

Abstract
Det har blivit alltmer uppenbart att den geografiska fördelningen och rörelse membrankomponenter såsom lipider och proteiner är nyckelfaktorer i regleringen av många cellulära funktioner. Men på grund av de snabba dynamiken och de små strukturer är involverade, är en mycket hög spatio-temporal upplösning som krävs för att fånga den verkliga beteendet hos molekyler. Här presenterar vi det experimentella protokollet för att studera dynamiken i fluorescensmärkta plasmamembran proteiner och lipider i levande celler med hög Spatiotemporal upplösning. Noterbart är, denna strategi behöver inte spåra varje molekyl, men det beräknar befolknings beteende med hjälp av alla molekyler i en viss region av membranet. Utgångspunkten är en snabb avbildning av en viss region på membranet. Efteråt är en komplett tid och rum autokorrelationsfunktion beräknas korrelerar förvärvade bilder öka tidsfördröjningar, exempelvis vardera 2, 3, n upprepningar. Det är möjligt att visa att breddeni toppen av de spatiala autokorrelationsfunktionen ökar vid ökande tidsfördröjning som funktion av partikelrörelse på grund av diffusion. Därför passar den serie av autokorrelationsfunktioner gör det möjligt att extrahera den faktiska proteinmedelkvadrat förskjutning från imaging (iMSD), här i form av sken diffusivitet vs genomsnittlig förskjutning. Detta ger en kvantitativ bild av de genomsnittliga dynamiken i enstaka molekyler med nanometer precision. Genom användning av en GFP-märkt variant av transferrinreceptorn (TfR) och en ATTO488 märkt 1-palmitoyl-2-hydroxi-sn-glycero-3-fosfoetanolamin (PPE) är det möjligt att observera spatiotemporal regleringen av protein och lipid diffusion på im stora membran regioner i mikro-till-milli-sekund tidsintervall.
Introduction
Med utgångspunkt från den ursprungliga "vätske mosaik" modell av Singer och Nicolson, bilden av cellplasmamembranet har kontinuerligt uppdaterats under de senaste decennierna för att inkludera den framväxande roll cytoskeleton och lipid domänerna 1,2.
De första observationerna erhölls genom fluorescerande återhämtning efter fotoblekning (FRAP) avtäckningen att en betydande fraktion av membranproteiner är orörlig 3-5. Dessa banbrytande studier, även om mycket informativt, led av den relativt dåliga upplösningen i rymden (mikron) och tid (sekunder) i FRAP inställningar. Dessutom är en ensemble medelvärdesmätning, FRAP saknar i att ge information om enda molekyl beteende.
I detta sammanhang är det möjligt att specifikt märka en enda molekyl med mycket ljusa taggar (möjliggör studiet av diffusionsprocessen en molekyl i taget) har varit mycket framgångsrikt. Speciellt, genom att trycka påtidsupplösning av den inre Particle Tracking (SPT) inställning till mikrosekunder tidsskalan, Kusumi, et al. fått tillgång till okända funktioner i lipid och proteindynamik som kraftigt bidragit till erkännandet av den roll aktin baserade membranskelett i membranfysiologi 6 , 7. Dessa fynd genererade den så kallade "strejkvakt och staket" modell, där lipid och protein diffusion regleras av aktin baserade skelettet. Men för att få tillgång till den enorma mängd information som SPT många experimentella frågor måste lösas. Speciellt är märkningsförfarandet vanligen består av många steg som produktion, rening och införande av de märkta arter i systemet. Dessutom stora etiketter, som kvantprickar eller metallnanopartiklar, ofta krävs för att nå under millisekund tid och tvärbindningen av målmolekyler med etiketten kunde inte undvikas i många fall. Slutligen, många banormåste registreras för att passa statistiska kriterier och samtidigt en låg densitet av etiketten krävs för att möjliggöra spårning.
Jämfört med SPT, fluorescens-korrelationsspektroskopi (FCS), övervinna många av dessa nackdelar, är ett mycket lovande tillvägagångssätt för att studera molekylär dynamik. I själva verket fungerar FCS bra även med dim och täta etiketter, som gör det möjligt att studera dynamiken i fluorescerande protein-märkta molekyler i gående transfekterade celler. Dessutom möjliggör det att nå höga statistik i en begränsad tid. Slutligen, trots den "höga" densitet av etiketter FCS erbjuder enstaka molekyler information. Tack vare alla dessa egenskaper, FCS är en mycket enkel metod och har tillämpats i stor utsträckning för att studera lipid och protein dynamik både i modellmembran och i levande-celler 8-10. Många olika metoder har föreslagits för att öka förmågan hos FCS att avslöja detaljerna i molekylär diffusion. Till exempel var det shegna som genom att utföra FCS på olika stora observationsområdena kan man definiera en "FCS diffusion lag" upplysande dolda funktioner i molekylär rörelse 11,12. Förutom att vara varierade i storlek, var det prioriterade området också dupliceras 13, flyttade i rymden längs linjerna 14-20 eller konjugerade med snabba kameror 21,22. Med hjälp av dessa "tid och rum" korrelation tillvägagångssätt, var relevanta biologiska parametrar för flera membrankomponenter kvantitativt beskrivas både modellmembran och faktiska biologiska sådana, vilket således ger inblick i membran rumslig organisation.
Men i alla FRAP och FCS-program hittills beskrivna storleken på det prioriterade området utgör en gräns i rumslig upplösning som inte kan övervinnas. Flera super upplösning avbildningsmetoder har nyligen utvecklats för att kringgå denna gräns. En del bygger på lokalisering precision, till exempel stokastiska optisk rekonstruktion mikroskopi (STORM) <sup> 23,24, foto lokalisering mikroskopi (PALM) 25, fluorescens PALM (FPALM) 26, och single-partikel tracking PALM (sptPALM) 27: den relativt stora mängd fotoner som krävs på varje stillbild, men begränsar den tid upplösning dessa metoder till åtminstone flera millisekunder, vilket hämmar deras tillämplighet in vivo.
Däremot har ett lovande alternativ för superupplösning avbildning öppnats av rumsligt modulera fluorescens emission med stimulerade utarmning utsläppsmetoder (STED eller reversibla mätt optiska fluorescens övergångar (RESOLFT)) 28,29. Dessa tillvägagångssätt kombinerar utformningen av observationsvolymen långt under diffraktionsgränsen med möjlighet att använda snabb scanning mikroskop och detekteringssystem. I kombination med fluorescensfluktuationer analys, STED mikroskopi tillåtet att direkt söka av nanoskala Spatiotemporal dynamik lipider och proteins i levande cellmembran 30,31.
Samma fysiska kvantiteter av STED baserad mikroskopi kan erhållas genom en modifierad tid och rum bildkorrelationsspektroskopi (STICS 32,33) metod som är lämplig för att studera dynamiken i fluorescerande-märkta membranproteiner och / eller lipider i levande celler och genom en kommersiell mikroskop. Den experimentella protokollet presenteras här består av några få steg. Den första kräver en snabb avbildning av regionen av intresse på membranet. Därefter resulte trave bilder används för att beräkna den genomsnittliga rumsliga-temporala korrelationsfunktioner. Genom att montera den serie av korrelationsfunktioner, kan den molekylära "diffusion lag" erhållas direkt från avbildning i form av en skenbar diffusivitet (D app) - vs -Genomsnittlig förskjutning tomt. Denna tomt är kritiskt beroende miljön utforskas av molekylerna och tillåter att erkänna direkt själva spridningssättav lipid / protein av intresse.
I mer detaljer, som tidigare visats 34, den tid och rum autokorrelationsfunktion för den förvärvade bildserie beror kritiskt på dynamiken i molekylerna rör sig i den insamlade bildserien (observera att samma resonemang kan tillämpas på en rad förvärv där bara en dimension i rymden anses). Särskilt definierar vi korrelationsfunktionen som:
 (1)
(1)
där  representerar den uppmätta fluorescensintensiteten i den position x, y och vid tiden t,
representerar den uppmätta fluorescensintensiteten i den position x, y och vid tiden t, ![]() och
och ![]() representerar avståndet i x-ochy-riktningarna respektive,
representerar avståndet i x-ochy-riktningarna respektive, ![]() representerar viss tid och
representerar viss tid och ![]() representerar genomsnittet. Denna funktion kan uttryckas som:
representerar genomsnittet. Denna funktion kan uttryckas som:
 (2)
(2)
där "N" representerar det genomsnittliga antalet molekyler i området observation, ![]() representerar konvolution drift i rymden, och
representerar konvolution drift i rymden, och  representerar autokorrelationen för den instrumen midjan. Detta senare kan tolkas som ett mått på hur fotoner från en enda sändare är utspridda i rymden på grund av att den optiska / inspelning setup (den så kallade punktspridningsfunktionen, PSF, genrally väl approximeras med en Gauss-funktion). Slutligen
representerar autokorrelationen för den instrumen midjan. Detta senare kan tolkas som ett mått på hur fotoner från en enda sändare är utspridda i rymden på grund av att den optiska / inspelning setup (den så kallade punktspridningsfunktionen, PSF, genrally väl approximeras med en Gauss-funktion). Slutligen  representerar sannolikheten för att finna en partikel på ett avstånd
representerar sannolikheten för att finna en partikel på ett avstånd ![]() och
och ![]() efter en tidsfördröjning
efter en tidsfördröjning ![]() . Om vi betraktar en diffusiva dynamik, där partiklar rör sig slumpmässigt i alla riktningar och nettoflöden inte är närvarande, är denna funktion också väl approximeras med en Gauss-funktion där variansen kan identifieras som Mean Square Displacement (MSD) i rörliga partikeln . Sålunda midjan på korrelationsfunktionen (även kallat
. Om vi betraktar en diffusiva dynamik, där partiklar rör sig slumpmässigt i alla riktningar och nettoflöden inte är närvarande, är denna funktion också väl approximeras med en Gauss-funktion där variansen kan identifieras som Mean Square Displacement (MSD) i rörliga partikeln . Sålunda midjan på korrelationsfunktionen (även kallat ![]() ), Kan definieras som summan av de partikel muskuloskeletala sjukdomar och den instrument midja och kan mätas genom en gaussisk passformting av korrelationsfunktionen för varje tidsfördröjning. Den uppmätta i MSD kan användas för att beräkna en skenbar diffusiviteten hos de rörliga molekyler
), Kan definieras som summan av de partikel muskuloskeletala sjukdomar och den instrument midja och kan mätas genom en gaussisk passformting av korrelationsfunktionen för varje tidsfördröjning. Den uppmätta i MSD kan användas för att beräkna en skenbar diffusiviteten hos de rörliga molekyler ![]() och en genomsnittlig förskjutning
och en genomsnittlig förskjutning ![]() som:
som:
 (3)
(3)
 (4)
(4)
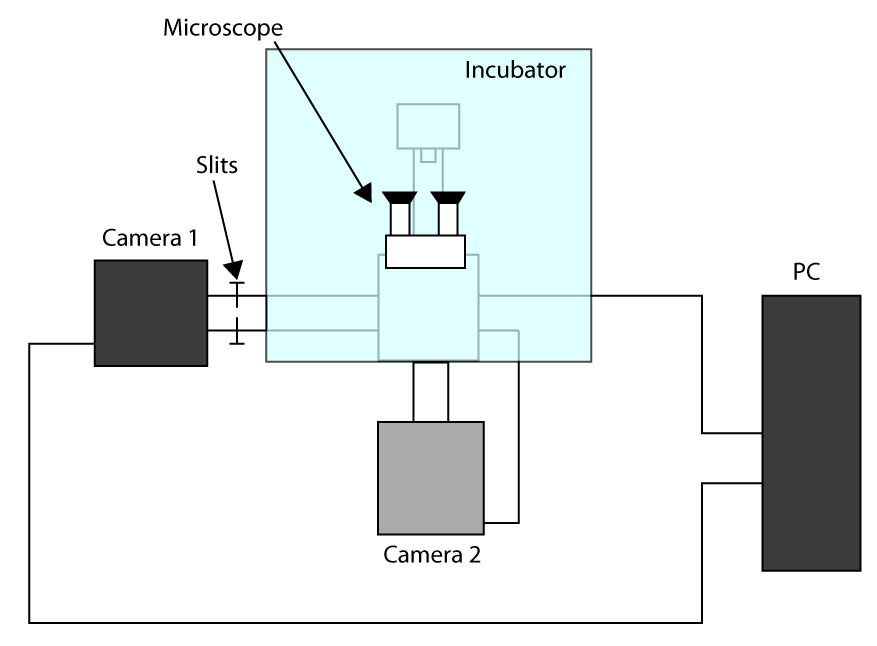
Några överväganden på experimentuppställning som används kan vägleda läsaren genom de följande avsnitten. För att selektivt excitera fluoroforema på basalmembranet av levande celler som vi kommer att använda en total inre reflektion (TIR) belysning, med hjälp av ett kommersiellt TIR fluorescens (TIRF) mikroskop (detaljer finns i materialet avsnitt). Dessutom, för att samla in the fluorescens vi kommer att använda en hög förstoring mål (100X NA 1,47, är hög numerisk apertur som krävs för TIRF belysning) och en EMCCD kamera (fysiska storleken på pixel på chipet 16 um). För att nå en pixelstorlek på 100 nm tillämpar vi en extra förstoringslins 1,6X. Såsom diskuteras nedan, skulle en tidsupplösning under en ms krävas för att korrekt beskriva dynamiken i snabba membranlipider nedanför 100 nm. För att uppnå denna tidsupplösning måste vi välja ett område av intresse (ROI) som är mindre än hela chipet av kameran (512 x 512). På detta sätt kommer kameran läsa ett reducerat antal linjer ökande tidsupplösningen. Men i denna avläsning regim ramtiden skulle begränsas av den tid som krävs för att flytta de avgifter från exponering för utläsningschipset på kameran och är oftast i storleksordningen millisekunder för 512 x 512 pixel EMCCD. För att slå denna gräns, kan en ny teknik flytta ROI-linjerna i stället för hela ramen, wed en praktisk effektiv minskning av den exponerade chip storlek (kallas Cropped Sensor läge i vårt EMCCD). För denna konfiguration för att vara effektiv, måste chipet utsidan av ROI täckas av ett par slitsar som är monterade i den optiska banan. Tack vare denna inställning en tidsupplösning ned till 10 -4 sekunder kan uppnås. Observera dock att detta tillvägagångssätt kan kopplas till många olika experimentella uppställningar, som förklaras i "diskussionen" avsnittet.
Demonstration av metoden kommer att tillhandahållas i levande celler, genom att använda både en ATTO488 märkt 1-palmitoyl-2-hydroxi-sn-glycero-3-fosfoetanolamin (ATTO488-PPE) och en GFP-märkt variant av transferrinreceptorn (GFP- TfR). I fallet med ATTO488-PPE kan detta tillvägagångssätt lyckades återställa en nästan konstant D appen som en funktion av genomsnittlig förskjutning indikerande en mestadels fri diffusion, såsom tidigare rapporterats 30,35. Däremot visar TfR-GFP en minskande D
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1 System Kalibrering
- Punktspridningsfunktion (PSF) kalibrering
- Späd 10 | il av 30 nm fluorescerande pärla lösning (ca 5 | iM) i 90 | il destillerat vatten och därefter sonikera lösningen under 20 min. Skär en kvadrat (1 cm x 1 cm) stycke av agarosgel (3%) och insättning 10 | il av lösningen på toppen av gelén. Störta den bit av gelen på botten glaset i en 2 cm petriskål och pressa droppe på glaset.
- Slå på förvärvs setup, placera provet i hållaren, ställ in kameran exponering och EMgain (100 msek och 1000 är bra parametrar, men optimera enligt det systemet) och vänta tills kameran har svalnat.
- Ställ kamerans exponering till 100 ms, kamera EMgain till 1000, förvärvsläge till Frame Transfer, 100 upprepning och automatisk spara inställningen.
- Genom att använda okularet och genomlysning fokus på gränsen av gelén och sedan flytta målet till mitten av gelen, justera fokus och start laserinriktningsförfarandet (i LAS AF, välj 'TIRF setup' och följ autojusteringsförfarandet).
- Hitta ett synfält med isolerade enstaka fläckar, exakt fokusera på den ljusare plats (som representerar oftast pärlor aggregat) som referens, förvärvar 100 bilder och upprepa steg 5-6 gånger för att förvärva flera enstaka fläckar.
- Importera förvärvad till ett databehandlingsprogram och genomsnitt stapeln i tiden (figur 1 A) och välj en isolerad kula. Var noga med att välja ut de allra minsta för att förhindra partikelaggregat.
- Montera den valda intensitetsfördelningen (ett exempel på enstaka pärlor profil visas i Figur 1 B) med en Gaussisk funktion med hjälp av kommandot "gaussfit" (i ICS-Matlab verktyg Materialet i Matlab). Verifiera godhet passningen genom att inspektera de erhållna residualer (ett exempel på monterade gaussprofil med motsvarande rester presenteras in Figur 1B).
- Kamera kalibrering
- Slå på kameran och vänta tills kameran har svalnat. Ställ inställnings kamera förvärv, (dvs. om den för den använda kameran vi ställa in exponeringen till 0,5 msek, kamera EMgain till 1000, förvärvsläge till Cropped läget, ROI storleken till 32 x 128, 10000 repetitioner) och börja förvärvet av kamerans bakgrunden signalen.
- Importera förvärvade ram serie till ett databehandlingsprogram. Beräkna och inspektera den genomsnittliga intensiteten i varje bildpunkt för att kontrollera att kameran bakgrund är approximativt plant i det valda området av chipet. I Cropped läge, tar bort det första och de sista horisontella linjer (3 till 10, beroende på storleken på ROI) för varje ram eftersom kameran bakgrunden brukar partisk i gränslinjerna.
- Skapa ett histogram över värdena (även definierat Digital Level, DL) i förvärvade bilder stack (med hjälp av kommandot "historik" i Matlab) och plotta logaritmenav resulterande frekvens (med semilogy kommando i Matlab). Ett exempel på DL fördelning för kamera bakgrund presenteras i figur 2.
OBS: Om kameran fungerar bra, kommer tomten visar en ungefär Gaussisk topp (en parabolisk profil i log-skala) representerar fördelningen av värden associerade till noll foton följt av en exponentiellt avtagande (en linje med negativ lutning i log-skala ) som representerar fördelningen av värden associerade till 1 foton (Figur 2). I synnerhet centrum och variansen för den gaussiska funktionen representerar kamerans offset och fel, respektive, medan sönderfallskonstanten av den exponentiella delen representerar en uppskattning av den DL tilldelats av kameran till varje enda foton. I Matlab använda avsnittet "CalibrateCamera" av Script i stödmaterial. - Gör motsvarande för alla valda Camera EMGain och Gain.
2. MärktCellberedning
- För att framställa liposomer som krävs för lipid inkorporering 36, upplösa sig 1 mg DOPE (1,2-dioleoyl- sn-glycero-3-fosfoetanolamin), 1 mg DOTAP (1,2-dioleoyl-3-trimetylammonium-propan), och 1 mg av PPE-ATTO488 i 1 ml kloroform. Blanda 0,5 ml DOPE-lösning, 0,5 ml DOTAP-lösning och 25 | il av PPE-ATTO488 lösning och torka under vakuum under 24 timmar. Lägg 0,5 ml HEPES-buffert 20 mM, vortex under 15 minuter och sonikera under 15 minuter vid 40 ° C.
- För att förbereda cellen, tvätta 3 gånger med PBS en p100 maträtt av sammanflytande CHO-K1 (kinesisk hamster), tillsätt 1 ml trypsin och butik i inkubator i 5 min. Suspendera lösgjorda celler lägga 9 ml DMEM / F12-medium kompletterat med 10% av FBS och ympa 150 | il av cell lösning i en petriskål med 800 pl av samma medium.
- Förvaras i inkubatorn under 24 h vid 37 ° C och 5% CO2. För lipid bolagsordning, byt cell medium med 500il av serumfritt medium; efter 30 minuter, tillsätt 2 pl av liposomer lösning; efter 15 min tvätt med PSB och lägga till nya DMEM / F12 medium för avbildning.
- För transfektion, transfektera celler enligt Lipofectamine protokollet (tillverkarens anvisningar) med hjälp av TfR-GFP plasmid och lagra 24 timmar i kuvös innan avbildning.
3 Data Acquisition
- Beredning Setup
- För att termostat mikroskopet, 24 timmar före försöket slå på inkubatorn.
- För att tillämpa den snabbaste uppnåeanskaffningstiden, arbete i Cropped Sensor läge (se inledningen) och använder en första kamera för avbildning (kamera 1) och andra kamera för att markera cellen (kamera 2). Ett system för installationskonfigurationen presenteras i kompletterande figur S1. Sedan, för att rikta in två kameror sväng på mikroskopet och vänta kamerorna svalna.
- Ställ på båda kamerorna parametrarna för genomlysning imaging (dvs.
- Placera proverna i hållaren och fokusera med okular, skicka ljus till kameran 1 och tryck försiktigt slitsarna som ljuset endast på ROI används för cell imaging (här ett 32 x 32 bildpunkter ROI).
- Flytta en cell i det markerade området och skicka ljuset till kameran 2, sedan rita en ROI i programvaran som styr kameran 2 för att få en referens.
- Imaging (figur 3A)
- Först och främst rikta TIRF laser enligt proceduren för din konfiguration. I vår inställning, väljer du "TIRF inställning" och starta den automatiska justeringsförfarandet. När lasern är inriktad inställd 70 nm av penetrationsdjup (cirka 70 °).
- Ställ exponeringstid till 70 msek och EMGain till 100 på både kamera 1 och kamera 2; sedan markerar du en cell med hjälp av kamera 1, sedan skicka ljuset på kamera 2 och noggrant fokusera cellmembranet. Ställ in minsta exponering på camera 2, 1000 EMGain, Cropped sensor-läge, 10 5 repetitioner och ställa autosave som passar filer (Flexible Image Transport System, ett format som lätt kan hanteras).
- Starta förvärvet att spela in bilden serien. Släpp Gain och den beskurna läge att låta temperaturstabilisering innan förvärva en ny cell och upprepa de sista två stegen för att förvärva 8-10 celler.
4 Beräkning av Mean Square Förskjutning från Imaging (i MSD)
OBS: Följande protokoll kan direkt appliceras på rådata. Samtidigt är hela protokollet gäller för uppgifter förvärv simulerade både i Matlab och i SimFCS. Kopplingen till de motsvarande handledning kan hittas i "Material" sektion.
- Beräkning av Matlab
- Importera förvärvad till Matlab med hjälp ImportImageSeries manus. Beräkna den genomsnittliga intensiteten för varje bild i tiden med hjälp av command menar på de första 2 dimensioner och använda tomten för att se den resulterande vektorn.
- Om mer än 10% av fotoblekning är närvarande, kassera serien eller ta bort den första delen av dem. Om den är lägre, försöka rätta effekten på korrelationsfunktionen genom att subtrahera varje bild sin genomsnittliga intensitet, som visas före 37.
- Beräkna den genomsnittliga intensiteten för varje pixel med hjälp av medelvärdet på den tredje dimensionen och se resulterande bilden.
OBS: Särskild uppmärksamhet krävs för att undvika artefaktuella korrelationer. I själva verket, som tidigare visats för liknande tekniker 38, cellramar samt av fokus vesiklar skulle kunna införa en stark korrelation. Om inspektionen av den genomsnittliga bilden avslöjar cellramar eller oskarp vesiklar, försök att utesluta inblandade annars kasta förvärvsområdet. För att korrigera effekten av denna orörliga strukturer subtrahera den genomsnittliga tids intensitet från varje pixel 39. - Beräkna tHan spatiotemporal korrelation (G (ξ, χ, τ)) med hjälp av funktionen CalculateSTICScorrfunc. Ta bort G (ξ, χ, 0), eftersom korrelationen grund skottet brus i svagt ljus regimen dominerar G (0,0,0); korrelationen beror på detektorn dominerar G (± 1,0,0) och partikelrörelse under exponeringstiden kan deformera G (ξ, χ, τ) för τ = 0 genom att öka den uppmätta midjan (denna effekt försvinner för τ > 0) 34.
- Genomsnittlig G (ξ, χ, τ> 0) med hjälp av en logaritmisk tidsfack för att minska bullret genom att använda "LogBinStack" funktion i underlag och sedan montera den resulte G (ξ, χ, τ) med hjälp av funktionen "gaussfit" av ICS-Matlab verktyg i material att återvinna i MSD (den andra kolumnen i den resulterande matrisen).
- Plotta den erhållna midja σ (τ) 2 (i MSD) som en funktion av tiden. Om uppgifterna är för bullrig, försök att öka antalet acquiRed ramar, ökar lasereffekten, i genomsnitt mer G (ξ, χ, τ) tillsammans.
- Beräkning av SimFCS
- Öppna de förvärvade filer med ImageJ använder BioFormat importör plugin och spara förvärvad som Tiff sekvens.
- Öppen SimFCS och välj RICS verktyget och välj Arkiv> Importera Flera bilder (kompletterande figur S2).
- Välj Fit, sätt in rätt insamlingsparametrar och stänga passnings fönstret (kompletterande figur S3).
- Välj Visa> Genomsnittlig Intensitet> CH1 och verifiera förekomsten av fotoblekning (kompletterande figur S4).
- Om mer än 10% av fotoblekning är närvarande kasse serien eller om det är möjligt lasten igen bildsekvensen att avlägsna den första delen av serien.
- Om blekning den är lägre än 10% väljer Verktyg> i MSD> inställda parametrar, kolla 'Använd glidande medelvärde ", som i ROI panelen till vänster enumbra av ramen för det glidande medelvärdet att uppmärksamma att korrespondentntiden är högre än den karakteristiska diffusionstiden (för partikel som rör sig med en xm 2 sek -1 en tid av 10 sek är en god glidande medelvärde)
- Välj Verktyg> iMSD> Beräkna i MSD (kompletterande figur S5) och passform och exportera i MSD från minnesanteckningar (kompletterande figur S6).
5. Beräkning av Diffusion lagen från den i: MSD
- Montera de första punkter att extrapolera intercept (σ 0 2) (5 poäng är oftast tillräckligt men fler poäng kan monteras om de visar ett linjärt beteende) och jämför detta värde med det tidigare uppmätta PSF 2. Om de är jämförbara, är dynamiken i enstaka fluoroforer följs. Om däremot σ 0 2 >> PSF 2 försök att skaffa snabbare attinga dolda dynamiken är närvarande 34.
- Beräkna den sken diffusivitet (D app) och den genomsnittliga förskjutningen (R) med hjälp av ekvationerna 3 och 4 (se Inledning).
- Plot D appen som en funktion av R för att erhålla en diffusion lag jämförbar med vad som mäts med fläck variation baserad FCS 12 (figur 3D).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
För att kalibrera den instrument midjan, kan bilden av en fluorescerande nano pärla vara åtgärd som beskrivs i protokoll steg 1.1. En typisk fluorescerande bild av dessa pärlor presenteras i Figur 1. Montering av intensitetsfördelningen från en 2D Gaussfunktion ger tillbaka goda rester och låter mäta instrumen midjan vid 270 nm. Detta värde är i god överensstämmelse med den förväntade diffraktionsgränsen uppskattas genom Rayleigh-ekvationen. Denna kalibrering inte är nödvändig för mätning av partikeldynamik men det erfordras att mäta den skenbara partikelstorleken.
En typisk fördelning av kamera bakgrundsfrekvensen presenteras i figur 2. Toppen vid ca 180 DL är beroende på kamera svar på att ingen fotonen, och det representerar bidraget av analog digital (AD-omvandlare). Detta bidrag kan approximeras som en Gaussisk fördelning för att uppskatta offset och variansenintroduceras av signalinspelning. Över 200 DL digital distribution nivån blir exponentiell (linjär i logaritmisk skala) och representerar den genomsnittliga kamera svar på en enda foton. Montering denna del med en exponentiell fördelning tillåter mätningar av den genomsnittliga DL delas varje enda foton. Ju högre är förhållandet mellan den genomsnittliga DL delas varje fotonen och AD-omvandlaren fel, kommer den nedre vara bullret i det beräknade korrelationsfunktionen. Dessutom tillåter den genomsnittliga enda foton gensvar uppskattningen av kameran dynamiskt område.
Ett diagram över den kompletta experimentella proceduren är sammanfattad i Figur 3 och en bild av Atto488-PPE insättning i membranet är representerat i figur 4A. Ett representativt TIRF bild av det basala membranet av en CHO-celler märkta med Atto488-PPE presenteras i figur 4B. Flera ljuspunkter kan finnas utanför cellen på grund av liposomes staplade på glaset. De kan kastas genom att välja en ROI på en membranparti mestadels enhetliga i fluorescens (dvs., Cellplasmamembran). Som väntat den uppmätta diffusion lagen (Figur 4C) för denna lipid är platt, vilket indikerar en mestadels fri diffusion som tidigare visat STED-FCS mätningar 30,35. Det är värt att nämna att alla visade förskjutningsvärdena ligger under diffraktionsgränsen, som tydligt anger förmågan hos denna inställning till super lösa medelmolekyl förskjutningar långt under diffraktionsgränsen och ner till några tiotal nanometer.
En schematisering av TfR-GFP dimer insättning i membranet representeras i figur 5A. Många studier har visat att den cytoplasmatiska svansen av denna receptor interagerar med membran skelett, som i sin tur verkar som ett stängsel för receptorn rörlighet 12,40. En representant TIRF bild av en CHO-cell som uttrycker TfR-GFP är presented i figur 5B. Låg fluorescensintensitets celler bör föredra, eftersom membranet är närmare den nativa tillstånd och sannolikheten för artefakter i samband med överexpression minimeras. Dessutom bör den centrala delen av cellen undvikas, eftersom effekterna av out-of-fokus fluorescens (från cytoplasman, till exempel) kan vara närvarande. Som väntat den uppmätta diffusion lag (figur 5C) för TfR-GFP visar ett första platt beteende nedanför 100 nm, med en genomsnittlig D applikation av ca 0,7 ^ m 2 sek -1, följt av åtföljande snabb minskning i skenbar diffusivitet ned till 0,2 | im 2 sek -1 (värdet typiskt uppmätt genom diffraktionsbegränsad FCS 12). Detta resultat visar att vårt tillvägagångssätt enkelt kan mäta den genomsnittliga förskjutningen av GFP-märkta proteiner med en upplösning av några tiotal nanometer. Dessutom den rumsliga skalan där D appen börjar sjunka sätter den karakteristiskarumslig skala av protein partiell inneslutning av membranskelett på runt 120 nm, i linje med tidigare beräkningar 6.

Figur 1 Kalibrering av punktspridningsfunktion. (A) pseudobild av en isolerad bead och pärlor aggregat. (B) 3D plot av intensitetsprofilen av en isolerad kula visar en väldefinierad gaussisk profil. (C) Fit av intensitetsfördelningen genom en gaussisk funktion (övre panel) med motsvarande restprodukter (nedre fältet). Den goda avtal mellan monterad fördelning och den uppmätta intensitetsprofilen är också ett bevis på att den instrument PSF kan approximeras med en gaussisk funktion. Klicka här till vi EW en större version av denna siffra.

Figur 2 Kalibrering av kamera svar på enstaka fotoner. Figuren visar den digitala nivå (DL) distributionen för kamera bakgrund i en 32 x 128 ROI, exponering 0,5 msek i Cropped sensor-läge. Toppen vid ca 180 DL representerar kameran svar på några fotoner. Särskilt utgör det bidrag Analoga Digitala (AD-omvandlare) och kan approximeras med en Gauss-funktion för att uppskatta offset och variansen införts av signalinspelnings. Över 200 DL distribution av digitala nivåer blir exponentiell och representerar den genomsnittliga kamera svar på en enda foton. Mätningen av dessa parametrar tillåter att uppskatta tätheten av fotoner som registrerats under förvärvet.s / ftp_upload / 51994 / 51994fig2highres.jpg "target =" _blank "> Klicka här för att se en större version av denna siffra.

Figur 3 schematisering av metoden. (A) Brett fält avbildning av EMCCD kamera används för att nå sub-millisekund upplösning, medan TIRF microcopy utnyttjas för att ge exakt optisk sektionering av plasmamembranet. (B) Den resulterande trave bilder är autokorrelerade för att beräkna den genomsnittliga rumsliga -temporal korrelationsfunktion. Denna korrelationsfunktion är väl approximeras med en gaussisk funktion (se inledningen) och den sprider ut i tid enligt partikel förskjutningar. (C) Således, för att kvantifiera spridningen av korrelationsfunktionen på grund av molekylär förskjutning, passande med ett Gauss ian funktionen utförs. Detta gör det möjligt att mäta den molekylära "diffusion rätten" direkt från bild, i form av sken diffusivitet vs genomsnittlig förskjutning tomt. (D) Tack vare denna tomt, kan molekylär diffusion lägen direkt identifieras utan behov av ett tolkningsmodell eller antaganden om spatial organisation av membranet. I själva verket kommer fritt diffunderar molekylerna visar en konstant skenbar diffusivitet som deras rörlighet inte beror på den rumsliga omfattningen av mätningen. Däremot kommer delvis trånga molekyler visa ett ganska konstant skenbar diffusivitet för förskjutningar mindre än förlossningen storlek, sedan en minskande diffusivitet för rumsliga skalor större än förlossningen storlek. Sålunda kan uppträdandet av en reduktion i den skenbara diffusivitet tolkas som ett fingeravtryck av transient inneslutning, medan tillhörande rumslig skala kan användas för att uppskatta den rumsliga utvidgningen av instängdhet. .jove.com / filer / ftp_upload / 51994 / 51994fig3highres.jpg "target =" _blank "> Klicka här för att se en större version av denna siffra.

Figur 4 ATTO488-PPE diffusion lag i levande cellmembran (A) Schematisk bild av ATTO488-PPE insättning i cellmembranet (B) TIRF bild av CHO basalmembran märkt med ATTO488-PPE:.. A ROI (röd ruta) väljs i en mestadels enhetlig del av cellen, undvika cell gränsen och mycket fluorescerande fläckar. (C) Spridning lagen mäts i den valda ROI visar en platt beteende bekräftar en fri diffusion modell för denna komponent. Klicka här för att se en större version av denna siffra.

Figur 5 TfR-GFP diffusion lag i levande cellmembran (A) Schematisk representation av TfR-GFP insättning i cellmembranet:. Den cytoplasmatiska svansen av receptorn interagerar med membran skelett, som fungerar som ett stängsel för receptor rörlighet (B. ) TIRF bild av CHO uttrycker TfR-GFP: a ROI väljs föredrar låga uttryckande celler för att undvika artefakter på grund av överuttryck (C) Den diffusion lag TfR (svarta prickar), till skillnad från PPE (grå linje, tagen från figur 4),. visar det typiska beteendet för delvis inneslutna diffusion där en första plan del följs av en minskning i D app. Klicka här för att se en större version av denna figur.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Spårning enda partikel (SPT) är en av de vanligaste strategierna för att studera molekyldynamik och har den stora fördelen att mäta partikelbanor. Detta i sin tur gör det möjligt att sondera beteende ens några märkta partiklar i ett komplext system. Men för att nå denna fördel SPT behöver normalt en låg densitet av sonden och mycket ljusa etiketter. Särskilt för att få hög tidsupplösning (ps intervall) oorganiska sond krävs ofta (t.ex. kvantprickar eller metallnanopartiklar): i detta fall en komplicerad procedur för produktion, märkning och insättning i systemet är nödvändig. Jämfört med SPT den nuvarande metoden visar några viktiga fördelar. Först av allt, kan användas för detta tillvägagångssätt i konjugering med fluorescerande proteiner. Således jämfört med SPT, är en högre tidsupplösning uppnås (på samma etikett) tack vare det lägsta av fotoner krävs 34. Mer i detalj, gör den här egenskapen trycka den temporala resolmepannor under 10 - 3 sek även vid användning av kodningsbara fluorescerande proteiner, och detta tidsskala ger exklusiv tillgång till nano dynamik membran beståndsdelar. Slutligen är det värt att notera att molekylär diffusion lagarna beskrivs genom att analysera hela rumtiden korrelationsfunktion, utan att behöva spåra varje molekyl.
Jämförelsen med STED Baserade FCS är också intressant. I en STED-FCS mätning genomsnittstransittiden för molekyler för minskande observations volymer mätes genom den tidsmässiga korrelationen av fluorescenssignalen. Detta tillåter att erhålla en lokal mätning av molekyldynamik också nedan diffraktionsgränsen. I det presenterade tillvägagångssätt diffusion lagen mäts som genomsnittet av alla partiklar som rör sig i det valda ROI, observeras med hjälp av standard, diffraktionsbegränsad, observation volym. Emellertid rapporterade resultat visar att denna metod inte är begränsad av diffraktion, men endast av the tids upplösning. Faktum är att även en diffraktionsbegränsad förvärvet används för att detektera fluktuationer (analogt till vad som görs i andra superupplösningstekniker, såsom PALM och STORM), molekylära förskjutningar långt under diffraktionsgränsen kan vara (direkt) beräknat, vilket redan visas genom användning STICS att mäta molekylär flöden 32. Till skillnad STED-FCS, detta tillvägagångssätt kan lätt appliceras på ett brett utbud av kommersiella och befintliga mikroskopi inställningar, till exempel raster scanning mikroskop eller breda fältkamerabaserade mikroskop. Det är värt att nämna att STED-FCS mätningar av molekylär diffusion lagar kräver absolut en fluorofor beroende kalibrering av storleken på den instrumen midjan. Motsatt, presenterade mätningen här kräver inte ett kalibreringssystem (behövs endast för beräkning av partikelstorlek).
Den faktiska upplösningen i mätningen av partikel förskjutningar av den presenterade metoden deberor av hur noggrant vi kan mäta korrelationsfunktionen. Följaktligen är det inte i sig begränsad av diffraktion, analogt med SPT fall där upplösningen beror på hur noggrant partikeln "bild" mätes. För att mäta en signifikant korrelation på mindre än 1 min för de experiment, några fotoner (vanligen under 10 fotoner) för varje partikel i varje ram är tillräckligt. I själva verket är det bidrag från alla de observerade partiklarna genomsnitt tillsammans när korrelationsfunktionen beräknas, även om partiklar inte är isolerade. Den här egenskapen är inneboende av fluktuation korrelationsmetoder och möjlighet finns att använda dim och täta etiketter, till exempel fluorescerande proteiner transfekterade i levande celler.
Med detta i åtanke verkar det klart att den minsta mätbara förskjutningen beror på diffusivitet av partikeln och på den tid upplösning på bildinställningar. Som ett exempel kan du överväga diffusion av molekyler på cellmembranet,där den maximala uppmätta diffusivitet för proteiner eller lipider är cirka 5 pm 2 sek -1. Under dessa förhållanden, behöver vi en tidsupplösning på ca 10 -4 sek för att fånga en genomsnittlig förskjutning av 50 nm. Denna tidsupplösning kan uppnås genom snabba scanningsmikroskop längs enkla linjer eller genom snabb EMCCD kamera, där tidsupplösning sammanfaller med exponeringstiden, som visade här.
En ytterligare viktig förutsättning för denna metod för att exakt beskriva molekyldynamik är en korrekt rumslig provtagning. I själva verket, för att passa korrelationsfunktionen behöver vi en spatial sampling (pixelstorlek) lägre än midjan på instrument PSF. I de flesta kommersiella mikroskop (konfokala eller brett fält), spänner över PSF midjan från 200 nm till 500 nm (beroende främst på den numeriska bländaröppningen för det valda målet och på våglängden som används) och kan enkelt mätas med en kalibreringsexperiment med hjälp av nano stora fluorescerande pärlor. Thus, kan en pixelstorlek på 70-150 nm (3 gånger lägre än den instrument midjan) räcka. Däremot kan pixelstorleken anpassas till det system som studeras med hänsyn till en enkel regel: sänka pixelstorlek, större noggrannhet i beskrivningen av korrelationsfunktionen. Dessutom den minsta storleken för den bild som ska förvärvas ska vara minst 3 gånger större än den maximala förskjutningen av intresse (plus den instrument midjan). Detta krävs för att nå en god samstämmighet i beslaget algoritm och en statistiskt signifikant sampling av molekylära förskjutningar. Som ett exempel, för att studera medelmolekyl förskjutningar mindre än några hundra nanometer (t.ex. 200 nm) är nog en bildstorlek på några mikrometer. Dessutom det totala antalet pixlar (med konstant pixelstorleken) effekter på kvaliteten på korrelationsfunktionen. I själva verket gör en större bild genomsnitt mer information i korrelationsfunktionen, även om på bekostnad av tiden Resolution. När det gäller kamerabaserat system som används här, observera att den fysiska storleken på pixeln på chipet är fast. Följaktligen minskar pixelstorleken sänker signalen i varje pixel (som beror på kvadraten av pixelstorlek), minskar synfältet, och kräver högre förstoringen. Å andra sidan, i ett avsökningssystem, där observationsområdet är fast, minskar pixelstorleken resulterar vanligen i ett ökat antal pixel på bekostnad av tidsupplösningen.
Några detaljer om detektorn används måste diskuteras. Till skillnad från enkel-fotondetektorer, EMCCD system mäter en genomsnittlig intensitet (digital nivå, DL) som inte är direkt proportionell mot det insamlade ljuset på grund av närvaron av en offset. Även om denna förskjutning är lågt jämfört med det dynamiska området för kameran (några hundra jämfört med 2 16 i 16 bitarsvärde) och försumbar i experiment där många fotoner samlas, måste det tas i beaktande attfå en korrekt normalisering av korrelationsfunktionen. Även förskjutningen kan användas som en referens i svagt ljus för att identifiera hur mycket signalen samlas. I syfte att uppskatta den genomsnittliga mängden fotoner som samlas under förvärvet, den genomsnittliga digitala nivån associerad till varje insamlade fotonen måste mätas. Denna kvantitet kan hämtas genom att exponera kameran till en mycket låg ljusintensitet (t.ex. bakgrunden ljus i rummet); i själva verket, i detta fall, kan vi rimligen anta att bara enstaka fotoner når kameran, dvs den uppmätta intensiteten kan relateras till enbart noll eller en foton.
Låt oss slutligen kommentera hur några alternativa anskaffningssystem (dvs olika mikroskopi inställningar) kan användas för att utföra de presenterade mätningarna. Först av allt, kan den "W'factor i ekvation 2 (som representerar autokorrelationen för den instrument PSF) anpassas till than särskilt förvärvssystem som används för att passa den experimentella korrelationsfunktionen. Som tidigare visats 34, är ett enkelt fall förvärvet dugg en laserskanningsmikroskop när avsökningshastigheten är betydligt högre än partikeldynamik. I ett sådant fall, i själva verket, rörelsen av partiklar under förvärvstiden (dvs., rad tid) kan anses vara försumbar och korrelationsfunktionen är väl approximeras med en Gauss-funktion. I samband med de nya avbildningstekniker, är en intressant strategi som bygger på möjligheten att framställa mycket tunna ljus ark (1-2 pm) genom provet 41. Ljuset fliken kan selektiv belysning av ett enda plan (ett plan belysning mikroskopi, SPIM) i provet och i kombination med ett kamerabaserat insamlingssystem, snabb optisk sektione i 3D 42. På grund av dessa egenskaper har SPIM framgångsrikt konjugerad med FCS 43 och skulle kunna utgöra ett valid verktyg för att förlänga den presenterade analys till 3D-miljöer, såsom cytoplasman eller kärnan i levande celler.
Sammanfatta, från en experimentell synvinkel denna metod kräver endast tillgång till ett mikroskop utrustat med en snabb förvärvsmodul. Proteinet av intresse kan märkas med någon fluorescerande protein eller organiska fluoroforen, vilket möjliggör också för flerfärgsbildframställning. I detta sammanhang föreställer vi möjligheten att använda kors i MSD-analys för att välja delpopulationer av molekyler och avslöja interaktioner och co-diffusion av levande cellmembran. Slutligen anser vi att detta tillvägagångssätt kan utgöra ett kraftfullt verktyg för att studera proteiner och / eller lipider som genomgår dynamisk partitionering inom nanodomains på plasmamembranet. I det här fallet, den mycket varierande storlek och livslängd nanodomains införa ytterligare en nivå av komplexitet i verkliga uppgifter som skulle kräva ytterligare metodologiska implementeringar inklusive 2-färgavbildning, lokalanalysen (t ex 2D par korrelation) och / eller fluorescensanisotropi.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}