-связывающий белок LexA и активации области VP16 вируса простого герпеса. Жертв создаются путем слияния кДНК или геномных ДНК производных фрагментов NubG части. Взаимодействие между приманкой и добычу белков в дрожжах хозяин приводит к воссоздание "псевдо-убиквитин 'полнометражный молекулы, с последующим признанием цитозольного deubiquitinating ферментов (дублирует) и протеолитических выпуск транскрипционный фактор. Транскрипционный фактор может ввести ядро клетки, и активировать системы ген-репортер (как правило, с участием выражение HIS3, LacZ и ADE2 генов), которые позволяют рост штаммов дрожжей на селективных средах, что свидетельствует о приманку и добычу взаимодействия [ 2-4].
Идентификация и характеристика интегрального взаимодействия белков мембраны предоставит информацию, которая поможет полностью понять их функции. Как мы более точно понимать и анализировать роль белков, которые взаимодействуют с интегральные мембранные белки, мы можем получить представление о динамическом взаимодействии участвует в регуляции этих белков и открыть для новых целей, которые могут иметь терапевтический потенциал.
2. Выбор приманки и соответствующие системы МИФ
- До проведения анализа МИФ, убедитесь, что белок имеет свой N-и / или С-конца в цитозоле клетки. Очень важно, чтобы C UB-LexA-VP16 теги быть приварены к вашим белка на такой вокзал, так как deubiquitinating ферменты, необходимые для транскрипции релиз фактор находятся в цитозоле [4].
- Затем решите, какой из двух основных вариантов МИФ подходит. Для неместных белками дрожжей, традиционные МИФ (tMYTH) могут быть использованы, в котором приманки гиперэкспрессия эктопически из плазмиды. Для нативных белков дрожжей, интегрированные МИФ (iMYTH) является методом выбора. В iMYTH приманки эндогенно с меткой C UB-LexA-VP16 теги, оставляя их под контролем родных промоутера. Это выгодно, как дикий тип уровня экспрессии приманки помогает устранить проблемы, связанные с гиперэкспрессией белка, такие как увеличение количества ложных срабатываний [3]. За исключением первоначального строительства приманку и селективных средах используются, обе формы МИФ осуществляются в основном одинаково. Для ясности, мы сосредоточимся в основном на использование tMYTH в этом докладе, так как этот вариант, в принципе, использоваться с мембраной белков практически из любого организма иТаким образом более широкое применение.
3. Поколение Bait и проверка
- Обязательные СМИ и пути их решения
- Стерильные DDH 2 O подготовлены в автоклаве при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- 3-амино-1 ,2,4-триазола (3-AT) раствор, приготовленный как 1М раствора акций DDH 2 O. Стерилизовать прохождения через фильтр 0,2 мкм.
- YPAD Рост СМИ, состоящую из 1% вес / экстракт дрожжей, 2% вес / пептон, 2% м / о глюкозы и 100 мкМ аденин подготовлен в DDH 2 O. Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- 10х аминокислот / нуклеотидные Mix Base: полная смесь содержит 1,0 ммоль аденина, урацила 1,8 мм, 1,0 мм аргинин, 1,0 мМ Гистидин, Изолейцин 2,3 мм, 7,6 мм лейцин, лизин 1,6 мм, 10,1 мм метионин, фенилаланин 3,0 мМ, 16,8 мМ треонин, триптофан 2,0 мМ, 1,7 мМ тирозин и 12,8 Валин мМ, подготовленный в DDH 2 O. Для отсева опустить необходимые аминокислоты (ы) и / или нуклеотидные основания (с). Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- Синтетические Drop-Out (SD) Рост Медиа, состоящий из 0,67% вес / объем дрожжей азотистого основания (без аминокислот, но с сульфатом аммония), 2% м / о глюкозы, 2% вес / агар и 1x Аминокислоты / нуклеотидные Mix, подготовлен в DDH 2 O. Подготовка жидких и твердых (содержащих 2% агара) SD-лейцин СМИ. Также приготовьте твердые SD-триптофан-лейцин и SD-триптофан-лейцин-аденин-Гистидин СМИ. Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут. Налейте в твердых средах 100x15 мм чашки Петри.
- Синтетические Drop-Out (SD) Рост средах, содержащих 3-AT. Подготовка SD-триптофан-лейцин-аденин-Гистидин средств массовой информации, как описано, но содержащий 3-АТ в концентрации 25, 50, 75 и 100 мм. Добавить необходимое количество 3-АТ от 1М стерильный раствор акции в средствах массовой информации после того, как автоклавного и охлажденной (но еще не затвердевшего). Налейте в 100x15 мм чашки Петри.
- PEG / ацетат лития смесь, состоящую из 40% вес / ПЭГ-3350, 120 мМ ацетат лития и 167 мкг / мл ДНК спермы лосося (тип III натриевая соль), подготовленный в DDH 2 O. Для обеспечения стерильности этой смеси приготовить из стерильной воды и растворов (например, автоклавного 50% ПЭГ-3350, автоклавного 1M ацетат лития и 2 мг / мл ДНК спермы лосося типа III натриевая соль, подготовленный в стерильных DDH 2 O).
- Ферментов и реагентов для проведения ПЦР.
- Коммерческая miniprep комплект.
- Сода извести стеклянные бусы (0,5 мм).
- Компетентные клетки кишечной палочки подходят для распространения плазмид (например DH5α, XL10 золото) и стандартных сред, пригодных для распространения и бактериальных плазмид выбора.
- Конкретные штаммы дрожжей, плазмиды и грунтовки, как описано в протоколе.
- Генерация tMYTH Приманки от Gap Ремонт
- Приманки должны быть клонирована в соответствующий вектор для маркировки и выражения. Разнообразие tMYTH векторов в настоящее время доступны для использования в строительстве приманку. Векторы, таких как pCMBV4, pAMBV4 и pTMBV4 позволяют строительство С-неизлечимо меткой приманки (приманка-C UB-LexA-VP16) под контролем CYC1 (слабый), Adh1 (сильный) и TEF1 (очень сильная) промоутеров, соответственно . N-неизлечимо меткой приманки (LexA-VP16-C UB-приманка) может быть создан с помощью векторов, таких как pTLB-1 и pBT3-N, под контролем TEF1 и CYC1 промоутеров, соответственно. Вектор выбор зависит от приманки и должен быть определен эмпирически. В некоторых случаях выше приманки выражение необходимо для того, чтобы обнаружить взаимодействий, в то время как в других случаях гиперэкспрессия приманки фактически может иметь пагубные последствия, что приводит к увеличению числа ложных срабатываний.
- Ограничение переварить выбранной плазмиды на соответствующий сайт ограничение (ы). Расщепление должно происходить только в непосредственной близости от C UB-LexA-VP16 тегом (вверх по течению от тегов для С-концевой пометки или вниз по течению для N-терминал пометки). Например, при использовании pAMBV4 вектор, SFII является идеальным выбором. Магазин переваривается плазмиды при температуре -20 ° С до готовности к использованию.
- Дизайн праймеров для амплификации и клонирование гена. 5 'конце вашего прямого праймера должна соответствовать примерно 35-40 нуклеотидов перед ограничением сайта, в то время как 3' конец должен соответствовать первому 18-20 нуклеотидов гена-мишени. 5 'конце обратного Грунтовка должна соответствовать обратном дополнение около 35-40 нуклеотидов ниже по течению от ограничения сайта, с 3' конца соответствующий обратный дополнение последние 18-20 нуклеотидов гена-мишени (опуская стоп-кодон если C UB-LexA-VP16 теги делается на С-конец). В зависимости от того, N-или С-концевой тегов выполняется, выберите 35-40 нуклеотидов переднего и заднего грунтовки, что ген-мишень клонируют в рамке с C UB-LexA-VP16 тега.
- Amplify интересующего гена методом ПЦР с использованием праймеров выше. ПЦР параметров будет зависеть от конкретного фермента и специфических праймеров.
- Преобразование ПЦР-продукта наряду с переваривается плазмиды в соответствующие лаборатории штамма дрожжей подшипников leu2 мутации (например, BY4741). Напряжение МИФ репортер (например THY.AP4 или L40) можно использовать, но не является необходимым, так как цель дрожжей в этот момент просто в качестве среды, в которой разрыв ремонт гомологичной рекомбинации может произойти. Преобразования должны проводиться следующим образом:
- Привить единственная колония выбранный штамм дрожжей в 5 мл стерильного СМИ YPAD и инкубировать в течение ночи при 30 ° С при постоянном встряхивании (200 оборотов в минуту).
- Развести ночной культуры в 50 мл свежей среды YPAD к OD600 от ~ 0,15 и инкубировать при температуре 30 ° C при встряхивании (200 оборотов в минуту). Вырасти в течение примерно 3-4 часов, пока OD600 ~ 0,6 будет достигнута.
- Центрифуга клеток 700xg в течение 5 минут и удалите супернатант.
- Ресуспендируют осадок клеток в 25 мл стерильного DDH 2 O и центрифуге при 700xg течение 5 минут.
- Удалить супернатант и ресуспендируют осадок клеток в 1 мл стерильного DDH 2 O.
- Добавить 100 мкл клеток, 300 мкл PEG / ацетат лития смесь, и переваривается плазмиды (50 фмоль) и ПЦР-продукта (250-500 фмоль), чтобы микроцентрифужную трубки.
- Инкубировать при температуре 30 ° С в течение 30 минут.
- Теплового шока при 42 ° С в течение 1 часа.
- Центрифуга на 3000xg в течение 5 минут и удалить супернатант.
- Ресуспендируют осадок клеток в 200 мкл стерильной DDH 2 O и пластины весь объем на твердую, SD-лейцин селективных средах. Рост при 30 ° С в течение 2-4 дней.
- Расти одной колонии преобразуются напряжение в 5 мл SD-лейцин жидкой среде при температуре 30 ° С в течение ночи.
- Центрифуга клеток 700xg в течение 5 минут и удалите супернатант.
- Изолировать приманки плазмидной ДНК из клеток гранулы с помощью любого коммерческого набора miniprep. Следуйте стандартным протоколом, с одной модификации. Для того, чтобы обеспечить достаточную лизиса клеток дрожжей, добавить небольшой объем 0,5 мм соды бисером известь стекла, чтобы шарик после первоначального ресуспендирования и вихревые энергично в течение 5 минут. Затем приступить к коммерческой протокол, как обычно.
- Преобразование выделенной ДНК дрожжей в компетентный E. палочки штамма подходит для плазмиды распространения (например, DH5α, XL10 Gold) с преобразованием КПД не менее 1х10 7 клеток / мкг ДНК. Обратите внимание, что приманка плазмиды могут быть выбраны для использования канамицина.
- Урожай плазмидной ДНК из трансформированных E. кишечной использовании стандартного метода выделения ДНК или коммерческого набора.
- Проверьте правильность строительство приманки плазмиды с помощью секвенирования.
- Преобразование проверено приманки построить в соответствующий штамм репортер МИФ (например THY.AP4, L40). Протокол дрожжей преобразования только что описал может быть использовано, подставляя приманки плазмидной ДНК на месте переваривается плазмиды и ПЦР-продукта.
- Приманка Validation - Правильная локализация
- Перед использованием приманки штаммов анализируются для того, чтобы они правильно локализован дрожжей мембраны. При использовании iMYTH, эта локализация будет зависеть в частности от свойств меткой приманкой. Для tMYTH, наживка плазмиды как правило, включают последовательность сигналов (например, Matα) направление выразил белка в плазме мембраны. Локализация определяется с помощью флуоресцентной микроскопии. Включение YFP молекулы в последовательность тэгов приманку (например, C UB-YFP-LexA-VP16) является наиболее простой и прямой подход, что позволяет прямой визуализации живых клеток, и обычно используется в iMYTH. Кроме того, стандартные иммунофлюоресценции подход с использованием антител против LexA или VP16 компоненты тег может быть использован.
- Приманка Validation - N иЬ G / I контрольный тест
- После надлежащей локализации приманка была создана, необходимо обеспечить, чтобы приманка не активирует репортер системы в одиночку или в присутствии невзаимодействующих жертвы (то есть убедиться, что приманка не является само собой активации). Это достигается с помощью N иЬ G / I Test, где приманка трансформируется с взаимодействующими (положительный) и невзаимодействующих (отрицательный) жертвы контролем, и рост оценивается на селективных средах. Приманка должна расти на селективных средах яN наличие положительного контроля и не растут в присутствии отрицательный контроль, для того, чтобы быть пригодными для использования в МИФ.
- Можно приступить к преобразованию приманки напрягаться, чтобы быть проверены с 100-200 нг контроль добычу плазмиды. Описанных ранее протокол дрожжей преобразование может быть использовано, подставляя SD-лейцин средств массовой информации в месте YPAD и использование SD-триптофан-лейцин средств массовой информации для последнего шага металлизации. Следующие конструкции добычу управления обычно используются:
- pOST1-N иь I (положительный контроль)
- pOST1-N иЬ G (отрицательный контроль)
- pFUR4-N иь I (положительный контроль)
- pFUR4-N иЬ G (отрицательный контроль)
- OST1 является компонентом oligosaccharyl сложные и локализуется в эндоплазматический ретикулум мембраны [9], а FUR4 является урацил permease и локализуется в мембране [10]. Хотя эти белки, как правило, нашим первым выбором для использования в качестве невзаимодействующих жертвы, их пригодности будет меняться от случая к случаю. В редких случаях, когда приманка, по прогнозам, действительно взаимодействуют с обоими из этих элементов управления, альтернативные жертвы должны быть выбраны. Напомним, что N иь я это дикого типа форма N-конце убиквитин, и спонтанно взаимодействует с C иь зависят от взаимодействия между белками, которые C иь и N иЬ слиты. Таким образом, N иь я жертв составляют положительного контроля, а N иЬ G жертвы (проведение Изолейцин 13 до Глицин мутация, которая препятствует спонтанной ассоциации N иь и иь C) выступают в качестве отрицательного контроля.
- Ресуспендируют одной колонии каждого преобразован приманку в 100 мкл стерильной DDH 2 O.
- Серийно разбавить ресуспендировали клеток в стерильных DDH 2 O для получения разведения 1 / 10, 1 / 100 и 1 / 1000.
- Spot 5 мкл неразведенного и разбавленного клеток на SD-триптофан-лейцин и SD-триптофан-лейцин-аденин-Гистидин средств массовой информации с учетом и без 3-АТ в диапазоне концентраций. 3-AT действует как конкурентный ингибитор HIS3 ген-репортер, и служит для увеличения жесткости отбора. Это может быть полезно в некоторых случаях для подавления неспецифической рост слабо или умеренно самоактивирующимися приманок.
- Разрешить пятна высохнуть, а затем инкубировать пластин при 30 ° С в течение 2-4 дней.
- Все трансформанты должны расти на SD-триптофан-лейцин пластин, указывая, что они были успешно трансформируется с добычей плазмиды. Приманки, которые не самостоятельно активировать вырастет на SD-триптофан-лейцин-аденин-Гистидин средства массовой информации только тогда, когда трансформированные N иь я добычей конструкции, а не с N иЬ G жертв. Обратите внимание, что концентрация 3-AT требуется в средствах массовой информации (если таковые имеются), поскольку это необходимо для использования во время проверки.
4. Скрининг
- Обязательные СМИ и пути их решения
- Стерильные DDH 2 O подготовлены в автоклаве при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- 0,9% NaCl Раствор готовят в DDH 2 O и стерилизовать в автоклаве при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- Фосфат натрия раствор, состоящий из 493 мМ фосфат натрия двузамещенный и 250 ммоль натрия фосфат однозамещенный в DDH 2 O. Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- X-Gal (5-бром-4-хлор-3-индолил-β-D-галактопиранозид) Раствор готовят, как 100 мг / мл маточного раствора в N, N-диметилформамид.
- 2xYPAD Рост СМИ, содержащий 2% вес / дрожжевой экстракт, 4% вес / пептона, 4% м / о глюкозы и 100 мкМ аденин, подготовленный в DDH 2 O. Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- Синтетические падением напряжения (SD) Рост Медиа, приготовленного, как описано ранее. Подготовка жидких SD-лейцин и твердых SD-триптофан-лейцин. Налейте твердых сред в оба 100x15 мм чашки Петри. Подготовка твердых SD-триптофан-лейцин-аденин-Гистидин СМИ в 150 мм круглых пластин, 16 пластин для каждого экрана, содержащий 3-АТ, при необходимости, при концентрации определяется из N иЬ G / I контрольный тест.
- Синтетические падением напряжения (SD) Средства массовой информации + 5-бром-4-хлор-3-Indoyl-β-D-галактопиранозид (X-Gal). Подготовка SD-триптофан-лейцин-аденин-Гистидин сред, содержащих агар, как описано выше. После автоклавирования, дать остыть, добавить 3-AT (при необходимости), а затем 1/10th объемом ое стерильный раствор фосфата натрия. Затем добавьте X-Gal Решение конечной концентрации 80 мкг / мл. Тщательно перемешать и разлить в 150 мм круглых пластин.
- PEG / ацетат лития Solution II, содержащая 40% ПЭГ-3350, 100 мМ ацетат лития, 1 мМ EDTA и 10 мМ Трис рН 7,5. Подготовка этого решения с использованием стерильных DDH 2 O и решений (например, автоклавного 50% ПЭГ-3350, 1 М ацетата лития, 100 мМ Трис рН 7,5 и 500 мМ ЭДТА рН 8,0).
- Литий ацетат / Tris ЭДТА раствором, содержащим 110 мМ ацетат лития, 11 мМ Трис рН 7,5 и 1,1 мМ ЭДТА. Подготовка этого решения с использованием стерильных DDH 2 O и решений (например, автоклавного 1M ацетат лития, 100 мМ Трис рН 7,5 и 500 мМ ЭДТА рН 8,0).
- 10x Трис ЭДТА раствор, состоящий из 100 мМ Трис рН 7,5 и 10 мМ ЭДТА, подготовленный в DDH 2 O. Стерилизовать автоклавированием при температуре 121 ° С, 15 фунтов на квадратный дюйм в течение 30 минут.
- Одноцепочечных ДНК перевозчика (оцДНК) Раствор, содержащий 2 мг / мл ДНК спермы лосося Тип III натриевая соль, подготовленный в стерильных DDH 2 O.
- Коммерческая miniprep комплект.
- Сода извести стеклянные бусы (0,5 мм).
- Компетентные клетки кишечной палочки подходят для распространения плазмид (например DH5α, XL10 золото) и стандартных сред, пригодных для распространения и бактериальных плазмид выбора.
- Конкретные дрожжевые штаммы и плазмиды, как описано в протоколе
- Большой масштабного преобразования
- Привить единственная колония штамма МИФ репортер содержащих вашу приманку в 5 мл SD-лейцин СМИ и инкубировать в течение ночи при 30 ° C при встряхивании (200 оборотов в минуту).
- Развести ночной культуры в 200 мл SD-лейцин СМИ OD600 = 0,15 и инкубировать при температуре 30 ° C при встряхивании (200 оборотов в минуту). Вырасти до OD600 = 0,6 - 0,7 (примерно 4-5 часов).
- Незадолго до целевых OD600 достигнута, оттепель аликвоту решение оцДНК. Варить при 100 ° С в течение 5 минут и затем охладить на льду. Повторите еще раз.
- Когда цель OD600 не было достигнуто, урожай клеток с помощью центрифугирования при 700xg в течение 5 минут (поделите 200 мл культуры между 4x50 мл завинчивающейся крышкой центрифужные пробирки).
- Вымойте каждый шарик с 30 мл стерильного DDH 2 O и кратко вихрь образца. Центрифуга на 700xg течение 5 минут.
- Удалите супернатант и ресуспендируют каждую крупинку в 1 мл ацетатного литий / Tris раствором ЭДТА. Трансфер в стерильные 1,5 мл трубки микроцентрифужную и центрифуге при 700xg течение 5 минут.
- Удалите супернатант и ресуспендируют каждую крупинку в 600 мкл ацетата литий / Tris раствором ЭДТА.
- Добавьте следующие 4x15 мл завинчивающейся крышкой трубы центрифуги:
- 2,5 мл ПЭГ / ацетат лития Solution II
- 600 мкл ресуспендировали клетки
- 100 мкл решение оцДНК
- 7 мкг хищных библиотеки ДНК
- Библиотеки содержащие жертв меткой на любом N-или С-конца с N иЬ G, и получают из различных кДНК или геномных источников, имеющихся в продаже ( www.dualsystems.com ). Конкретной библиотеки, используемой должны определяться в каждом конкретном случае на индивидуальной основе, в зависимости от выбранной приманки и экспериментальных задач.
- Вихревые трубы в течение 1 минуты, чтобы обеспечить тщательное перемешивание, а затем инкубируют в 30 ° С водяной бане в течение 45 минут. Смешайте кратко каждые 15 минут.
- Добавить 160 мкл диметилсульфоксида (ДМСО) в каждую пробирку и смешать сразу обращением труб.
- Инкубируйте в 42 ° С водяной бане в течение 20 минут.
- Сбор трансформантов центрифугированием при 700xg течение 5 минут.
- Удалите супернатант. Восстановление трансформантов по ресуспендирования каждой гранулы в 3 2xYPAD мл. Бассейн все образцы вместе в одном 50 мл завинчивающейся крышкой центрифуги трубы.
- Инкубировать при 30 ° С в течение 90 минут для восстановления клеток.
- Центрифуга на 700xg в течение 5 минут и отбросить супернатант.
- Ресуспендируют ячейки гранул в 4,9 мл стерильного 0,9% NaCl.
- Использование 100 мкл ресуспендировали клетки подготовить 10-кратный серийных разведений в стерильной 0,9% NaCl от 10x до 1000x.
- Пластины по 100 мкл 100x и 1000x разведения на SD-триптофан-лейцин СМИ и инкубировать при температуре 30 ° С в течение 2-3 дней. Эти плиты служат контролем и используются для расчета эффективности трансформации.
- Столь же разделить оставшиеся 4,8 мл ресуспендировали клетки и пластиной на большие (150 мм) SD-триптофан-лейцин-аденин-Гистидин плит,содержащие необходимое количество 3-AT, который определен в N иЬ G / I тестирования и инкубировать при температуре 30 ° С в течение 3-4 дней.
- Ресуспендируют одной колонии (каждый из которых представляет клеток, содержащих потенциальные взаимодействия приманки добычи пара) в 100 мкл 0,9% NaCl и пластины 5 мкл аликвот на SD-триптофан-лейцин-аденин-гистидин + X-Gal СМИ (и в том числе 3-АТ, если требуется). Разрешить расти в течение 2-4 дней. Этот шаг служит второй раунд селективного скрининга, а также помогает в снятии ложных срабатываний, полученные в первый раунд. Только колоний, которые показывают высокие темпы роста и синего цвета выбраны для дальнейшего анализа.
- Prey Изоляция ДНК и секвенирования
- Изолировать плазмидной ДНК из синих колоний дрожжей, используя протокол miniprep с изменениями, описанных выше. Будьте уверены, чтобы вырастить клетки в SD-триптофан СМИ только, чтобы выбрать для удержания добычи, но не приманка, плазмид. Для экранов, которые производят большое количество хитов, имеющихся в продаже высокой пропускной miniprep комплект может быть выгодно в этой точке.
- Преобразование изолированных дрожжей плазмида ДНК в компетентные E. палочки штамма подходит для плазмиды распространения (например, DH5α, XL10 Gold) с преобразованием КПД не менее 1х10 7 клеток / мкг ДНК. Обратите внимание, что жертвой плазмиды могут быть выбраны для использования ампициллин.
- Изолировать плазмидной ДНК из трансформированных E. кишечной использовании стандартного метода выделения ДНК или коммерческого набора. Еще раз, высокой пропускной miniprep Набор может быть полезно, если номер пробы велика. Усиление ДНК в E. кишечной значительно повышает урожайность и плазмиды гарантирует, что достаточное количество ДНК присутствует как для секвенирования и дальнейшего анализа.
- Последовательность изолированных плазмид использованием грунтовки дополнением к последовательности в N иЬ Г.
- Обобщение и анализ всех данных секвенирования собрать ваш предварительный список interactors. Это может быть сделано вручную или в автоматическом режиме с помощью соответствующих программ.
- Тестирование Bait зависимость
- После сборки предварительный список interactors важно перепроверить взаимодействий и устранить беспорядочный жертвы, которые взаимодействуют / активировать репортер системы таким образом, независимо от приманки идентичности. Это достигается с помощью испытаний Bait зависимостей. В этом тесте все выявленные interactors превращаются обратно в исходный штамм приманки, а также напряжение укрывательство контроль искусственные приманки, состоящие из одного домена трансмембранного слит с C UB-LexA-VP16 тега. Преобразование производится в соответствии со стандартным протоколом описано выше, с помощью SD-лейцин средств массовой информации в месте YPAD и SD-триптофан-лейцин твердых сред для последнего шага металлизации.
- Ресуспендируют одной из колоний выше преобразований в 100 мкл стерильной DDH 2 O и SPOT 5 мкл на SD-триптофан-лейцин-аденин-гистидин + X-Gal СМИ (и в том числе 3-АТ при необходимости). Плиты затем выдерживают в течение 2-4 дней при температуре 30 ° C. В идеале нескольких трансформантов должны быть выбраны для каждого добычу, и оба оригинальные приманки и искусственные приманки должны быть выставлены на ту же пластинку.
- Дрожжи проведения искусственной приманки и добычей, которые вызывают активацию репортера системе (то есть рост и синего цвета) считаются беспорядочные и что конкретные добычу удаляется из списка interactors.
- Жертвы, которые вызывают рост и синее окрашивание в дрожжах с приманки из интереса, а не искусственные приманки, подтверждает это специфическое взаимодействие. Если, однако, дрожжи укрывательство добычу и вашу приманку интересов, не растут, эта жертва будет удален из списка interactors.
- Остальные жертвы составляют полный список выявленных в interactors МИФ экране.
5. Дальнейшие исследования
Как только МИФ скрининга была завершена, дальнейших анализов должны быть выполнены для того, чтобы проверить и определить биологическое значение обнаружено взаимодействий. Конкретных исследований, выполняемых будет меняться от случая к случаю, и должны быть определены отдельным исследователем. Некоторые общие примеры последующей работы включают в себя совместное иммунопреципитации экспериментов и удаление исследования в родной организм. Кроме того, численный анализ полученных данных могут быть полезны для выявления моделей, и помогая определить потенциальное значение и роль различных взаимодействий может играть. Таким образом, миф технология служит мощным первым шагом »в направлении выявления и понимания критических функциональных взаимодействий мембранных белков. В сочетании с подробным последующих исследований и других недавно развитых и развивающихся тэchnologies, он обещает быть ценным инструментом в раскрытии тайн клетки.

Рисунок 1. Принцип сплит убиквитин. A. Убиквитин могут быть разделены на две фрагменты: С-концевой половине (C иь) и N-концевой половине (N иь I). Эти фрагменты спонтанно восстановить из-за их высокое сродство к одной другую. b. Точки иь я мутация в позиции 13 из изолейцин, чтобы глицина (N иЬ G) предотвращает это спонтанное повторного объединения. c. В МИФ системы, C иь сливается с приманкой интересов, (B) и добычу слит с N иЬ G (A). А. Б. взаимодействия белка воссоздает псевдо-убиквитин.
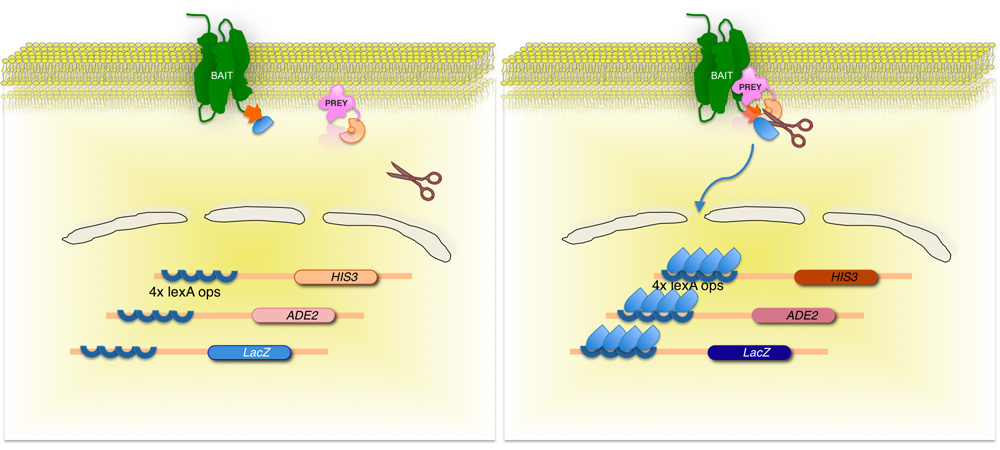
Рисунок 2. Сплит-убиквитин основан дрожжей мембраны двух гибридных (МИФ) системы.
Мембранный белок интересов, (приманка) сливается с С-концевой половине дрожжей убиквитин (С Ub), конъюгированные с фактором транскрипции. Использование кДНК или gDNA библиотеки, каждый белок, кодируемый библиотеки (добычу) сливается с соответствующим N-конце убиквитин фрагмент (N Ub G). Если два белка не взаимодействуют, транскрипционный фактор остается на мембране интерфейс (слева). Однако, если белки взаимодействуют, два убиквитин фрагменты присоединиться, в результате расщепления убиквитин специфических протеаз. Расщепление релизы транскрипционный фактор, в результате чего выражение репортер генов (справа)

Рисунок 3. МИФ трубопровода.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Миф первый высокой пропускной способности системы, что позволяет идентифицировать взаимодействие между полнометражный мембранных белков и цитозольного или мембраной партнеров. Он был использован для изучения мембранных белков из различных организмов [3-7]. Есть, однако, конкретные детали, которые, возможно, должны быть тщательно проанализированы для обеспечения белком интересов, потенциально пригодных для исследования с МИФ.
Многие мембраной белков направлены на мембране с помощью сигнальной последовательности, которая впоследствии расщепляется, чтобы произвести зрелый белок. Эта последовательность организма конкретного и вполне возможно, что это родной сигнальной последовательности приведет к неправильной локализации белка интересов, при выраженных в дрожжах, так как последовательность сигналов, остается незамеченной. Чтобы обойти эту проблему, мы инженерии этих специфических белков быть объединены в последовательность дрожжей сигнал, полученный из альфа Спаривание фактор (MATα). Этот пептид последовательности (так называемые MFα-сс) повторно локализует белков плазматической мембраны дрожжей и главное, расщепляется пептидаз дрожжей сигнала. Этот пептид последовательности находится в плазмиды pTMBV-MFα и pAMBV-MFα.
Другим важным параметром, который необходимо подчеркнуть, заключается в уровне приманки выражения. Промоутер, который управляет выражением приманки регулирует этот параметр. Это может потребоваться для оптимизации приманки уровни экспрессии использованием NubG / Нуби тестирования и количестве 3-АТ, необходимых для устранения артефактов гиперэкспрессия, где приманка "самоактивирующимися" (т.е. без разбора взаимодействует со многими неспецифические белки добычи) . Экспертиза дрожжей белки производит наиболее физиологически соответствующие приманки концентрациях, когда iMYTH применяется. В этом случае ген интересов, помечается Cub-TF в gDNA. Кроме того, экзогенные белки могут быть выражены с pBT3-STE и pCMBV плазмиды, несущие CYC1 промоутер в результате низкой экспрессией приманки. Плазмид pTMBV и pTLB1 гавани TEF1 промоутер в то время как pAMBV имеет Adh1 промоутер, как то, что диск сильным выражением приманки белка. Если уровень белка приманки требуют дальнейшей оптимизации, это может быть необходимо использовать pTLB-1 плазмиды, которая несет TEF1 промоутера, однако, ДНК-связывающий домен LexA мутирует в R156G стихать склонность к экзогенным промоутеров гена-репортера, в конечном счете, снижению Вероятность самодеятельности [5].
Другим фактором, который играет важную роль для МИФ успех библиотеки выбора используется для скрининга. Это будет зависеть от эндогенных профилей выражения приманки. Например, приманка может быть выражена в конкретных тканей, и поэтому важно использовать библиотеку, которая строится из данной ткани. Это позволит обеспечить физиологически соответствующих взаимодействий обнаружено.
МИФ системы простой и быстрый инструмент, который предоставляет большое количество информации о класс белков, которые были трудны для изучения. Эти определены взаимодействия может помочь в выяснении полной биологической функции мембранных белков.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Игорь Stagljar является соучредителем DualSystems Biotech, Швейцария.
Acknowledgments
Мы хотели бы поблагодарить Рассвет Эдмондс для критического прочтения этой статьи. Лаборатория Stagljar поддерживается за счет средств Канадского фонда для инноваций (CFI), Канадский институт исследований в области здравоохранения (CIHR), сердца и инсульта фонда Канадского общества рака, и Novartis.
Materials
| Name | Company | Catalog Number | Comments |
| Polyethenlene Glycol (PEG3350) | BioShop Canada | PEG335 | |
| Lithium Acetate Bihydrate | BioShop Canada | LIA001 | |
| X-Gal (5-Bromo-4-Chloro-3-Indolyl-b-D-galactopyranoside) | BioShop Canada | XGA001 | |
| N`,N-dimethyl formamide | BioShop Canada | DMF 451 | |
| 3-amino-1,2,4-triazole (3-AT) | BioShop Canada | ATT124 | |
| Sodium phosphate dibasic | BioShop Canada | SPD307 | |
| Sodium phosphate monobasic | Fisher Scientific | BP329-500 | |
| Salmon Sperm DNA | VWR international | CA80601-120 | |
| D-Glucose | BioShop Canada | GLU501 | |
| LB Broth LENOX | BioShop Canada | LBL405 | |
| Yeast Nitrogen Base | BioShop Canada | YNB406 | |
| Yeast Extract | BioShop Canada | YEX401 | |
| Peptone | BD Biosciences | 211677 | |
| Bio-Tryptone | BioShop Canada | TRP402 | |
| Adenine Sulphate | BioShop Canada | ADS201 | |
| L-Uracil | BioShop Canada | URA241 | |
| L-Threonine | BioShop Canada | THR002 | |
| L-Histidine | BioShop Canada | HIS200 | |
| L-Methionine | BioShop Canada | MET222 | |
| L-Valine | BioShop Canada | VAL201 | |
| L-Phenylalanine | BioShop Canada | PHA302 | |
| L-Isoleucine | BioShop Canada | ISO910 | |
| L-Tyrosine | BioShop Canada | TYR333 | |
| L-Leucine | BioShop Canada | LEU222 | |
| L-Arginine | BioShop Canada | ARG006 | |
| L-Tryptophane | Fisher Scientific | BP395-100 | |
| L-Lysine | BioShop Canada | LYS101 | |
| L-Alanine | Fisher Scientific | BP369-100 | |
| Agar | BioShop Canada | AGR001 | |
| Soda Lime Galss Beads | Biospec Products | 11079105 | |
| Sodium Chloride | BioShop Canada | SLD002 |
References
- Stagljar, I., Fields, S. Analysis of membrane protein interactions using yeast-based technologies. Trends Biochem Sci. 27 (11), 559-563 (2002).
- Iyer, K. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci STKE. 275, pl3-pl3 (2005).
- Paumi, C. M. Mapping protein-protein interactions for the yeast ABC transporter Ycf1p by integrated split-ubiquitin membrane yeast two-hybrid analysis. Mol Cell. 26 (1), 15-25 (2007).
- Stagljar, I. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc Natl Acad Sci U S A. 95 (9), 5187-5192 (1998).
- Gisler, S. M. Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol Cell Proteomics. 7 (7), 1362-1377 (2008).
- Scheper, W. Coordination of N-glycosylation and protein translocation across the endoplasmic reticulum membrane by Sss1 protein. J Biol Chem. 278 (39), 37998-38003 (2003).
- Thaminy, S. Identification of novel ErbB3-interacting factors using the split-ubiquitin membrane yeast two-hybrid system. Genome Res. 13 (7), 1744-1753 (2003).
- Johnsson, N., Varshavsky, A. Split ubiquitin as a sensor of protein interactions in vivo. Proc Natl Acad Sci U S A. 91 (22), 10340-10344 (1994).
- Kelleher, D. J., Gilmore, R. The Saccharomyces cerevisiae oligosaccharyltransferase is a protein complex composed of Wbp1p, Swp1p, and four additional polypeptides. J Biol Chem. 269 (17), 12908-12917 (1994).
- Chevallier, M. R. Cloning and transcriptional control of a eucaryotic permease gene. Mol Cell Biol. 2 (8), 977-984 (1982).
