

Abstract
זה הפך להיות ברור יותר ויותר כי הפריסה המרחבית והתנועה של רכיבי קרום כמו שומנים וחלבונים הם גורמי מפתח בוויסות של פונקציות רבות תאיות. עם זאת, בשל הדינמיקה מהירה והמבנים הזעירים המעורבים, נדרשת רזולוציה מרחב ובזמן גבוהה מאוד כדי לתפוס את ההתנהגות האמיתית של מולקולות. כאן אנו מציגים את פרוטוקול הניסוי ללימוד הדינמיקה של חלבוני פלזמה קרום fluorescently שהכותרת ושומנים בתאים חיים עם רזולוציה spatiotemporal גבוהה. יש לציין, גישה זו אינה צריכה לעקוב אחר כל מולקולה, אך היא מחשבת התנהגות אוכלוסייה תוך שימוש בכל המולקולות באזור נתון של הקרום. נקודת המוצא היא הדמיה מהירה של אזור נתון על הממברנה. לאחר מכן, פונקצית autocorrelation מרחב ובזמן מוחלטת מחושבת מקשרת תמונות שנרכשו בהגדלת עיכובי זמן, למשל כל 2, 3, n חזרות. ניתן להוכיח כי הרוחבשל השיא של עליות פונקצית autocorrelation מרחבית בהגדלת עיכוב זמן כפונקציה של תנועת חלקיקים עקב דיפוזיה. לכן, ראוי של הסדרה של פונקציות autocorrelation מאפשר לחלץ את החלבון בפועל מתכוון עקירה מרובעת מההדמיה (iMSD), כאן מוצג בצורה של diffusivity לכאורה נגד עקירה ממוצעת. זה מניב מבט כמותית של הדינמיקה הממוצעת של מולקולות בודדות עם דיוק ננומטר. על ידי שימוש בגרסת GFP-tagged של transferrin קולטן (TFR) וATTO488 המסומנת 1-palmitoyl-2-הידרוקסי sn -glycero-3-phosphoethanolamine (PPE) ניתן לצפות ברגולצית spatiotemporal של דיפוזיה חלבון והשומנים על אזורי קרום מיקרומטר בגודל בטווח זמן מיקרו ל-מילי שניות.
Introduction
החל מהמודל המקורי "פסיפס נוזל" על ידי זינגר וניקולסון, התמונה של קרום פלזמה סלולארי כבר מעודכנת באופן רציף במהלך העשורים האחרונים על מנת לכלול את התפקיד המתפתח של תחומים שלד תא ושומנים בדם 1,2.
התצפיות הראשונות התקבלו על ידי שחזור ניאון לאחר photobleaching הסרת לוט (FRAP) שחלק ניכר של חלבונים בממברנה הוא נייח 3-5. מחקריו החלוציים הללו, למרות שמאוד אינפורמטיבי, סבלו מעניה יחסית ברזולוציה במרחב (מיקרון) וזמן (שניות) של מערכי FRAP. כמו כן, להיות מדידת מיצוע הרכב, FRAP חסר במתן מידע על מולקולה בודדת התנהגות.
בהקשר זה, את האפשרות לתייג מולקולה בודדת עם תגים בהירים מאוד (המאפשרים מחקר של המולקולה אחת תהליך דיפוזיה בכל פעם) במיוחד היה מוצלח מאוד. במיוחד, על ידי לחיצה עלרזולוציה של הגישה יחידה חלקיקים מעקב (SPT) ללוח הזמנים מיקרו, Kusumi, et גישה צברה אל. לתכונות ידועות של דינמיקת שומנים וחלבונים שתרמו רב להכרה בתפקידו של שלד קרום מבוסס אקטין בפיזיולוגיה קרום 6 זמן , 7. ממצאים אלה שנוצרו מה שנקרא 'כלונסאות וגדר' מודל, שבו דיפוזיה שומנים והחלבונים מוסדרת על ידי שלד מבוסס אקטין. עם זאת, על מנת לקבל גישה לכמות העצומה של מידע המסופק על ידי נושאים רבים ניסיוניים SPT יש לטפל. במיוחד, הליך התיוג מורכב בדרך כלל על ידי צעדים רבים כמו ייצור, טיהור והכנסה של המינים שכותרתו לתוך המערכת. יתר על כן, חברות תקליטים גדולות, כמו נקודות קוונטיות או חלקיקי מתכת, נדרשות לעתים קרובות כדי להגיע ללוח זמנים משנה אלפית השנייה וcrosslinking של מולקולות המטרה על ידי התווית לא ניתן היה להימנע במקרים רבים. לבסוף, מסלולים רביםיש שיירשם כדי שיתאים לקריטריונים סטטיסטיים ובו זמנית בצפיפות נמוכה של התווית נדרש כדי לאפשר מעקב.
בהשוואה לSPT, ספקטרוסקופיה מתאם הקרינה (FCS), להתגבר על רבים מחסרונות אלה, מייצגת גישה מאוד מבטיחה ללמוד דינמיקה מולקולרית. למעשה, FCS עובד היטב גם עם תוויות עמומות וצפופות, מה שמאפשר ללמוד את הדינמיקה של מולקולות מתויג חלבון פלואורסצנטי בתאי transfected הזמני. כמו כן, זה מאפשר להגיע לסטטיסטיקה גבוהה בכמות מוגבלת של זמן. לבסוף, למרות הצפיפות "הגבוהה" של תוויות FCS מספק מידע מולקולות בודדות. תודה לכל המאפיינים האלה, FCS מייצגת גישה מאוד פשוטה ויושם בהרחבה ללמוד דינמיקת שומנים וחלבונים הן בממברנות מודל ובשידור חיים תאים 8-10. גישות רבות ושונות הוצעו כדי להגביר את היכולת של FCS כדי לחשוף את הפרטים של דיפוזיה מולקולרית. לדוגמא, זה היה shשלו כי על ידי ביצוע FCS באזורי תצפית שונה בגודל אחד יכול להגדיר תכונות "חוק דיפוזיה FCS" מאיר עיניים מוסתרות של תנועה המולקולרית 11,12. חוץ מזה שהיה שונה בגודל, באזור המוקד היה גם משוכפל 13, עבר בחלל לאורך קווי 14-20 או מצומדות עם מצלמות מהירות 21,22. באמצעות המתאם 'מרחב ובזמן "אלה גישות, פרמטרים ביולוגיים רלוונטיים של מספר מרכיבי קרום תוארו כמותית משני קרומי מודל ואלה ביולוגיים בפועל, ובכך תובנה מניבות לארגון המרחבי קרום.
עם זאת, בכל FRAP ויישומי FCS שתואר עד כה בגודל של אזור המוקד מייצג מגבלה ברזולוציה מרחבית שלא ניתן להתגבר עליו. שיטות הדמיה ברזולוציה סופר כמה פותחו לאחרונה כדי לעקוף מגבלה זו. חלקם מבוססים על דיוק לוקליזציה, כגון מיקרוסקופיה אופטית סטוכסטיים שחזור (STORM) <sup> 23,24, מיקרוסקופיה לוקליזציה photoactivation (PALM) 25, PALM הקרינה (FPALM) 26, וPALM מעקב חלקיק יחיד (sptPALM) 27: הכמות הגדולה יחסית של פוטונים הנדרשים בכל תמונה, לעומת זאת, מגביל את הרזולוציה של הזמן שיטות אלה לפחות כמה אלפיות השניה, ובכך פוגעות תחולתם in vivo.
בניגוד לכך, חלופה מבטיחה להדמיה ברזולוציה סופר נפתח על ידי מרחבית ויסות פליטת הקרינה עם שיטות מגורה דלדול פליטה (STED או מעברים הפיכים saturable אופטי הקרינה (RESOLFT)) 28,29. גישות אלה משלבים עיצוב תצפית הנפח נמוך בהרבה מהמגבלה העקיפה עם האפשרות להשתמש מיקרוסקופי סריקה מהירה ומערכות איתור. בשילוב עם ניתוח תנודות הקרינה, מיקרוסקופיה STED אפשרה לחקור ישירות את דינמיקת spatiotemporal ננומטריים של שומנים וproteins בקרום תא חי 30,31.
ניתן להשיג את אותו כמויות הפיזיות של מיקרוסקופיה מבוססת STED ידי ספקטרוסקופיה הותאם מרחב ובזמן מתאם תמונה (STICS 32,33) שיטה שמתאימה ללימוד הדינמיקה של חלבוני fluorescently- מתויג קרום ו / או שומנים בתאים חיים ועל ידי מיקרוסקופ מסחרי. פרוטוקול הניסוי המובא כאן מורכב על ידי כמה צעדים. הראשון דורש הדמיה מהירה של האזור של עניין על הממברנה. לאחר מכן, הערימה של תמונות וכתוצאה מכך משמשת לחישוב פונקציות מתאם מרחב וזמן הממוצע. על ידי התאמת הסדרה של פונקציות מתאם, ניתן להשיג "חוק דיפוזיה 'המולקולרי ישירות מהדמיה בצורה של diffusivity לכאורה (אפליקציה D) - לעומת עלילת עקירת -average. עלילה זו באופן ביקורתי תלויה בסביבה נבחנה על ידי המולקולות ומאפשרת הכרה ישירות מצבי דיפוזיה בפועלשל השומנים בדם / החלבון של עניין.
בפרטים נוספים, כפי שמוצג 34 בעבר, פונקצית קורלציה אוטומטית מרחב ובזמן של סדרת תמונה שנרכשה באופן ביקורתי תלויה בדינמיקה של המולקולות נעות בסדרת התמונה שנאסף (יש לציין כי באותו ההיגיון ניתן להחיל ברכישת אונליין שבו רק ממד אחד בחלל נחשב). בפרט, אנו מגדירים את פונקצית המתאם הגבוהה ככל:
 (1)
(1)
בי  מייצג את עוצמת הקרינה שנמדדה בx העמדה, y ובזמן t,
מייצג את עוצמת הקרינה שנמדדה בx העמדה, y ובזמן t, ![]() ו
ו ![]() מייצג את המרחק בx וכיווני y בהתאמה,
מייצג את המרחק בx וכיווני y בהתאמה, ![]() מייצג את פער הזמן, ו
מייצג את פער הזמן, ו ![]() מייצג את הממוצע. פונקציה זו יכולה לבוא לידי ביטוי כמו:
מייצג את הממוצע. פונקציה זו יכולה לבוא לידי ביטוי כמו:
 (2)
(2)
שבו 'N' מייצגת את המספר הממוצע של מולקולות באזור התצפית, ![]() מייצגת את פעולת פיתול בחלל, ו
מייצגת את פעולת פיתול בחלל, ו  מייצג את autocorrelation של המותניים אינסטרומנטלי. זה אחרון יכול להתפרש כמדד של כמה הפוטונים של פולט אחד פרושים במרחב בשל ההתקנה האופטית / הקלטה (פונקצית התפשטות הנקודה שנקרא, כוחות הביטחון הפלסטיניים, גןעצרת מקורב היטב על ידי פונקצית גאוס). לבסוף,
מייצג את autocorrelation של המותניים אינסטרומנטלי. זה אחרון יכול להתפרש כמדד של כמה הפוטונים של פולט אחד פרושים במרחב בשל ההתקנה האופטית / הקלטה (פונקצית התפשטות הנקודה שנקרא, כוחות הביטחון הפלסטיניים, גןעצרת מקורב היטב על ידי פונקצית גאוס). לבסוף,  מייצג את ההסתברות למצוא חלקיק במרחק
מייצג את ההסתברות למצוא חלקיק במרחק ![]() ו
ו ![]() לאחר השהיה
לאחר השהיה ![]() . אם ניקח בחשבון את דינמיקת diffusive, שבו חלקיקים לנוע באופן אקראי לכל הכיוונים ונתיבים נטו אינם נוכחים, פונקציה זו גם מקורב היטב על ידי פונקצית גאוס בי השונות יכולות להיות מזוהות כממוצעת כיכר העקירה (MSD) של החלקיקים המרגשים . לפיכך, המותניים של פונקצית המתאם (המכונה גם כ
. אם ניקח בחשבון את דינמיקת diffusive, שבו חלקיקים לנוע באופן אקראי לכל הכיוונים ונתיבים נטו אינם נוכחים, פונקציה זו גם מקורב היטב על ידי פונקצית גאוס בי השונות יכולות להיות מזוהות כממוצעת כיכר העקירה (MSD) של החלקיקים המרגשים . לפיכך, המותניים של פונקצית המתאם (המכונה גם כ ![]() ), יכול להיות מוגדר כסכום של MSDS החלקיקים ומותני אינסטרומנטלי וניתן למדוד על ידי גאוס בכושרטינג של פונקצית המתאם עבור כל עיכוב זמן. אני MSD נמדד ניתן להשתמש כדי לחשב diffusivity לכאורה של המולקולות נעות
), יכול להיות מוגדר כסכום של MSDS החלקיקים ומותני אינסטרומנטלי וניתן למדוד על ידי גאוס בכושרטינג של פונקצית המתאם עבור כל עיכוב זמן. אני MSD נמדד ניתן להשתמש כדי לחשב diffusivity לכאורה של המולקולות נעות ![]() ועקירה ממוצעת
ועקירה ממוצעת ![]() כ:
כ:
 (3)
(3)
 (4)
(4)
שיקולים כמה בהגדרת הניסוי נעשה שימוש יכול להנחות את הקורא בכל הסעיפים הבאים. על מנת לרגש באופן סלקטיבי fluorophores על הקרום הבסיסי של תאי חיים נשתמש תאורה כוללת ההשתקפות פנימית (TIR), תוך שימוש בקרינת TIR מסחרי מיקרוסקופ (TIRF) (ניתן למצוא את פרטים בסעיף החומר). יתר על כן, על מנת לאסוף את ההקרינה דואר נשתמש מטרת הגדלה גבוהה (100x NA 1.47, צמצם מספרי גבוה נדרש לתאורת TIRF) ומצלמת EMCCD (גודל פיזי של פיקסל על מיקרומטר שבב 16). כדי להגיע לגודל פיקסל של 100 ננומטר אנחנו מיישמים עדשת הגדלה נוספת של 1.6x. כפי שנראה להלן, תידרש זמן ברזולוציה מתחת ל -1 msec כדי לתאר כראוי את הדינמיקה של שומני קרום מהירים להלן 100 ננומטר. על מנת להגיע לפתרון זמני זה אנחנו צריכים לבחור אזור של העניין (ROI) קטן יותר מכל השבב של המצלמה (512 x 512). בדרך זו, המצלמה תהיה לקרוא מספר מופחת של קווי הגדלת הרזולוציה הזמן. עם זאת, במשטר קריאה זו מסגרת הזמן יהיה מוגבל על ידי הזמן הנדרש כדי להעביר את החיובים מהחשיפה לשבב הקריאה על המצלמה והיא בדרך כלל בסדר הגודל של אלפיות שנייה לEMCCD 512 x 512 פיקסל. להכות מגבלה זו, טכנולוגיה מתפתחת מאפשרת הסטת ROI-הקווים רק במקום כל המסגרת, with הפחתה יעילה מעשית של הגודל נחשף שבב (הנקרא חיישן קצוץ מצב בEMCCD שלנו). לתצורה זו תהיה אפקטיבית, השבב מחוץ להחזר על ההשקעה חייב להיות מכוסה על ידי כמה חריצים רכובים בנתיב האופטי. הודות להתקנה זו רזולוציה זמן עד ל10 -4 שניות יכולים להיות מושגת. שים לב, עם זאת, כי גישה זו יכולה להיות בשילוב עם setups ניסיוני רב ושונה, כפי שהוסברה בסעיף "הדיון".
הפגנה של השיטה תסופק בתאים חיים, על ידי השימוש בשתי ATTO488 מסומן 1-palmitoyl-2-הידרוקסי sn -glycero-3-phosphoethanolamine (ATTO488-PPE) וגרסה שכותרתו GFP של קולטן transferrin (GFP- TFR). במקרה של ATTO488-PPE גישה זו יכולה לשחזר בהצלחה יישום D כמעט קבוע כפונקציה של תזוזה ממוצעת המצביעה על דיפוזיה לרוב ללא תשלום, כפי שדווחה בעבר 30,35. בניגוד לכך, TFR-GFP תערוכות D יורד
Subscription Required. Please recommend JoVE to your librarian.
Protocol
כיול מערכת .1
- פונקצית מורחים נקודת כיול (כוחות הביטחון הפלסטיניים)
- לדלל 10 μl של 30 פתרון חרוז ניאון ננומטר (כ -5 מיקרומטר) ב90 μl של מים מזוקקים ולאחר מכן sonicate הפתרון עבור 20 דקות. חותך ריבוע (1 סנטימטר x 1 סנטימטר) חתיכת agarose ג'ל (3%) והפיקדון 10 μl של הפתרון בחלק העליון של הג'ל. להפוך את פיסת ג'ל על הזכוכית התחתונה של 2 סנטימטר צלחת פטרי ולסחוט את הטיפה על הזכוכית.
- הפעל את תוכנית ההתקנה של הרכישה, לשים המדגם בעל, להגדיר את חשיפת המצלמה וEMgain (100 אלפיות שניים ו -1,000 הם פרמטרים טובים, אבל לייעל בהתאם למערכת) ולחכות למצלמה כדי להתקרר.
- חשיפת מצלמה מוגדרת 100 אלפיות שני, EMgain מצלמה ל1,000, מצב רכישה למסגרת העברה, 100 חזרות ואוטומטיות לשמור את ההגדרה.
- שימוש בעינית ופוקוס אור מועבר על גבול ג'ל ולאחר מכן להעביר את המטרה למרכז של ג'ל, לשנות את הפוקוס וstart הליך יישור לייזר (בלאס AF, בחר 'התקנת TIRF' ובצע את הליך היישור האוטומטי).
- מצא את שדה הראייה עם כתמים מבודדים אחת, מדויק להתמקד בנקודה בהירה (כי בדרך כלל מייצג המצרפי חרוזים) כנקודת התייחסות, לרכוש 100 מסגרות ולחזור על השלב 5-6 פעמים כדי לרכוש כמה נקודות בודדות.
- לייבא את הסדרה נרכשה לתכנית עיבוד נתונים וממוצע הערימה בזמן (איור 1 א) ובחר חרוז בודד אחד. תשמור על עצמך כדי לבחור את הקטן ביותר, כדי למנוע אגרגטים חלקיקים.
- התאם את התפוצה שנבחרה בעוצמה (דוגמא לפרופיל חרוזים בודד מוצגת באיור 1 ב ') עם פונקצית גאוס באמצעות הפקודה "gaussfit" (בכלי ICS-Matlab בחומרים בMatlab). ודא טוב לנכון על ידי בדיקת השאריות שהושגו (דוגמא לפרופיל גאוס המצויד עם השאריות המקבילה מוצגת in איור 1 ב).
- כיול מצלמה
- הפעל את המצלמה ולחכות למצלמה כדי להתקרר. הגדרת רכישת המצלמה להגדיר, (כלומר, למצלמה המשמשת לנו להגדיר את החשיפה ל0.5 אלפית שני, EMgain מצלמה ל1,000, מצב רכישה לקצוץ מצב, גודל ההחזר על ההשקעה ל32 x 128, 10,000 חזרות) ולהתחיל את רכישת רקע המצלמה אות.
- סדרת מסגרת היבוא שנרכש לתכנית עיבוד נתונים. לחשב ולבדוק את העצמה הממוצעת בכל פיקסל על מנת לוודא שהרקע המצלמה הוא כ שטוח באזור הנבחר של השבב. בקצוץ מצב, להסיר הראשון וכמה קווים אופקיים שעבר (3 עד 10 בהתאם לגודל של ROI) עבור כל מסגרת, כי רקע המצלמה בדרך כלל מוטה בקווי הגבול.
- צור היסטוגרמה של הערכים (גם מוגדרת דיגיטלי רמה, DL) בתמונות שנרכשו ערימה (באמצעות 'היסטוריה' הפקודה בMatlab) ואת העלילה הלוגריתםשל תוצאת תדר (באמצעות פקודת semilogy בMatlab). דוגמא להפצת DL לרקע מצלמה מוצגת באיור 2.
הערה: אם המצלמה עובדת היטב, העלילה תציג שיא כ גאוס (פרופיל פרבולית ביומן בקנה מידה) המייצג את התפלגות ערכים הקשורים לאפס פוטון אחרי דעיכה מעריכית (קו עם שיפוע שלילי ביומן בקנה המידה ) שמייצג את התפלגות ערכים הקשורים לפוטון 1 (איור 2). בפרט, במרכז ואת השונות של פונקצית גאוס מייצגים את המצלמה לקזז וטעייה, בהתאמה, ואילו קבוע הדעיכה של חלק מעריכי מייצג הערכה של DL שהוקצה על ידי המצלמה לכל פוטון יחיד. בMatlab להשתמש בסעיף "CalibrateCamera" של סקריפט בתמיכה בחומרים. - חזור על הפעולה עבור כל המצלמה שנבחרה EMGain ורווח.
.2 תוויתהכנת תא
- כדי להכין את יפוזומים הנדרשים לשילוב שומנים 36, לפזר בנפרד 1 מ"ג של DOPE (1,2-dioleoyl- sn -glycero-3-phosphoethanolamine), 1 מ"ג של DOTAP (1,2-dioleoyl-3-trimethylammonium-פרופאן), ו1 מ"ג של PPE-ATTO488 ב 1 מ"ל של כלורופורם. מערבבים יחד 0.5 מ"ל של תמיסת DOPE, 0.5 מ"ל של תמיסת DOTAP, ו25 μl של פתרון PPE-ATTO488 ויבש תחת ואקום ל24 שעות. הוסף 0.5 מ"ל של HEPES חיץ 20 מ"מ, מערבולת במשך 15 דקות וsonicate ל15 דקות ב 40 ° C.
- כדי להכין את התא, לשטוף 3 פעמים עם PBS צלחת P100 של המחוברות CHO-K1 (סינית שחלות), להוסיף 1 מ"ל של טריפסין וחנות בחממה במשך 5 דקות. להשעות תאים מנותקים הוספת 9 מ"ל של מדיום DMEM / F12 בתוספת 10% מFBS וזרע 150 μl של פתרון תא בצלחת פטרי המכילה 800 μl של אותו המדיום.
- חנות בחממה ל24 שעות ב37 מעלות צלזיוס, 5% CO 2. עבור התאגדות שומנים, להחליף תאים בינוניים עם 500μl של מדיום סרום ללא; לאחר 30 דקות, להוסיף 2 μl של פתרון ליפוזומים; לאחר 15 לשטוף דקות עם PSB ולהוסיף בינוני DMEM / F12 חדש להדמיה.
- לTransfection, תאי transfect על פי פרוטוקול Lipofectamine (הוראות יצרן) באמצעות פלסמיד TFR-GFP וחנות hr 24 בחממה לפני ההדמיה.
.3 Data Acquisition
- הכנת תוכנית ההתקנה
- כדי טרמוסטט מיקרוסקופ, 24 שעות לפני הניסוי להפעיל את החממה.
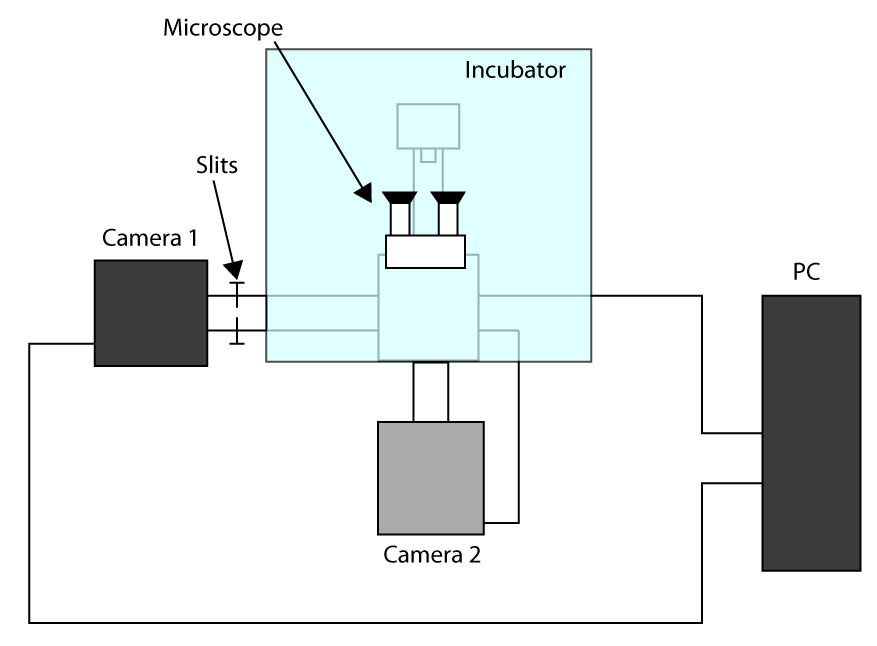
- על מנת להחיל את זמן רכישת השגה המהירה ביותר, עבודה בחיישן קצוץ מצב (ראה מבוא) ולהשתמש במצלמה ראשונה להדמיה (מצלמה 1) ומצלמה שנייה כדי לבחור את התא (המצלמה 2). ערכה של הגדרת התצורה מוצגת במשלים איור S1. ואז, כדי ליישר את תור שתי מצלמות על המיקרוסקופ ולחכות למצלמות כדי להתקרר.
- להגדיר בשני מצלמות הפרמטרים להדמית אור מועברת (כלומר
- שים את הדגימות בבעל ולהתמקד באמצעות עינית, לשלוח האור למצלמה 1 ודחף בעדינות את החריצים המאפשרים אור היחיד בROI המשמש להדמית תא (כאן ROI 32 x 32 פיקסלים).
- הזז תא באזור נבחר ולשלוח האור למצלמה 2, ולאחר מכן לצייר את ההחזר על ההשקעה בתוכנה ששולטות במצלמה 2 כדי שתהיה לי התייחסות.
- הדמיה (איור 3 א)
- קודם כל, ליישר את לייזר TIRF בהתאם לנוהל של ההתקנה שלך. בהגדרה שלנו, לבחור את "הגדרת TIRF 'ולהתחיל את הליך היישור האוטומטי. כאשר הלייזר מיושר להגדיר 70 ננומטר של עומק חדירה (כ -70 °).
- להגדיר זמן חשיפה ל70 אלפית שניים וEMGain 100 משני המצלמה 1 והמצלמה 2; לאחר מכן, בחר תא באמצעות מצלמה 1, ולאחר מכן לשלוח את האור במצלמה 2 ובאופן מדויק להתמקד קרום התא. הגדר את החשיפה המינימלית על caMera 2, 1,000 EMGain, חיישן קצוץ מצב, 10 5 חזרות ושמירה אוטומטית מוגדרת כמתאימות קבצים (מערכת תחבורה תמונה גמישה, פורמט שיכול להיות בקלות הצליח).
- הפעל את הרכישה כדי להקליט את סדרת התמונה. שחרר את רווח והמצב הקצוץ, כדי לאפשר ייצוב טמפרטורה לפני הרכישה סלולארי חדש, ולאחר מכן לחזור על שני השלבים האחרונים במטרה לרכוש 8-10 תאים.
.4 חישוב הממוצע כיכר העקירה מהדמיה (i MSD)
הערה: הפרוטוקול הבא יכול להיות מיושם ישירות לנתונים גולמיים. באותו הזמן, כל הפרוטוקול תקף לנתונים רכישות מדומה הן בMatlab ובSimFCS. ניתן למצוא הקישור להדרכות המתאימות בסעיף 'חומרי'.
- חישוב על ידי Matlab
- לייבא את הסדרה נרכשה לMatlab באמצעות ImportImageSeries תסריט. לחשב את העצמה הממוצעת של כל תמונה בזמן באמצעות שיתוףmmand אומר על 2 הממדים הראשונים ולהשתמש בעלילה כדי לראות את הווקטור וכתוצאה מכך.
- אם יותר מ10% מphotobleaching הוא הווה, לבטל את הסדרה או להסיר את חלקו הראשון שלהם. אם הוא נמוך, לנסות ולתקן את ההשפעה על תפקוד המתאם על ידי הפחתה לכל תמונה בעוצמה הממוצעת שלה, כפי שמוצג לפני 37.
- לחשב את העצמה הממוצעת של כל פיקסל באמצעות ממוצע בממד השלישי ותראה את תוצאת תמונה.
הערה: תשומת לב מיוחדת הנדרשת על מנת להימנע ממתאמי artifactual. למעשה, כפי שהוצג קודם לטכניקות דומות 38, גבולות תא, כמו גם משלפוחית המוקד יכול להציג את המתאם חזק. אם הבדיקה של התמונה הממוצעת מגלה גבולות תא או החוצה של שלפוחית מוקד, לנסות להוציא את האזור המעורב בדרך אחרת לבטל את הרכישה. כדי לתקן את ההשפעה של מבנים נייחים זה להחסיר את עוצמת הזמנית הממוצעת מכל פיקסל 39. - לחשב tהוא מתאם spatiotemporal (G (ξ, χ, τ)) על ידי שימוש בפונקציה CalculateSTICScorrfunc. הסר G (ξ, χ, 0) כי המתאם בשל רעש הירייה במשטר התאורה נמוך שולט G (0,0,0); המתאם בשל הגלאי שולט G (± 1,0,0), ותנועת חלקיקים בזמן החשיפה יכול לעוות G (ξ, χ, τ) לτ = 0 על ידי הגדלת המותניים נמדדו (אפקט זה נעלם לτ > 0) 34.
- G הממוצע (ξ, χ, τ> 0), תוך שימוש בזמן bin לוגריתמים כדי להפחית את הרעש על ידי שימוש בפונקציה "LogBinStack" בתמיכה בחומר ולאחר מכן להתאים את G וכתוצאה מכך (ξ, χ, τ) "gaussfit" באמצעות הפונקציה של הכלים ICS-Matlab בחומרים להתאושש i MSD (העמודה השנייה של המערך וכתוצאה מכך).
- העלילה σ הושג המותניים (τ) 2 (i MSD) כפונקציה של זמן. אם הנתונים הם רועשים מדי, תנסה להגדיל את מספר acquמסגרות ired, להגדיל את כוח הלייזר, ממוצע יותר G (ξ, χ, τ) יחד.
- חישוב על ידי SimFCS
- פתח את הקבצים שנרכשו עם ImageJ באמצעות תוסף היבואן BioFormat ולשמור סדרה נרכשה כרצף Tiff.
- כלי פתוח SimFCS ובחר RICS ובחר קובץ> תמונות מרובות יבוא (S2 איור משלים).
- בחר Fit, להכניס את הפרמטרים לרכישה הנכונות ולסגור את החלון בכושר (S3 איור משלים).
- בחר תצוגה> עוצמה ממוצעת> CH1 ולוודא את נוכחותו של photobleaching (S4 איור משלים).
- אם יותר מ10% מphotobleaching קיים קלפים שנזרקו הסדרה או אם זה עומס אפשרי שוב את התמונה ברצף הסרת החלק הראשון של הסדרה.
- אם הלבנת זה בחר באפשרות כלים נמוך מ10%> אני MSD> הפרמטרים שנקבע, לבדוק 'השתמש בממוצע נע ", שנקבע בפנל את ההחזר על ההשקעה בעזבהחום אדמדם של מסגרת לתשומת הלב משלם הממוצע נע שזמן הכתב הוא גבוה יותר מאשר בפעם דיפוזיה האופיינית (לחלקיקים הנעים בשניות 1 מיקרומטר 2 -1 זמן של 10 שניות הוא ממוצע נע טוב)
- בחרו כלים> iMSD> חישוב i MSD (איור משלים S5) ובכושר ולייצא את i MSD מהפנקס (איור משלים S6).
.5 חישוב חוק דיפוזיה מi MSD
- להתאים כמה נקודות הראשונות להסיק ליירט (σ 0 2) (5 נקודות הן בדרך כלל מספיק, אבל יכולות להיות מצוידות יותר נקודות אם הם מראים התנהגות ליניארית) ולהשוות את הערך הזה עם כוחות הביטחון הפלסטיניים שנמדדו בעבר 2. אם הם דומים, את הדינמיקה של fluorophores המבודד הם עוקבים אחריו. לעומת זאת, אם σ 0 2 >> כוחות הביטחון הפלסטיניים 2 לנסות לרכוש יותר מהר כדי להבטיח שדינמיקה לא מוסתר נמצאות 34.
- לחשב את diffusivity לכאורה (האפליקציה D) ועקירה הממוצעת (R) באמצעות משוואות 3 ו -4 (ראה מבוא).
- האפליקציה D עלילה כפונקציה של R כדי להשיג חוק דיפוזיה השוואה עם מה שנמדד עם וריאציה מקום FCS 12 (איור 3D) המבוססת.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
על מנת לכייל את המותניים אינסטרומנטלי, את דמותו של ננו חרוז ניאון בודד יכולה להיות מדד כמתואר בשלב פרוטוקול 1.1. תמונת ניאון אופיינית של חרוזים אלה מוצגת באיור 1. ההולם של חלוקת עוצמה על ידי פונקצית גאוס 2D נותן בחזרה שאריות טובות ומאפשר מדידת המותניים אינסטרומנטלי ב270 ננומטר. ערך זה הוא בהסכם טוב עם המגבלה העקיפה הצפויה המוערכת על ידי משוואת ריילי. כיול זה אינו הכרחי למדידת הדינמיקה של חלקיקים, אבל זה נדרש כדי למדוד את גודל חלקיקים הנראית לעין.
חלוקת תדר אופיינית של רקע מצלמה מוצגת באיור 2. השיא בכ -180 DL נובע מתגובת המצלמה לשום פוטון, והיא מייצגת את התרומה של אנלוגי דיגיטלי ממיר (AD). תרומה זו יכולה להיות מקורב כמו הפצת גאוס להעריך את הקיזוז ואת השונותהוצג על ידי הקלטת האות. מעל 200 DL הפצת הרמה הדיגיטלית הופכת מעריכי (ליניארי בסולם לוגריתמים) ומייצגת את תגובת המצלמה הממוצעת לפוטון יחיד. התאמת חלק זה עם התפלגות מעריכית מאפשרת מדידות של DL הממוצע שהוקצה לכל פוטון יחיד. גבוה יותר הוא היחס בין DL הממוצע שהוקצה לכל פוטון ושגיאת ממיר AD, נמוך יותר יהיה הרעש בפונקצית המתאם מחושבת. יתר על כן, תגובת הפוטון היחידה הממוצעת מאפשרת ההערכה של הטווח הדינמי המצלמה.
תרשים של ההליך ניסיוני המלא מסוכם באיור 3 ותמונה של הכנסת Atto488-PPE לתוך הקרום מיוצג באיור 4 א. תמונת TIRF נציג של קרום הבסיס של תאי CHO שכותרתו עם Atto488-PPE מוצגת באיור 4. כמה כתמים בהירים יכולים להיות נוכחים מחוץ לתא בשל liposomes נערם על הזכוכית. הם יכולים להיות מושלכים על ידי בחירת החזר על השקעה על חלק קרום בעיקר אחידה בקרינה (כלומר., קרום הפלזמה הסלולרי). כצפוי חוק דיפוזיה נמדד (איור 4C) לשומנים בדם זה הוא שטוח, המצביע על דיפוזיה לרוב ללא תשלום, כפי שמוצג בעבר על ידי מדידות STED-FCS 30,35. ראוי להזכיר כי כל ערכי העקירה המוצגים הם מתחת לגבול ההשתברות, המציין את יכולתה של גישה זו לבבירור סופר לפתור התקות מולקולריות ממוצעת נמוכות בהרבה ממגבלת העקיפה ועד לכמה עשרות ננומטרים.
Schematization של הכנסת דימר TFR-GFP לתוך הקרום מיוצג באיור 5 א. מחקרים רבים הראו כי זנב cytoplasmic של קולטן זה אינטראקציה עם שלד הקרום, אשר בתורו פועל כגדר ל12,40 ניידות קולט. תמונת TIRF נציג של תא CHO להביע TFR-GFP היא presented באיור 5. תאי עוצמת הקרינה נמוכה יש להעדיף, כקרום הוא קרוב יותר למצב היליד וההסתברות של חפצים הקשורים לביטוי היתר ממוזער. בנוסף, יש להימנע מחלק מהתא המרכזי, כמו את ההשפעות של הקרינה מחוץ לפוקוס (מציטופלסמה, למשל) עשוי להיות נוכח. כצפוי חוק דיפוזיה נמדד (איור 5 ג) לTFR-GFP מציג התנהגות שטוחה ראשונה מתחת 100 ננומטר, עם יישום D ממוצע של כ 0.7 מיקרומטר 2 שניות -1, ואחריו ירידה מהירה כתוצאה מכך בdiffusivity לכאורה עד 0.2 מיקרומטר 2 שניות -1 (הערך שנמדד בדרך כלל על ידי 12 FCS עקיפה מוגבל). תוצאה זו מראה כי הגישה שלנו בקלות יכולה למדוד את התזוזה הממוצעת של חלבוני GFP שכותרתו עם רזולוציה של כמה עשרות ננומטרים. יתר על כן בקנה מידה המרחבי שבו האפליקציה D מתחילה ירידת סטים האופייניתבקנה מידה המרחבי של כליאה חלקית חלבון על ידי שלד קרום בסביבות 120 ננומטר, בקנה אחד עם הערכות קודמות 6.

כיול .1 איור של פונקצית התפשטות נקודה. תמונת Pseudocolor של אגרגטים חרוז וחרוזים מבודדים (). עלילת 3D (B) של פרופיל העצמה של חרוז בודד מציגה פרופיל גאוס מוגדר היטב. (C) Fit של חלוקת העצמה על ידי פונקצית גאוס (פנל עליון) עם השאריות המקבילות (פנל תחתון). ההסכם הטוב בין ההתפלגות המצויד ופרופיל העצמה נמדד גם הוכחה לכך שכוחות הביטחון הפלסטיני סייעו יכולים להיות מקורבים על ידי פונקצית גאוס. אנא לחץ כאן כדי VI EW גרסה גדולה יותר של דמות זו.

כיול איור 2 של תגובת מצלמה לפוטונים בודדים. איור מציג את ההפצה דיגיטלית ברמה (DL) על רקע מצלמה בהחזר על השקעת 32 x 128, חשיפת 0.5 אלפיות שניים, בחיישן קצוץ מצב. השיא בכ -180 DL מייצג את תגובת המצלמה לשום פוטונים. במיוחד, הוא מייצג את תרומתו של ממיר האנלוגי הדיגיטלי (AD) ויכול להיות מקורב על ידי פונקצית גאוס להעריך את קיזוז ואת השונות שהוצגו על ידי הקלטת האות. מעל 200 DL חלוקת הרמות דיגיטליות הופכת מעריכי ומייצגת את תגובת המצלמה הממוצעת לפוטון יחיד. המדידה של פרמטרים אלה מאפשרת לאמוד את הצפיפות של פוטונים שנרשמו במהלך הרכישה.s / ftp_upload / 51994 51994fig2highres.jpg "target =" / _blank "> לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

איור 3 Schematization של השיטה. הדמיה שדה רחבה על ידי המצלמה EMCCD () מוחלת על להגיע לפתרון תת אלפית שנייה, ואילו microcopy TIRF מנוצל על מנת לספק חתך אופטי מדויק של קרום הפלזמה. (B) הערימה של תמונות שנוצרה autocorrelated כדי לחשב את מרחבי הממוצע פונקצית קורלציה -temporal. פונקצית קשר זה מקורב היטב על ידי פונקצית גאוס (ראה מבוא) והוא מתפשט החוצה בזמן על פי התקות חלקיקים. (C) וכך, על מנת לכמת את הפצה של פונקצית קורלציה בשל עקירה מולקולרית, התאמה עם גאוס פונקצית איאן מתבצעת. זה מאפשר מדידה "חוק דיפוזיה 'המולקולרי ישירות מהדמיה, בצורה של diffusivity לכאורה לעומת עלילת עקירה ממוצעת. (ד') הודות לעלילה זו, מצבי דיפוזיה מולקולריים יכולים להיות מזוהה באופן ישיר ללא צורך במודל או הנחות פרשניים על ארגון המרחבי של הקרום. למעשה, מולקולות לשדר באופן חופשי תציג diffusivity לכאורה קבוע כמו הניידות שלהם אינה תלויות בקנה המידה המרחבי של המדידה. לעומת זאת, מולקולות מוגבלות באופן חלקי תציג diffusivity לכאורה די קבוע עבור התקות קטנות יותר מגודל כליאה, אז diffusivity יורד לקני מידה מרחביים גדולים יותר מגודל כליאה. כך, המראה של ירידה בdiffusivity לכאורה יכול להתפרש כטביעת אצבע של כליאה חולפת, ואילו בקנה מידה המרחבי הקשורות ניתן להשתמש כדי להעריך את ההארכה המרחבי של הכליאה. קבצים / ftp_upload / 51994 51994fig3highres.jpg "target =" .jove.com / / _blank "> לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

איור 4 חוק ATTO488-PPE דיפוזיה בקרום תא חי ייצוג סכמטי של הכנסת ATTO488-PPE בקרום תא (א) (ב) תמונת TIRF של קרום בסיס CHO שכותרתו עם ATTO488-PPE:.. ROI (תיבה אדומה) נבחר בחלק בעיקר אחיד של התא, הימנעות גבול תא וכתמי ניאון מאוד. (C) דיפוזיה החוק נמדד בהחזר על ההשקעה שנבחרה מציג התנהגות שטוחה המאשרת מודל דיפוזיה חופשי לרכיב זה. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של נתון זה.

איור חוק .5 TFR-GFP דיפוזיה בקרום תא חי () ייצוג סכמטי של הכנסת TFR-GFP בקרום תא:. זנב cytoplasmic של הקולטן אינטראקציה עם שלד הקרום, הפועל כגדר לניידות קולט (B. ) תמונת TIRF של CHO TFR-GFP להביע: החזר על השקעה נבחרה והעדיפה תאים לבטא נמוכים, כדי למנוע חפצים בשל ביטוי יתר (C) חוק דיפוזיה של TFR (נקודות שחורות), שלא כמו PPE (קו אפור, נלקח מהאיור 4),. תערוכות ההתנהגות הטיפוסית של דיפוזיה מוגבלת באופן חלקי שבו חלק שטוח ראשון ואחריו ירידה באפליקצית D. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של fi זהתרשים.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
מעקב אחר חלקיקים בודדים (SPT) מייצג את אחת מהאסטרטגיות הנפוצות ביותר ללמוד דינמיקה מולקולרית ויש לו את היתרון הגדול של מדידת מסלולי חלקיקים. זה בתורו מאפשר חיטוט ההתנהגות של כמה אפילו חלקיקים שכותרתו במערכת מורכבת. עם זאת, כדי להגיע ליתרון זה SPT בדרך כלל זקוק לצפיפות נמוכה של החללית ותוויות בהירות מאוד. במיוחד, כדי לקבל רזולוציה גבוהה של זמן (טווח μsec) בדיקה אורגנית בדרך כלל נדרשת (לדוגמא, נקודות קוונטיות או חלקיקי מתכת): במקרה זה הליך מורכב של ייצור, תיוג ולהכניסו למערכת יש צורך. בהשוואה לSPT השיטה הנוכחית מראה כמה יתרונות מרכזיים. קודם כל, גישה זו ניתן להשתמש בנטייה עם חלבוני ניאון. לפיכך, בהשוואה לSPT, החלטה זמנית גבוהה יותר מושגת (עליה התווית) הודות לכמות נמוכה יותר של פוטונים הנדרשת 34. יותר בפירוט, נכס זה מאפשר דוחף Resol הזמניution להלן 10 - 3 שניות גם בעת שימוש בחלבוני ניאון encodable, ולוח זמנים זה נותן גישה בלעדית לדינמיקה בקנה מידה ננומטרי של מרכיבי קרום. לבסוף, ראוי לציין כי חוקי דיפוזיה מולקולריים מתוארים על ידי ניתוח פונקצית מתאם החלל במשרה המלאה, ללא צורך לעקוב אחר כל מולקולה.
ההשוואה עם FCS מבוסס STED היא גם מעניינת. במדידה STED-FCS זמן המעבר הממוצע של מולקולות להפחתת כמויות תצפית נמדד על ידי המתאם הזמני של אותות הקרינה. זה מאפשר קבלת מדידה מקומית של הדינמיקה המולקולרית גם מתחת לגבול ההשתברות. בגישה הציגה את חוק דיפוזיה נמדד כממוצע של כל החלקיקים שנעים בהחזר על ההשקעה שנבחרה, שנצפה על ידי אמצעי סטנדרטי, מוגבל עקיפה, תצפית הנפח. עם זאת, דיווח על תוצאות מראות כי שיטה זו אינה מוגבלת על ידי עקיפה, אלא רק על ידי ההחלטה זמנית דואר זמין. למעשה, למרות שרכישה עקיפה מוגבל משמשת כדי לזהות תנודות (באנלוגיה למה שנעשה בטכניקות אחרות ברזולוציה סופר, כגון PALM וSTORM), התקות מולקולריות היטב מתחת לגבול ההשתברות יכולה להיות (באופן ישיר) מחושב, כפי שכבר הוכיח באמצעות STICS למדוד מולקולרי זורם 32. יתר על כן, בניגוד STED-FCS, גישה זו יכולה להיות מיושמת בקלות למגוון רחב של מערכים מיקרוסקופי מסחריים וקיימים, כגון מיקרוסקופי סריקת סריקה או מיקרוסקופים מבוססי מצלמה שדה רחב. זה ראוי לציון כי מדידות STED-FCS של חוקי דיפוזיה המולקולריים דורשות כיול fluorophore תלוי בגודל של המותניים אינסטרומנטלי בקפדנות. מנגד, המדידה שהוצגה כאן אינו דורשת כיול מערכת (נחוץ רק להערכת של גודל חלקיקים).
ההחלטה בפועל במדידה של התקות חלקיקים על ידי דה השיטה המוצגתpends על איך במדויק אנחנו יכולים למדוד את פונקציית המתאם. כתוצאה מכך, היא אינה מוגבלת מטבעו על ידי עקיפה, באנלוגיה למקרה SPT בי הרזולוציה תלויה איך במדויק "התמונה" החלקיקים נמדדה. כדי למדוד קורלציה משמעותית בדקות פחות מ 1 לניסויים המוצעים, כמה פוטונים (בדרך כלל מתחת 10 פוטונים) לכל חלקיק בכל מסגרת הם מספיק. למעשה, התרומה של כל החלקיקים שנצפו היא בממוצע יחד כאשר פונקצית המתאם מחושבת, גם אם חלקיקים אינם מבודדים. מאפיין זה הוא מהותי של שיטות מתאם תנודות ומאפשר שימוש בתוויות עמומות וצפופות, כגון חלבוני ניאון transfected בתאים חיים.
עם זאת בחשבון שהוא מופיע ברור שעקירת המדידה המינימלית תלויה בdiffusivity של החלקיקים ועל הרזולוציה של התקנת ההדמיה הזמן. כדוגמא, בבקשה לשקול דיפוזיה של מולקולות על קרום התא,בי diffusivity המרבי שנמדד לחלבונים או שומנים הוא סביב 5 מיקרומטר 2 -1 שניות. בתנאים אלה, אנו זקוקים לרזולוציה של כ 10 -4 שניות זמן כדי לתפוס תזוזה ממוצעת של 50 ננומטר. רזולוציה זמן זו יכולה להיות מושגת על ידי מיקרוסקופי סריקה מהירים לאורך קווים בודדים או על ידי מצלמה EMCCD מהירה, שבו ברזולוציה הזמן עולה בקנה אחד לזמן החשיפה, כהראתה כאן.
דרישה הכרחית נוספת לשיטה זו כדי לתאר במדויק את הדינמיקה מולקולרית היא דגימה המרחבי נכונה. למעשה, על מנת להתאים את פונקצית קורלציה אנחנו צריכים דגימה מרחבית (גודל פיקסל) נמוכה מהמותנים של כוחות הביטחון הפלסטיניים סייעו. ברוב המיקרוסקופים המסחריים (שדה confocal או רחב), המותניים וכוחות הביטחון הפלשתינים משתרעים מ200 ננומטר 500 ננומטר (תלוי בעיקר בצמצם המספרי של המטרה שנבחרה ועל אורך הגל בשימוש) וניתן למדוד בקלות על ידי ננו באמצעות ניסוי כיול חרוזי ניאון בגודל. Tהוס, גודל פיקסל של 70-150 ננומטר (3 פעמים נמוכות יותר מאשר המותניים אינסטרומנטלי) יכול להיות מספיק. עם זאת, גודל פיקסל יכול להיות מותאם למערכת תחת מחקר לוקחת בחשבון כלל פשוט: הקטן את גודל פיקסל, גבוה יותר הדיוק בתיאור של פונקצית המתאם. יתר על כן, הגודל המינימאלי של התמונה שיש רכש צריך להיות לפחות 3 פעמים-גדול יותר מהעקירה המרבית של ריבית (בתוספת המותניים אינסטרומנטלי). זה נדרש כדי להגיע להתכנסות טובה של האלגוריתם הראוי ודגימה משמעותית מבחינה סטטיסטית של התקות מולקולריות. כדוגמא, ללמוד התקות מולקולריות ממוצעת קטנות יותר מכמה מאות ננומטרים (לדוגמא, 200 ננומטר) גודל תמונה של מיקרונים אחדים היא מספיק. יתר על כן, המספר הכולל של פיקסלים (לקיחה קבועה גודל פיקסל) משפיע על איכות תפקוד המתאם. למעשה, תמונה מוגדלת מאפשרת בממוצע יותר מידע בפונקציית המתאם, גם אם על חשבון של resolut הזמןיון. בנוגע למערכת מבוססת המצלמה המשמשת כאן, שים לב שהגודל הפיזי של פיקסל על השבב קבוע. כתוצאה מכך, להקטין את גודל פיקסל מוריד את האות בכל פיקסל (שתלויה בכיכר בגודל פיקסל), מקטין את שדה הראייה, ודורש כוח הגדלה גבוה יותר. מצד השני, במערכת סריקה, שבו אזור התצפית קבועה, להקטין את גודל פיקסל גורם בדרך כלל למספר הפיקסלים גדל על חשבון הרזולוציה הזמן.
פרטי מעטים על גלאים המשמשים יש לדון. שלא כמו גלאי פוטון יחיד, מערכות EMCCD למדוד עוצמה ממוצעת (ברמה דיגיטלית, DL) שאינה עומד ביחס ישר לאור שנאסף בשל נוכחותם של קיזוז. גם אם זה בקיזוז הוא נמוך בהשוואה לטווח הדינמי של המצלמה (כמה מאה לעומת 2 16 ב16 קריאה קצת) וזניח בניסויים שבם פוטונים רבים שנאספו, יש לו שיש לקחת בחשבון ללהשיג נורמליזציה נכונה של פונקצית המתאם. כמו כן, קיזוז יכול לשמש כנקודת התייחסות בתנאי תאורה נמוכה על מנת לזהות את כמות האות שנאסף. יתר על כן, על מנת להעריך את הכמות הממוצעת של פוטונים שנאספו במהלך הרכישה, הרמה הדיגיטלית הממוצעת הקשורים לכל פוטון שנאסף יש כדי להימדד. כמות זו יכולה להאסף על ידי חשיפת המצלמה לעוצמת אור נמוכה מאוד (למשל, האור ברקע בחדר); למעשה, במקרה זה, סביר להניח שפוטונים רק בודדים מגיעים למצלמה, כלומר את עוצמת נמדדה יכולה להיות קשורה לאפס או אחד פוטון היחיד.
לבסוף, תן לנו תגובה על איך כמה מערכות רכישה חלופית (כלומר, הגדרות מיקרוסקופיה שונות) יכולות לשמש לביצוע המדידות שהוצגו. קודם כל, 'W'factor במשוואה 2 (המייצג את autocorrelation של כוחות הביטחון הפלסטיניים אינסטרומנטלי) יכול להיות מותאם לtמערכת שהוא בפרט רכישה משמשת כדי להתאים את פונקצית המתאם הניסיונית. כפי שניתן לראות 34 בעבר, מקרה קל הוא הרכישה כהוא מיקרוסקופ סריקת לייזר כאשר מהירות הסריקה גבוהה משמעותי מדינמיקה של חלקיקים. במקרה כזה, למעשה, התנועה של חלקיקים בזמן הרכישה (כלומר, קו זמן) יכולה להיחשב זניחה ופונקצית המתאם מקורבת היטב על ידי פונקצית גאוס. בהקשר של טכנולוגיות הדמיה המתעוררות, גישה מעניינת מבוססת על האפשרות לייצר יריעות דקות מאוד אור (1-2 מיקרומטר) באמצעות המדגם 41. גיליון האור מאפשר תאורה סלקטיבית של מטוס בודד (מיקרוסקופיה תאורת מטוס בודדת, SPIM) במדגם ו, בשילוב עם מערכת מבוססת מצלמה רכישה, חתך אופטי מהיר ב3D 42. בגלל מאפיינים אלה, SPIM כבר מצומדת בהצלחה עם FCS 43 ויכול לייצג הוואליכלי ד להאריך את הניתוח שהוצג לסביבות 3D, כגון ציטופלסמה או הגרעין של תאי חיים.
בסיכום, מנקודת המבט ניסויית גישה זו דורשת גישה רק למיקרוסקופ מצויד במודול רכישה מהיר. החלבון של עניין יכול להיות מתויג עם כל חלבון פלואורסצנטי או fluorophore האורגני, ובכך מאפשר גם להדמיה ססגוניות. בהקשר זה, אנו רואים את האפשרות להשתמש צולב ניתוח אני MSD כדי לבחור תת אוכלוסיות של מולקולות ולחשוף אינטראקציות ושיתוף דיפוזיה בקרום תא חי. לבסוף, אנו מאמינים כי גישה זו עשויה לייצג כלי רב עוצמה כדי לחקור חלבונים ו / או שומנים עוברים מחיצות דינמיות בתוך nanodomains על קרום הפלזמה. במקרה זה, בגודל ובחיים מאוד משתנה מnanodomains להציג את רמה נוספת של מורכבות בנתונים אמיתיים שהיה דורשות יישומים מתודולוגיים נוספים כוללים הדמיה 2 צבעים, מקומיניתוח (למשל, מתאם 2D זוג) ו / או אנאיזוטרופיה הקרינה.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}