

Abstract
أصبح من الواضح بشكل متزايد أن التوزيع المكاني وحركة مكونات غشاء مثل الدهون والبروتينات هي عوامل أساسية في تنظيم العديد من الوظائف الخلوية. ولكن نظرا لديناميكية سريعة وهياكل صغيرة المعنية، مطلوب قرار المكانية والزمانية عالية جدا للقبض على السلوك الحقيقي من الجزيئات. هنا نقدم بروتوكول تجريبي لدراسة ديناميات البروتينات والدهون البلازما غشاء fluorescently المسمى في الخلايا الحية لقرار الزمانية المكانية عالية. والجدير بالذكر، لا يحتاج هذا النهج لتتبع كل جزيء، لكنه يحسب سلوك السكان الذين يستخدمون جميع الجزيئات في منطقة معينة من الغشاء. نقطة البداية هي التصوير السريع في منطقة ما على غشاء. بعد ذلك، يتم احتساب المكانية والزمانية وظيفة كاملة الارتباط الذاتي ربط الصور المكتسبة في زيادة تأخير الوقت، على سبيل المثال كل 2، 3، ن التكرار. فمن الممكن إثبات أن العرضمن ذروة المكانية زيادات ظيفة الارتباط الذاتي في زيادة تأخير الوقت بوصفها وظيفة من حركة الجسيمات بسبب نشر. لذا، من المناسب من سلسلة من الوظائف تمكن الارتباط الذاتي لاستخراج البروتين الفعلي يعني تشريد مربع من التصوير (iMSD)، قدم هنا في شكل واضح انتشارية مقابل متوسط النزوح. هذه تعطي الرأي الكمي من متوسط ديناميكية الجزيئات واحدة مع دقة نانومتر. باستخدام متغير GFP الموسومة من مستقبلات ترانسفيرين (TFR) وATTO488 المسمى 1-بالميتويل-2-هيدروكسي SN -glycero-3-phosphoethanolamine (PPE) فمن الممكن لمراقبة تنظيم الزماني المكاني للبروتين والدهون نشر على المناطق غشاء ميكرون الحجم في نطاق الوقت الجزئي لملي ثانية.
Introduction
بدءا من النموذج الأصلي "فسيفساء السائل" حسب المغني ونيكلسون، وصورة من الغشاء البلازمي الخلوي قد تم تحديثه بشكل مستمر خلال العقود الماضية من أجل إدراج دور الناشئة في الهيكل الخلوي والدهون المجالات 1،2.
تم الحصول على الملاحظات الأولى الانتعاش بعد photobleaching من الفلورسنت (FRAP) إزاحة الستار أن جزءا كبيرا من البروتينات الغشاء هو متحرك 3-5. هذه الدراسات الرائدة، على الرغم مفيدة للغاية، عانى من قرار فقيرة نسبيا في الفضاء (ميكرون) والزمن (ثانية) من الاجهزة FRAP. أيضا، ويجري قياس المتوسط الفرقة، FRAP يفتقر في إعطاء معلومات عن سلوك جزيء واحد.
في هذا السياق، وإمكانية لتسمية تحديدا جزيء واحد مع علامات مشرقة جدا (السماح دراسة عملية الانتشار جزيء في وقت واحد) كان ناجحا جدا. بشكل خاص، عن طريق دفعقرار وقت نهج احدة تتبع الجسيمات (SPT) إلى الجدول الزمني ميكروثانية، Kusumi، وآخرون. تمكنت من الوصول إلى ميزات لم تكن معروفة من الدهون والبروتين الديناميات التي أسهمت إلى حد كبير في التعرف على دور القائم على الأكتين هيكل عظمي في غشاء غشاء علم وظائف الأعضاء 6 7. ولدت هذه النتائج ما يسمى في "اعتصام والسور 'النموذج، الذي ينظم الدهون والبروتين نشر بواسطة هيكل عظمي القائم الأكتين. ومع ذلك، من أجل الحصول على كمية هائلة من المعلومات التي قدمتها SPT العديد من القضايا التجريبية لا بد من معالجتها. خاصة، وعادة ما تتألف الإجراء وضع العلامات خطوات كثيرة مثل إنتاج وتنقية وإدخال الأنواع المسمى في النظام. وعلاوة على ذلك، والعلامات الكبيرة، مثل نقاط الكم أو الجسيمات النانوية المعدنية، وغالبا ما يطلب للوصول إلى الجدول الزمني ميلي ثانية واحدة فرعية ويشابك من الجزيئات المستهدفة من قبل التسمية لا يمكن تجنبها في كثير من الحالات. أخيرا، العديد من مساراتوقد ليتم تسجيلها لتناسب المعايير الإحصائية وبصورة متزامنة مطلوب كثافة منخفضة التسمية للسماح تتبع.
مقارنة SPT، مضان الطيفي الارتباط (FCS)، والتغلب على كثير من هذه السلبيات، يمثل نهجا واعدا جدا لدراسة ديناميات الجزيئية. في الواقع، يعمل FCS جيدا أيضا مع تسميات قاتمة وكثيفة، مما يتيح لدراسة ديناميكية الجزيئات الموسومة بروتين فلوري في خلايا عابر. أيضا، فإنه يتيح الوصول إلى إحصاءات عالية في كمية محدودة من الزمن. وأخيرا، على الرغم من "عالية" كثافة العلامات يوفر معلومات FCS الجزيئات واحدة. بفضل كل هذه الخصائص، FCS يمثل نهجا واضحة جدا وطبقت على نطاق واسع لدراسة الدهون والبروتين ديناميكية في كل من الأغشية ونموذج في الخلايا الحية 8-10. لقد تم اقتراح العديد من الطرق المختلفة لزيادة قدرة FCS للكشف عن تفاصيل الانتشار الجزيئي. على سبيل المثال، كان SHالخاصة التي عن طريق أداء FCS على مختلف المناطق الملاحظة الحجم يمكن لأحد أن تعريف "القانون نشر FCS" ملامح المنير الخفية للحركة الجزيئات 11،12. إضافة إلى كونه تختلف في الحجم، وقد تتكرر مجال التركيز أيضا 13، وانتقل في الفضاء على طول خطوط 14-20 أو مترافق مع الكاميرات سريعة 21،22. باستخدام هذه العلاقة "المكانية والزمانية" النهج، وقد وصفت المعلمات البيولوجية ذات الصلة من عدة مكونات غشاء كميا على كل الأغشية البيولوجية نموذج ومنها الفعلية، وبالتالي البصيرة ذات العوائد إلى غشاء التنظيم المكاني.
ومع ذلك، في كل FRAP والتطبيقات FCS وصفها حتى الآن حجم المنطقة المحورية يمثل الحد في القرار المكانية التي لا يمكن التغلب عليها. وقد وضعت عدة طرق التصوير فائقة الدقة مؤخرا لتجاوز هذا الحد. وتقوم بعض على دقة الترجمة، مثل إعادة الإعمار العشوائية المجهر الضوئي (STORM) <سوب> 23،24، تنشيط ضوئي توطين المجهري (النخيل) 25، PALM مضان (FPALM) 26، واحد الجسيمات تتبع PALM (sptPALM) 27: على كمية كبيرة نسبيا من الفوتونات المطلوبة في كل لقطة، ومع ذلك، يحد القرار وقت هذه الأساليب لعدة ميلي ثانية على الأقل، مما يعيق إمكانية تطبيقها في الجسم الحي.
في المقابل، تم فتح بديلا واعدا للقرار التصوير الفاخر وتحوير مكانيا انبعاث مضان مع طرق استنزاف الانبعاث المستحث (STED أو عكسها تشبع التحولات مضان البصرية (RESOLFT)) 28،29. الجمع بين هذه النهج في تشكيل حجم الملاحظة أقل بكثير من الحد حيود مع إمكانية استخدام المجاهر المسح السريع وأنظمة الكشف. في تركيبة مع تحليل مضان التذبذب، STED المجهري يسمح للتحقيق مباشرة ديناميات الزمانية المكانية النانو من الدهون وصroteins في أغشية الخلايا الحية 30،31.
ويمكن الحصول على نفس الكميات الفيزيائية المجهر مقرها STED من قبل معدلة المكانية والزمانية صورة ارتباط التحليل الطيفي (عصي 32،33) الطريقة التي هي مناسبة لدراسة ديناميات بروتينات fluorescently الموسومة الغشاء و / أو الدهون في الخلايا الحية وبواسطة المجهر التجاري. ويتكون البروتوكول التجريبي المقدمة هنا خطوات قليلة. أول واحد يتطلب التصوير السريع من منطقة الفائدة على الغشاء. ثم، يتم استخدام كومة الناتجة من الصور لحساب متوسط ظائف علاقة المكانية والزمانية. عن طريق تركيب سلسلة من المهام ارتباط والجزيئي "القانون نشر" يمكن الحصول عليها مباشرة من التصوير في شكل واضح انتشارية (D التطبيق) - مقابل متوسط درجات مؤامرة التهجير. هذه المؤامرة يعتمد بشكل حاسم على البيئة استكشافها من قبل الجزيئات ويسمح تدرك مباشرة وسائط نشر الفعليةمن الدهون / البروتين من الفائدة.
في مزيد من التفاصيل، كما هو موضح سابقا 34 وظيفة لصناعة السيارات في العلاقة المكانية والزمانية من سلسلة صورة المكتسبة يعتمد بشكل حاسم على ديناميكية الجزيئات تتحرك في صورة سلسلة جمعها (يرجى ملاحظة أن نفس المنطق يمكن تطبيقه في اقتناء خط حيث يعتبر البعد واحد فقط في الفضاء). على وجه الخصوص، نحدد وظيفة العلاقة على النحو التالي:
 (1)
(1)
أين  يمثل قياس كثافة مضان في موقف X، Y و في الزمن t،
يمثل قياس كثافة مضان في موقف X، Y و في الزمن t، ![]() و
و ![]() يمثل المسافة في x والاتجاهات ذ التوالي،
يمثل المسافة في x والاتجاهات ذ التوالي، ![]() يمثل تأخر الوقت، و
يمثل تأخر الوقت، و ![]() يمثل المعدل. هذه الوظيفة يمكن التعبير عنها على النحو التالي:
يمثل المعدل. هذه الوظيفة يمكن التعبير عنها على النحو التالي:
 (2)
(2)
حيث يمثل 'N' متوسط عدد الجزيئات في مجال المراقبة، ![]() تمثل عملية الالتواء في الفضاء، و
تمثل عملية الالتواء في الفضاء، و  يمثل الارتباط الذاتي من الخصر فعال. هذا الأخير يمكن أن تفسر على أنها مقياس لمدى تنتشر الفوتونات من باعث واحد في الفضاء بسبب البصرية / تسجيل الإعداد (ما يسمى انتشار نقطة وظيفة، قوات الأمن الفلسطينية، الجينيحشد يقترب بشكل جيد من قبل وظيفة جاوس). أخيرا،
يمثل الارتباط الذاتي من الخصر فعال. هذا الأخير يمكن أن تفسر على أنها مقياس لمدى تنتشر الفوتونات من باعث واحد في الفضاء بسبب البصرية / تسجيل الإعداد (ما يسمى انتشار نقطة وظيفة، قوات الأمن الفلسطينية، الجينيحشد يقترب بشكل جيد من قبل وظيفة جاوس). أخيرا،  يمثل احتمال لإيجاد الجسيمات على مسافة
يمثل احتمال لإيجاد الجسيمات على مسافة ![]() و
و ![]() بعد فترة زمنية
بعد فترة زمنية ![]() . إذا اعتبرنا ديناميكية ناشر، والتي تتحرك الجزيئات بشكل عشوائي في جميع الاتجاهات وصافي التدفقات ليست موجودة، وهذه الوظيفة أيضا يقترب بشكل جيد من قبل وظيفة جاوس حيث يمكن تحديد التباين مثل متوسط ميدان الإزاحة (MSD) للجسيمات تتحرك . وهكذا، فإن الخصر وظيفة ارتباط (ويشار أيضا
. إذا اعتبرنا ديناميكية ناشر، والتي تتحرك الجزيئات بشكل عشوائي في جميع الاتجاهات وصافي التدفقات ليست موجودة، وهذه الوظيفة أيضا يقترب بشكل جيد من قبل وظيفة جاوس حيث يمكن تحديد التباين مثل متوسط ميدان الإزاحة (MSD) للجسيمات تتحرك . وهكذا، فإن الخصر وظيفة ارتباط (ويشار أيضا ![]() )، يمكن تعريفها بأنها مجموع الجسيمات المشاكل العضلية الهيكلية والخصر فعال ويمكن قياس نوبة جاوستينغ وظيفة ارتباط لكل تأخير الوقت. ويقاس ط MSD يمكن استخدامها لحساب انتشارية واضح من الجزيئات تتحرك
)، يمكن تعريفها بأنها مجموع الجسيمات المشاكل العضلية الهيكلية والخصر فعال ويمكن قياس نوبة جاوستينغ وظيفة ارتباط لكل تأخير الوقت. ويقاس ط MSD يمكن استخدامها لحساب انتشارية واضح من الجزيئات تتحرك ![]() والتشريد متوسط
والتشريد متوسط ![]() على النحو التالي:
على النحو التالي:
 (3)
(3)
 (4)
(4)
بعض الاعتبارات على الإعداد التجريبية المستخدمة يمكن أن توجه القارئ طوال الأقسام التالية. من أجل إثارة انتقائي fluorophores على الغشاء القاعدي الخلايا سوف نستخدم مجموع التأمل الداخلي (TIR) إضاءة الحية، باستخدام TIR التجاري مضان (TIRF) المجهر (التفاصيل يمكن العثور عليها في قسم المواد). وعلاوة على ذلك، من أجل جمع عشرالبريد مضان سوف نستخدم هدف تضخم عالية (100X NA 1.47، مطلوب الفتحة العددية العالية للإضاءة TIRF) وكاميرا EMCCD (الحجم الفعلي للبكسل على ميكرون رقاقة 16). ليصل إلى حجم بكسل من 100 نانومتر نطبق عدسة التكبير إضافية من 1.6X. كما هو مبين أدناه، ستكون هناك حاجة إلى حل وقت أقل من 1 مللي ثانية لوصف صحيح ديناميات نسبة الدهون في غشاء بسرعة أقل من 100 نانومتر. من أجل الوصول إلى هذا القرار الزماني نحن بحاجة إلى تحديد المنطقة ذات الاهتمام (ROI) أصغر من شريحة كاملة من الكاميرا (512 x 512). في هذه الطريقة، فإن الكاميرا قراءة انخفاض عدد خطوط زيادة القرار الوقت. ومع ذلك، في هذا النظام قراءات الإطار الزمني سيكون محدودا من الوقت اللازم لتحويل الرسوم من التعرض لرقاقة قراءات على الكاميرا وعادة ما تكون في حدود 512 ميلي ثانية لX 512 بكسل EMCCD. للتغلب على هذا الحد، تكنولوجيا ناشئة تتيح تحويل خطوط العائد على الاستثمار فقط بدلا من الإطار كله، ثإيث انخفاض فعالية العملي لحجم رقاقة يتعرض (اقتصاص دعا الاستشعار الوضع في EMCCD لدينا). لهذا التكوين لتكون فعالة، يجب أن تغطي الشريحة خارج العائد على الاستثمار من قبل اثنين من الشقوق التي تقام في مسار بصري. ويمكن تحقيق ذلك بفضل هذا الإعداد قرار الوقت وصولا الى 10 -4 ثواني. يرجى ملاحظة، مع ذلك، أن هذا النهج يمكن أن يقترن مع العديد من الاجهزة التجريبية المختلفة، كما هو موضح في قسم 'نقاش' و.
وسوف تقدم دليلا على الأسلوب في الخلايا الحية، وذلك باستخدام كلا من ATTO488 المسمى 1-بالميتويل-2-هيدروكسي SN -glycero-3-phosphoethanolamine (ATTO488-PPE) ومتغير المسمى GFP من مستقبلات ترانسفيرين (GFP- معدل الخصوبة الإجمالي). في حالة ATTO488-PPE هذا النهج يمكن استرداد بنجاح D التطبيق المستمر تقريبا بوصفها وظيفة من النزوح متوسط يدل على نشر معظمها مجانا، كما ذكرت سابقا 30،35. على النقيض من ذلك، يظهر معدل الخصوبة الإجمالي GFP-D لخفض
Subscription Required. Please recommend JoVE to your librarian.
Protocol
معايرة نظام 1.
- نقطة انتشار وظيفة (PSF) المعايرة
- تمييع 10 ميكرولتر من 30 نانومتر حل حبة الفلورية (حوالي 5 ميكرومتر) في 90 ميكرولتر من الماء المقطر ثم يصوتن الحل لمدة 20 دقيقة. قطع مربع (1 سم × 1 سم) قطعة من هلام الاغاروز (3٪) والودائع 10 ميكرولتر من الحل في الجزء العلوي من هلام. إسقاط قطعة من هلام على الزجاج السفلي من 2 سم طبق بتري والضغط الانخفاض على الزجاج.
- بدوره على الإعداد الاستحواذ، ووضع العينة في حامل، تعيين التعرض الكاميرا وEMgain (100 ميللي ثانية و 1،000 المعلمات هي جيدة ولكن الأمثل وفقا للنظام) وانتظر الكاميرا ليبرد.
- ضبط التعرض الكاميرا إلى 100 ميللي ثانية، وكاميرا EMgain إلى 1،000، ووضع إطار لاكتساب نقل، و 100 التكرار والسيارات حفظ الإعداد.
- باستخدام العدسة والتركيز الضوء المرسل على الحدود من هلام ومن ثم نقل إلى مركز الهدف من هلام، وضبط التركيز وستاغ الإجراء محاذاة الليزر (LAS في AF، اختر 'الإعداد TIRF "واتبع الإجراء محاذاة السيارات).
- العثور على مجال الرؤية مع وجود بقع معزولة واحدة، بدقة التركيز على الفور أكثر إشراقا (التي عادة ما يمثل الخرز مجموع المباراتين) كمرجع، والحصول على 100 لقطة وكرر الخطوة 5-6 مرات من أجل الحصول على العديد من المواقع احدة.
- استيراد سلسلة المكتسبة لبرنامج معالجة البيانات والمتوسط مكدس في الوقت المناسب (الشكل 1A) وحدد حبة واحدة معزولة. رعاية لتحديد أصغر منها لتجنب المجاميع الجسيمات.
- تناسب كثافة التوزيع المحددة (مثال واحد من الخرز الشخصي ويرد في الشكل 1B) مع وظيفة جاوس باستخدام الأمر "gaussfit" (في أدوات ICS-ماتلاب في المواد في Matlab). تحقق الخير من يصلح عن طريق فحص بقايا الحصول عليها (ويرد مثال الشخصي جاوس المجهزة مع مخلفات المقابلة طن الشكل 1B).
- الكاميرا المعايرة
- تشغيل الكاميرا وانتظر الكاميرا ليبرد. تعيين إعداد اقتناء كاميرا، (أي للكاميرا المستخدمة وضعناها التعرض إلى 0.5 ميللي ثانية، وكاميرا EMgain إلى 1،000، وطريقة اكتساب لاقتصاص الوضع، وحجم العائد على الاستثمار إلى 32 × 128، 10،000 التكرار) والبدء في الاستحواذ على الكاميرا الخلفية إشارة.
- استحوذت الواردات سلسلة الإطار إلى برنامج معالجة البيانات. وتفقد حساب متوسط كثافة في كل بكسل من أجل التحقق من أن الكاميرا الخلفية مسطحة تقريبا في المنطقة المحددة للرقاقة. في الوضع المحصولية، وإزالة أولا ومشاركة عدد قليل من خطوط أفقية (3 إلى 10 تبعا لحجم ROI) لكل إطار لعادة منحازة الكاميرا الخلفية في خطوط الحدود.
- إنشاء رسم بياني للقيم (أيضا يعرف المستوى الرقمي، DL) في الصور المكتسبة المكدس (باستخدام الأمر 'اصمت "في Matlab) ورسم لوغاريتممن الناتج التردد (باستخدام القيادة semilogy في Matlab). ويرد مثال على توزيع DL للكاميرا الخلفية في الشكل 2.
ملاحظة: إذا كان يعمل الكاميرا بشكل جيد، سوف المؤامرة تظهر ذروة حوالي جاوس (لمحة مكافئ في نطاق سجل) يمثل توزيع القيم المرتبطة إلى الصفر الفوتون يعقبه الاضمحلال الأسي (خط مع ميل سلبي في نطاق سجل- ) الذي يمثل توزيع القيم المرتبطة إلى 1 الفوتون (الشكل 2). على وجه الخصوص، المركز والتباين وظيفة جاوس تمثل الكاميرا تعويض والخطأ، على التوالي، في حين ثابت الانحلال للجزء الأسي يمثل تقدير DL تعيين بواسطة الكاميرا إلى كل فوتون واحد. في Matlab استخدام قسم "CalibrateCamera" من سيناريو في المواد الداعمة. - كرر هذه العملية لجميع الكاميرا المحددة EMGain والربح.
2. إعتبرإعداد خلية
- لإعداد الليبوزومات اللازمة لدمج 36 الدهون، ويحل بشكل منفصل 1 ملغ من مخدر (1،2-dioleoyl- SN -glycero-3-phosphoethanolamine)، 1 ملغ من DOTAP (1،2-dioleoyl-3-trimethylammonium-البروبان)، و 1 ملغ من PPE-ATTO488 في 1 مل من الكلوروفورم. مزيج معا 0.5 مل من محلول مخدر، و 0.5 مل من محلول DOTAP، و 25 ميكرولتر من الحل PPE-ATTO488 والجافة في ظل فراغ لمدة 24 ساعة. إضافة 0.5 مل من HEPES عازلة 20 ملي، دوامة لمدة 15 دقيقة ويصوتن لمدة 15 دقيقة عند 40 درجة مئوية.
- لإعداد الخلية، ويغسل 3 مرات مع برنامج تلفزيوني طبق P100 من متكدسة CHO-K1 (الصينية الهامستر المبيض)، إضافة 1 مل من التربسين وتخزينها في الحاضنة لمدة 5 دقائق. تعليق خلايا منفصلة مضيفا 9 مل من DMEM / F12 المتوسطة تستكمل مع 10٪ من FBS والبذور 150 ميكرولتر من الحل خلية في طبق بتري تحتوي على 800 ميكرولتر من نفس المتوسطة.
- متجر في الحاضنة لمدة 24 ساعة عند 37 درجة مئوية و 5٪ CO 2. لإدراجها الدهون، تحل محل الخلايا المتوسطة مع 500ميكرولتر من المصل خالية المتوسطة. بعد 30 دقيقة، إضافة 2 ميكرولتر من الجسيمات الشحمية حلا؛ بعد 15 دقيقة يغسل مع الحزب الاشتراكى البرازيلى وإضافة جديدة المتوسطة DMEM / F12 للتصوير.
- لترنسفكأيشن، وخلايا بالنقل وفقا لبروتوكول Lipofectamine (تعليمات الشركة المصنعة) باستخدام معدل الخصوبة الاجمالي-GFP البلازميد ومخزن 24 ساعة في حاضنة قبل التصوير.
3. الحصول على البيانات
- إعداد الإعداد
- من أجل ترموستات المجهر، 24 ساعة قبل التجربة بدوره على الحاضنة.
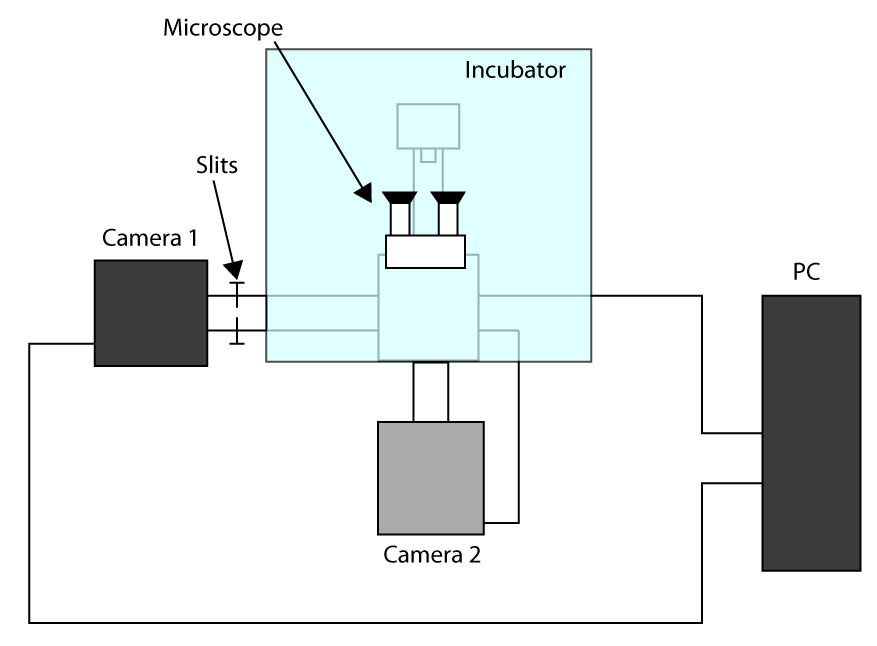
- من أجل تطبيق اكتساب الوقت الأسرع للتحقيق، والعمل في اقتصاص الاستشعار وضع (انظر المقدمة) واستخدام كاميرا الأولى للتصوير (كاميرا 1) والكاميرا الثانية لتحديد الخلية (كاميرا 2). ويرد مخطط التكوين في الإعداد التكميلية الشكل S1. ثم، لتحقيق المواءمة بين اثنين من الكاميرات بدوره على المجهر وانتظر الكاميرات ليبرد.
- تعيين على كل من الكاميرات المعلمات للتصوير الضوء المرسل (أي
- وضع العينات في حامل والتركيز باستخدام العدسة، وإرسال ضوء إلى الكاميرا 1 وبلطف دفع الشقوق السماح الضوء فقط على العائد على الاستثمار المستخدمة في التصوير الخلية (هنا 32 × 32 بكسل ROI).
- نقل الخلايا في المنطقة المحددة وإرسال ضوء إلى الكاميرا 2، ثم رسم العائد على الاستثمار في البرمجيات التي تتحكم الكاميرا 2 من أجل أن يكون مرجعا.
- التصوير (الشكل 3A)
- أولا وقبل كل شيء، محاذاة الليزر TIRF وفقا لإجراءات الإعداد. في الإعداد لدينا، وحدد "الإعداد TIRF" والبدء في إجراء محاذاة السيارات. عندما يتم محاذاة الليزر ضبط 70 نانومتر من عمق الاختراق (حوالي 70 درجة).
- ضبط الوقت التعرض ل70 ميللي ثانية وEMGain إلى 100 على كل كاميرا وكاميرا 2 1. ثم، حدد خلية باستخدام كاميرا 1، ثم ترسل الضوء على كاميرا بدقة 2 والتركيز على غشاء الخلية. تحديد الحد الأدنى للتعرض على كاليفورنياميرا 2، 1،000 EMGain، اقتصاص الاستشعار الوضع، 10 5 التكرار ووضع الحفظ التلقائي كما يناسب الملفات (صورة مرنة نظام النقل، وهو الشكل الذي يمكن بسهولة إدارتها).
- بدء عملية الاستحواذ لتسجيل سلسلة الصورة. الافراج عن الربح والوضع اقتصاص للسماح تثبيت درجة الحرارة قبل الحصول خلية جديدة، ثم كرر الخطوات الماضيين من أجل الحصول على 8-10 الخلايا.
4. احتساب متوسط ميدان المهجرين من التصوير (ط MSD)
ملاحظة: بروتوكول التالية يمكن تطبيقها مباشرة على البيانات الخام. في نفس الوقت، وبروتوكول كله صالحا لعمليات استحواذ بيانات محاكاة سواء في ماتلاب وSimFCS. الارتباط إلى الدروس المقابلة يمكن العثور عليها في قسم "المواد".
- الحساب بواسطة ماتلاب
- استيراد سلسلة المكتسبة إلى ماتلاب باستخدام ImportImageSeries النصي. حساب متوسط كثافة كل صورة في الوقت المناسب باستخدام المشاركmmand يعني على الأبعاد الأولى 2 واستخدام مؤامرة لرؤية ناقلات الناتجة عن ذلك.
- إذا كان أكثر من 10٪ من photobleaching من هو الحاضر، تجاهل سلسلة أو إزالة الجزء الأول منها. إذا كان أقل، في محاولة لتصحيح تأثير على وظيفة ارتباط بطرح كل صورة إلى متوسط شدته، كما هو مبين قبل 37.
- حساب متوسط كثافة كل بيكسل باستخدام الوسط على البعد الثالث، وانظر الصورة الناتجة.
مطلوب اهتمام خاص من أجل تجنب الارتباطات مصطنعة: ملاحظة. في الواقع، كما هو موضح سابقا لتقنيات مشابهة 38، حدود الخلية وكذلك من الحويصلات التركيز يمكن أن يعرض ارتباط قوي. إذا كان التفتيش من الصورة متوسط يكشف حدود الخلية أو من الحويصلات التركيز، في محاولة لاستبعاد المنطقة المعنية خلاف ذلك تجاهل عملية الاستحواذ. لتصحيح تأثير هذه الهياكل متحركة طرح متوسط كثافة الزمنية من كل بكسل 39. - حساب ركان الارتباط الزماني المكاني (G (ξ، χ، τ)) باستخدام الدالة CalculateSTICScorrfunc. إزالة G (ξ، χ، 0) لأن ارتباط بسبب الضوضاء بالرصاص في نظام الإضاءة المنخفضة يهيمن G (0،0،0). العلاقة بسبب كاشف يهيمن على G (± 1،0،0)، وحركة الجسيمات خلال فترة التعرض يمكن أن تشوه G (ξ، χ، τ) لτ = 0 عن طريق زيادة قياس الخصر (هذا التأثير يختفي لτ > 0) 34.
- متوسط G (ξ، χ، τ> 0) استخدام لمرة وبن لوغاريتمي للحد من الضوضاء باستخدام وظيفة "LogBinStack" في دعم المواد ومن ثم تناسب الناتجة G (ξ، χ، τ) باستخدام وظيفة "gaussfit" ل أدوات ICS-ماتلاب في المواد لاسترداد ط MSD (العمود الثاني من المصفوفة الناتجة).
- رسم σ الحصول الخصر (τ) 2 (ط MSD) بوصفها وظيفة من الزمن. إذا كانت البيانات صاخبة جدا، في محاولة لزيادة عدد acquإطارات IRED، وزيادة قوة الليزر، متوسط أكثر G (ξ، χ، τ) معا.
- الحساب بواسطة SimFCS
- فتح ملفات المكتسبة مع يماغيج باستخدام BioFormat المستورد البرنامج المساعد وحفظ سلسلة المكتسبة وتسلسل المشاجرة.
- فتح أداة SimFCS وحدد RICS وحدد ملف> استيراد صور متعددة (التكميلي الشكل S2).
- اختر صالح، أدخل المعلمات اقتناء الصحيحة وإغلاق النافذة مناسبا (التكميلي الشكل S3).
- اختر عرض> متوسط الكثافة> CH1 والتحقق من وجود photobleaching من (التكميلي الشكل S4).
- إذا كان أكثر من 10٪ من photobleaching من هو تجاهل الحاضر سلسلة أو إذا كان من الممكن الحمل مرة أخرى في تسلسل الصور إزالة الجزء الأول من هذه السلسلة.
- إذا تبيض هو أقل من 10٪ اختر الأدوات> ط MSD> تعيين المعلمات، والتحقق من "استخدام المتوسط المتحرك"، تعيين في لوحة دوروا على اليسار وبني مصفر من إطار للمتوسط المتحرك الاهتمام أن الوقت مراسل أعلى من وقت الانتشار المميز (لالجسيمات تتحرك في 1 ميكرون 2 ثانية -1 زمن 10 ثانية هو المتوسط المتحرك جيدا)
- اختر الأدوات> iMSD> احسب ط MSD (الشكل التكميلي S5) ومناسبا وتصدير ط MSD من مذكرة منصة (الشكل التكميلي S6).
5. حساب القانون الانتشار من ط MSD
- تناسب النقاط القليلة الأولى لاستقراء اعتراض (σ 0 2) (5 نقاط وعادة ما تكون كافية ولكن المزيد من النقاط يمكن تركيبها إذا كانت تظهر سلوكا الخطي) ومقارنة هذه القيمة مع قوات الأمن الفلسطينية تقاس سابقا 2. إذا كانت قابلة للمقارنة، وتجري متابعة ديناميات fluorophores معزولة. على النقيض من ذلك، إذا σ 0 2 2 >> PSF محاولة لاكتساب أسرع لضمان أنديناميات لا مخبأة موجودة 34.
- حساب انتشارية واضح (D التطبيق) وتشريد متوسط (R) باستخدام المعادلات 3 و 4 (انظر المقدمة).
- مؤامرة D التطبيق بوصفها وظيفة من R للحصول على نشر القانون مقارنة مع ما يقاس مع اختلاف بقعة استنادا FCS 12 (الشكل 3D).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
من أجل معايرة الخصر فعال، يمكن للصورة واحدة الفلورسنت النانو حبة يكون الإجراء كما هو موضح في البروتوكول خطوة 1.1. تقدم صورة الفلورسنت نموذجية من هذه الخرز في الشكل 1، وتركيب توزيع كثافة بواسطة دالة جاوس 2D يعطي ظهره مخلفات جيدة ويسمح قياس الخصر دور فعال في 270 نانومتر. هذه القيمة هي في اتفاق جيد مع الحد حيود المتوقع حسب تقدير معادلة رايلي. هذه المعايرة ليست ضرورية لقياس ديناميكية الجسيمات ولكن هو مطلوب منها لقياس حجم الجسيمات واضح.
ويرد توزيع الترددات نموذجية من الكاميرا الخلفية في الشكل 2. هي ذروة بحوالي 180 DL بسبب استجابة الكاميرا إلى أي الفوتون، وأنه يمثل مساهمة تماثلي رقمي (AD) تحويل. يمكن تقريب هذه المساهمة بمثابة توزيع جاوس لتقدير تعويض والتباينالتي أدخلها تسجيل إشارة. فوق 200 DL يصبح مستوى التوزيع الرقمي الأسي (خطي في مقياس لوغاريتمي) ويمثل متوسط استجابة الكاميرا لفوتون واحد. تركيب هذا الجزء مع التوزيع الأسي يسمح للقياسات متوسط DL المخصصة لكل فوتون واحد. وكلما هو النسبة بين متوسط DL المخصصة لكل فوتون وتحويل الخطأ م، وسوف يكون أقل ضوضاء في وظيفة ارتباط المحسوبة. وعلاوة على ذلك، فإن متوسط استجابة فوتون واحد يسمح للتقدير النطاق الديناميكي الكاميرا.
وتتلخص رسم تخطيطي لإجراء التجريبي الكامل في الشكل 3، ويمثل صورة Atto488-PPE الإدراج في غشاء في الشكل 4A. ويقدم صورة TIRF تمثيلية من الغشاء القاعدي من خلايا CHO المسمى مع Atto488-PPE في الشكل 4B. قد يكون العديد من النقاط المضيئة الحالي خارج الخلية بسبب لىposomes مكدسة على الزجاج. يمكن التخلص منها عن طريق تحديد العائد على الاستثمار على جزء الغشاء معظمها موحدة في مضان (أي.، الغشاء البلازمي الخلوي). كما هو متوقع القانون نشر قياس (الشكل 4C) لهذا الدهن مسطح، مما يدل على أن نشر معظمها مجانا كما هو موضح سابقا القياسات STED-FCS 30،35. ومن الجدير بالذكر أن كل القيم التشريد المعروضة هي أقل من الحد الحيود، تشير بوضوح قدرة هذا النهج فائقة لحل متوسط التشريد الجزيئية أقل بكثير من الحد حيود ونزولا إلى بضع عشرات من نانومتر.
يتم تمثيل schematization من معدل الخصوبة الاجمالي-GFP ديمر الإدراج في غشاء في الشكل 5A. وأظهرت العديد من الدراسات أن ذيل حشوية هذا مستقبلات تتفاعل مع الهيكل العظمي الغشاء، الذي يعمل بدوره على أنه سياج لل12،40 تنقل مستقبلات. صورة TIRF تمثيلية من خلية CHO معربا عن TFR-GFP هي بريسيnted في الشكل 5B. ينبغي تفضيل خلايا انخفاض كثافة مضان، والغشاء هو أقرب إلى حالة الأم واحتمال القطع الأثرية المتعلقة يتم التقليل من الإفراط في التعبير. بالإضافة إلى ذلك، ينبغي تجنب الجزء المركزي من الخلية، وآثار خارج التركيز مضان (من السيتوبلازم، على سبيل المثال) قد تكون موجودة. كما هو متوقع القانون نشر قياس (الشكل 5C) لمعدل الخصوبة الاجمالي-GFP يظهر أول السلوك شقة أقل من 100 نانومتر، بمتوسط D التطبيق من حوالي 0.7 ميكرون 2 ثانية -1، يليه انخفاض سريع في انتشارية يترتب على ذلك واضح وصولا الى 0.2 ميكرون 2 ثانية -1 (القيمة المقاسة عادة عن طريق FCS 12 محدودة حيود). هذه النتيجة تبين أن نهجنا يمكن بسهولة قياس تشريد متوسط البروتينات GFP المسمى مع قرار من بضع عشرات من نانومتر. وعلاوة على ذلك النطاق المكاني الذي يبدأ D التطبيق لتقليل مجموعات السمةالنطاق المكاني الحبس الجزئي البروتين من الغشاء الهيكل العظمي عند حوالي 120 نانومتر، وذلك تمشيا مع التقديرات السابقة 6.

الرقم 1. معايرة نقطة انتشار وظيفة. (A) Pseudocolor صورة لحبة الخرز ومعزولة المجاميع (B) مؤامرة 3D من ملف كثافة من حبة معزولة يظهر الملف الشخصي جاوس واضحة المعالم. (C) صالح لتوزيع كثافة بواسطة دالة جاوس (اللوحة العليا) مع مخلفات المقابلة (اللوحة السفلى). اتفاق جيد بين التوزيع المجهزة وملف كثافة يقاس هو أيضا دليل على أن قوات الأمن الفلسطينية دورا يمكن أن يقترب من وظيفة جاوس. الرجاء النقر هنا لالسادس EW نسخة أكبر من هذا الرقم.

الشكل 2. معايرة استجابة الكاميرا إلى الفوتونات واحدة. هذا الرقم يدل على المستوى الرقمي (DL) توزيع للكاميرا الخلفية في 32 × 128 العائد على الاستثمار، والتعرض 0.5 ميللي ثانية، في اقتصاص الاستشعار الوضع. ذروة في حوالي 180 DL تمثل استجابة الكاميرا إلى أي الفوتونات. خاصة، لأنها تمثل مساهمة تحويل تماثلي رقمي (AD) ويمكن أن يقترب من وظيفة جاوس لتقدير تعويض والفرق التي أدخلتها تسجيل إشارة. فوق 200 DL توزيع مستويات رقمية يصبح الأسي ويمثل متوسط استجابة الكاميرا لفوتون واحد. قياس هذه المعايير يسمح تقدير كثافة الفوتونات التي يتم تسجيلها خلال عملية الاستحواذ.ق / ftp_upload / 51994 / 51994fig2highres.jpg "الهدف =" _ على بياض "> اضغط هنا لمشاهدة نسخة أكبر من هذا الرقم.

الرقم 3. Schematization هذه الطريقة. (A) على نطاق واسع مجال التصوير بواسطة كاميرا EMCCD يتم تطبيقها للوصول إلى قرار ميلي ثانية واحدة من الباطن، في حين يتم استغلال TIRF microcopy لتوفير باجتزاء بصري دقيق للغشاء البلازما. (B) وربط تلقائيا مكدس الناتجة من الصور من أجل حساب متوسط المكانية -temporal ظيفة الارتباط. هذه الوظيفة ارتباط ويقترب بشكل جيد من قبل وظيفة جاوس (انظر المقدمة) وينتشر في الوقت وفقا لنزوح الجسيمات. (C) وهكذا، وذلك لقياس انتشار وظيفة العلاقة بسبب النزوح الجزيئي، من المناسب مع غاوس يتم تنفيذ وظيفة إيان. وهذا يسمح قياس "القانون نشر" الجزيئية مباشرة من التصوير، في شكل واضح انتشارية مقابل متوسط مؤامرة التهجير. (D) شكرا لهذه المؤامرة، وسائط نشر الجزيئية يمكن تحديدها مباشرة دون الحاجة للنموذج التفسيري أو افتراضات حول التنظيم المكاني للغشاء. في الواقع، فإن جزيئات نشرها بحرية يعرض انتشارية واضح وثابت حركتهم لا تعتمد على مقياس المكاني للقياس. على النقيض من ذلك، فإن الجزيئات محصورة جزئيا تعرض انتشارية واضح ثابت تماما إزاحة أصغر من حجم الحبس، ثم انتشارية لخفض مستويات مكانية أكبر من حجم الحبس. وهكذا، فإن ظهور انخفاض في انتشارية واضح يمكن أن تفسر على أنها بصمة الحبس عابرة، في حين أن النطاق المكاني ذات الصلة يمكن أن تستخدم لتقدير التمديد المكاني للالحبس. .jove.com / الملفات / ftp_upload / 51994 / 51994fig3highres.jpg "الهدف =" _ على بياض "> اضغط هنا لمشاهدة نسخة أكبر من هذا الرقم.

يتم تحديد العائد على الاستثمار (مربع أحمر): الشكل 4. ATTO488-PPE القانون نشر في أغشية الخلايا الحية (A) تمثيل تخطيطي للATTO488-PPE الإدراج في غشاء الخلية (B) صورة TIRF من CHO الغشاء القاعدي المسمى مع ATTO488-PPE. في جزء معظمها الموحد للخلية، وتجنب الحدود الخلية والبقع الفلورسنت عالية. (C) ونشر القانون في قياس العائد على الاستثمار المختارة يظهر السلوك شقة مؤكدا نموذجا نشر مجانا لهذا المكون. الرجاء انقر هنا لمشاهدة نسخة أكبر من هذا الرقم.

يتفاعل الذيل حشوية من مستقبلات مع الهيكل العظمي الغشاء، التي تعمل بمثابة سياج للتنقل مستقبلات (B: الرقم 5. معدل الخصوبة الاجمالي-GFP القانون نشر في أغشية الخلايا الحية (A) تمثيل تخطيطي من معدل الخصوبة الاجمالي-GFP الإدراج في غشاء الخلية. ) صورة TIRF من CHO معربا عن TFR-GFP: يتم تحديد العائد على الاستثمار مفضلا الخلايا معربا منخفضة لتجنب القطع الأثرية بسبب overexpression (C) قانون انتشار معدل الخصوبة الإجمالي (النقاط السوداء)، على عكس PPE (خط رمادي، مأخوذة من الشكل 4). معارض سلوك نموذجي من نشر تقتصر جزئيا حيث يتبع الجزء الأول ثابت عن طريق انخفاض في D التطبيق. الرجاء النقر هنا لمشاهدة نسخة أكبر من هذا فايجوري.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
واحدة تتبع الجسيمات (SPT) يمثل واحدة من الاستراتيجيات الأكثر شيوعا لدراسة ديناميات الجزيئية ولها ميزة كبيرة لقياس مسارات الجسيمات. هذا بدوره يسمح يحقق في سلوك الجسيمات قليلة حتى وصفت في نظام معقد. ومع ذلك، للوصول إلى هذه الميزة SPT يحتاج عادة كثافة منخفضة من التحقيق وملصقات مشرقة جدا. بشكل خاص، للحصول على دقة عالية الزمني (المدى μsec) وعادة ما يطلب التحقيق غير العضوية (مثل نقاط الكم أو الجسيمات النانوية المعدنية): في هذه الحالة إجراء معقد الإنتاج، ووضع العلامات والإدراج في النظام أمر ضروري. مقارنة SPT يبين طريقة الحالي بعض المزايا الرئيسية. أولا وقبل كل شيء، يمكن استخدام هذا النهج في الاقتران مع البروتينات الفلورية. وبالتالي، مقارنة SPT، يتم التوصل إلى القرار الزماني أعلى (على نفس التسمية) بفضل لكمية أقل من الفوتونات المطلوبة 34. أكثر في التفاصيل، هذه الخاصية تتيح دفع RESOL الزمنيution أدناه 10-3 ثانية أيضا عند استخدام البروتينات الفلورية encodable، وهذا يعطي الجدول الزمني وصول خاص إلى ديناميات النانو من مكونات الغشاء. وأخيرا، تجدر الإشارة إلى أن قوانين نشر الجزيئية موصوفة من خلال تحليل الزمكان وظيفة ارتباط كامل، ولا حاجة لتتبع كل جزيء.
المقارنة مع FCS استنادا STED، هي أيضا مثيرة للاهتمام. في قياس STED-FCS يتم قياس متوسط الوقت عبور الجزيئات لخفض كميات المراقبة من قبل الارتباط الزمني للإشارة مضان. وهذا يسمح الحصول على قياس المحلي لديناميات الجزيئية أيضا أقل من الحد الحيود. في النهج قدمت يقاس نشر القانون على أنه متوسط جميع الجسيمات تتحرك في العائد على الاستثمار المحدد، لوحظ من خلال معيار، حيود محدودة، وحجم الملاحظة. ومع ذلك، ذكرت وتظهر النتائج أن هذه الطريقة لا يقتصر بواسطة حيود، ولكن فقط قبل عشرالبريد قرار الزمني المتاح. في الواقع، على الرغم من أن يتم استخدام الاستحواذ محدودة الحيود للكشف عن تقلبات (بالقياس إلى ما يحدث في تقنيات فائقة الدقة أخرى، مثل النخيل وSTORM)، وتشريد الجزيئية أقل بكثير من الحد الحيود يمكن أن يكون (مباشرة) المحسوبة، كما سبق تظاهر باستخدام عصي لقياس تدفقات الجزيئية 32. وعلاوة على ذلك، على عكس STED-FCS، وهذا النهج يمكن تطبيقها بسهولة إلى مجموعة واسعة من الاجهزة المجهري التجارية والقائمة، مثل المجاهر المسح النقطية أو المجاهر القائم على الكاميرا واسعة المجال. ومن الجدير بالذكر أن القياسات STED-FCS قوانين نشر الجزيئية تتطلب حصرا المعايرة المعتمدة على fluorophore من حجم الخصر فعال. معاكس، قدم القياس هنا لا يحتاج الى معايرة نظام (يحتاج فقط لتقدير حجم الجسيمات).
القرار الفعلي في قياس الجسيمات النزوح من الطريقة المعروضة ديpends على مدى دقة يمكننا قياس وظيفة الارتباط. بالتالي، لا يقتصر ذلك بطبيعتها بواسطة حيود، بالقياس إلى حالة SPT حيث يعتمد القرار على مدى دقة يتم قياس الجسيمات "صورة". لقياس ارتباط كبير في أقل من 1 دقيقة للتجارب المقترحة، قليلة الفوتونات (عادة أقل من 10 الفوتونات) لكل جسيم في كل إطار كافية. في الواقع، وبلغ متوسط مساهمة كل الجسيمات المرصودة معا عندما يتم احتساب وظيفة ارتباط، حتى لو لم يتم عزل الجزيئات. هذه الخاصية الجوهرية للطرق ارتباط التذبذب وتسمح باستخدام تسميات قاتمة وكثيفة، مثل البروتينات الفلورية transfected في الخلايا الحية.
ومن هذا المنطلق يبدو واضحا أن النزوح الحد الأدنى للقياس يعتمد على انتشارية من الجسيمات وعلى القرار وقت الإعداد التصوير. كمثال، يرجى النظر في انتشار الجزيئات على غشاء الخلية،حيث أقصى قياس انتشارية للبروتينات أو الدهون حوالي 5 ميكرون 2 ثانية -1. في ظل هذه الظروف، نحن بحاجة إلى قرار من الوقت حوالي 10 -4 ثانية للقبض على النزوح من متوسط 50 نانومتر. ويمكن تحقيق هذا القرار وقت المجاهر المسح السريع على أسس واحدة أو عن طريق كاميرا EMCCD سريعة، حيث يتزامن قرار وقت إلى وقت التعرض، كما أظهر هنا.
شرط أساسي إضافي لهذا الأسلوب لتصف بدقة ديناميات الجزيئية هو أخذ العينات المكانية الصحيحة. في الواقع، وذلك لتناسب وظيفة ارتباط نحتاج إلى أخذ العينات المكانية (حجم بكسل) أقل من الخصر للPSF فعال. في معظم المجاهر التجارية (حقل مبائر أو واسعة)، والخصر PSF يمتد من 200 نانومتر إلى 500 نانومتر (اعتمادا أساسا على الفتحة العددية للهدف المحدد وعلى طول الموجة المستخدمة) ويمكن قياسها بسهولة من قبل النانوية باستخدام تجربة المعايرة حبات الفلورسنت الحجم. THUS، حجم بكسل من 70-150 نانومتر (أقل من 3 مرات الخصر فعال) يمكن أن يكون كافيا. ومع ذلك، يمكن تكييفها حجم بكسل للنظام قيد الدراسة مع مراعاة قاعدة بسيطة: خفض حجم بكسل، أعلى دقة في وصف وظيفة الارتباط. وعلاوة على ذلك، فإن الحد الأدنى لحجم الصورة التي سيتم شراؤها يجب أن يكون على الأقل 3 مرات أكبر من النزوح القصوى من الفائدة (زائد الخصر فعال). هذا أمر مطلوب من أجل التوصل إلى تقارب جيد من الخوارزمية المناسب وأخذ العينات ذات دلالة إحصائية من التشريد الجزيئية. كمثال، لدراسة معدل النزوح الجزيئية أصغر من بضعة مئات نانومتر (على سبيل المثال، 200 نانومتر) حجم صورة من بضعة ميكرونات يكفي. وعلاوة على ذلك، فإن العدد الإجمالي للبكسل (مع الأخذ ثابتة حجم بكسل) الآثار على نوعية وظيفة الارتباط. في الواقع، صورة أكبر يسمح في المتوسط مزيد من المعلومات في وظيفة ارتباط، حتى لو كان على حساب الوقت resolutأيون. بشأن نظام قائم على الكاميرا المستخدمة هنا، يرجى ملاحظة أن حجم الفعلي للبكسل على رقاقة ثابتة. بالتالي، خفض حجم بكسل يقلل من إشارة في كل بكسل (التي تعتمد على مربع من حجم بكسل)، يقلل من مجال الرؤية، ويتطلب ارتفاع قوة التكبير. من ناحية أخرى، في نظام المسح الضوئي، حيث يتم إصلاح منطقة المراقبة، وخفض حجم بكسل يؤدي عادة إلى زيادة عدد بكسل على حساب القرار الوقت.
تفاصيل قليلة عن كاشف المستخدمة يجب أن تناقش. خلافا للكشف عن واحد الفوتون، وأنظمة EMCCD تقيس متوسط كثافة (مستوى الرقمي، DL) التي لا يتناسب طرديا مع ضوء جمعها بسبب وجود إزاحة. حتى لو كان هذا تعويض منخفضة مقارنة النطاق الديناميكي للكاميرا (بضع مئات بالمقارنة مع 2 16 في 16 قراءات بت) وتذكر في التجارب حيث يتم جمع العديد من الفوتونات، فإنه يجب أن يؤخذ في الاعتبار لالحصول على التطبيع الصحيح وظيفة الارتباط. أيضا، يمكن استخدامها كمرجع في ظروف الإضاءة المنخفضة من أجل تحديد كمية إشارة جمع الإزاحة. وعلاوة على ذلك، من أجل تقدير متوسط كمية الفوتونات التي يتم جمعها خلال عملية الاستحواذ، متوسط المستوى الرقمي يرتبط كل فوتون جمعها لابد من قياسه. هذه الكمية يمكن استرجاعها من خلال تعريض الكاميرا إلى كثافة الضوء منخفضة جدا (على سبيل المثال، وعلى ضوء الخلفية في الغرفة). في الواقع، في هذه الحالة، يمكننا أن نفترض معقول أن الفوتونات واحدة فقط تصل إلى الكاميرا، أي قياس كثافة يمكن أن تكون ذات صلة إلى صفر أو واحد الفوتون فقط.
أخيرا، دعونا التعليق على كيف يمكن استخدام بعض النظم اكتساب البديل (أي الاجهزة المجهري مختلفة) لإجراء القياسات المقدمة. أولا وقبل كل شيء، "W'factor في المعادلة 2 (التي تمثل الارتباط الذاتي من PSF فعال) يمكن تكييفها لرانه معين نظام الحصول على استخدامها وذلك لتناسب وظيفة ارتباط التجريبية. كما هو موضح سابقا 34 حالة سهلة لاكتساب مثقال ذرة مجهر المسح بالليزر عندما تكون سرعة المسح الضوئي هو أعلى بكثير من ديناميات الجسيمات. في مثل هذه الحالة، في الواقع، حركة الجسيمات خلال اكتساب الوقت (أي وقت خط) يمكن اعتبار يذكر وظيفة ارتباط ويقترب بشكل جيد من قبل وظيفة جاوس. في سياق تقنيات التصوير الناشئة، ويستند هذا النهج المثير للاهتمام حول إمكانية لإنتاج صفائح رقيقة جدا ضوء (1-2 ميكرون) من خلال عينة 41. ورقة الخفيفة تسمح الإضاءة انتقائية من طائرة واحدة (واحدة إضاءة الطائرة المجهري، SPIM) في العينة و، جنبا إلى جنب مع نظام اكتساب القائم على الكاميرا، باجتزاء بصري سريع في 3D 42. بسبب هذه الخصائص، وقد مترافق SPIM بنجاح مع FCS 43 ويمكن أن تمثل فاليد أداة لتمديد التحليل المقدم إلى بيئات 3D، مثل السيتوبلازم أو النواة من الخلايا الحية.
يلخص، من نقطة التجريبية نظر هذا النهج يتطلب سوى الوصول إلى المجهر مجهزة مع وحدة اكتساب سريع. البروتين من الفائدة يمكن الموسومة ب أي بروتين فلوري أو fluorophore العضوية، مما يسمح أيضا للتصوير متعدد الألوان. في هذا السياق، فإننا نتصور إمكانية استخدام العابرة للتحليل ط MSD لتحديد السكان الفرعي من الجزيئات وتكشف التفاعلات وشارك في نشرها على أغشية الخلايا الحية. وأخيرا، فإننا نعتقد أن هذا النهج قد يمثل أداة قوية لدراسة البروتينات و / أو الدهون تمر التقسيم الديناميكي داخل nanodomains على غشاء البلازما. في هذه الحالة، وحجم عالية متغير وعمر nanodomains إدخال مستوى إضافي من التعقيد في البيانات الحقيقية التي تتطلب المزيد من التطبيقات المنهجية بما في ذلك التصوير 2 لون والمحليةتحليل (على سبيل المثال، 2D ارتباط الزوج) و / أو تباين مضان.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}