

Abstract
그것은 공간적 분포 및 지질과 단백질과 같은 막 성분의 움직임이 많은 세포 기능의 조절에 중요한 요인이라는 것을 분명하게 증가했다. 그러나, 빠른 역학과 관련된 작은 구조로, 매우 높은 시공간 해상도는 분자의 실제 동작을 잡을 필요합니다. 여기에서 우리는 높은 시공간 해상도의 살아있는 세포에 형광 표지 된 플라즈마 막 단백질과 지질의 역학을 공부 실험 프로토콜을 제시한다. 특히,이 방법은 각각의 분자를 추적 할 필요가 없지만, 그 막의 소정 영역에있는 모든 분자를 사용하는 인구의 동작을 계산한다. 시작점은 막에 소정 영역의 고속 촬상이다. 이후, 완전한 시공간적 자기 상관 함수는 예를 들어, 각각 2, 3, N 번 반복 시간 지연을 증가에서 획득 된 영상의 상관 관계를 계산한다. 이는 폭을 입증 할 수있다확산으로 인한 입자 운동의 함수로서 시간 지연을 증가에서 공간적 자기 상관 함수 증가 피크. 따라서, 자기 상관 함수의 일련의 피팅하는 것은 실제의 단백질이 여기에 평균 겉보기 변위 대 확산의 형태로 표시, 영상 (iMSD)에서 제곱 평균 변위를 추출 할 수있다. 이 나노 미터 정밀도로 단일 분자의 평균 역학의 양적보기를 얻을 수 있습니다. 표지 된 트랜스페린 수용체 (TFR) 및 ATTO488의 GFP 표지 된 변형을 사용하여 1 - 팔미 토일 -2 - 하이드 록시 SN -glycero-3-phosphoethanolamine (PPE)가 켜져 단백질 및 지질 확산 시공간 규제를 준수 할 수있다 마이크로 - 투 - 밀리 초의 시간 범위에서 μm의 막 크기의 영역.
Introduction
가수 Nicolson 의해 일본어 "유체 모자이크"모델부터 출발 세포 원형질막의 영상이 지속적으로 세포 골격과 지질 1,2 도메인의 새로운 역할을 포함하기 위해 지난 수십 년 동안 업데이트되었습니다.
첫번째 관찰은 막 단백질의 상당 부분이 3-5 움직입니다 (FRAP) 발표를 photobleaching에 후 형광 복구에 의해 얻어졌다. 이러한 선구적인 연구는 매우 유익하지만, FRAP의 설정의 상대적으로 가난한 공간 (마이크론)의 해상도와 시간 (초)으로 고통. 또한, 앙상블 평균 측정되는, FRAP는 단일 분자의 행동에 관한 정보를주는 없다.
이러한 맥락에서, 특히 가능성 (시 확산 처리 한 분자의 연구를 허용) 매우 밝은 태그와 단일 분자 라벨을 매우 성공적이었다. 특히, 밀어등 마이크로 척도, 쿠스미 크게 막 생리 6 액틴 기반 막 골격의 역할 인식에 기여 지질과 단백질 역학의 알 수없는 기능에 알. 얻은 액세스 단일 입자 추적 (SPT) 방식의 시간 해상도 , 7. 이러한 결과는 지질 단백질 및 액틴이 확산 기반 골격에 의해 조절되는 '피켓 울타리'모델을, 소위 생성. 그러나 여러 실험적인 문제 SPT에 의해 제공되는 정보의 양에 큰 권한을 부여하기 위해 해결되어야한다. 특히, 라벨 절차는 일반적으로 시스템에 표시된 종의 생산, 정제 및 도입 등 여러 단계로 구성되어있다. 또한, 라벨은 큰, 양자점 또는 금속 나노 입자와 같이 자주 서브 - 밀리 세컨드 타임 스케일 많은 경우에 회피 될 수없는 라벨 표적 분자의 가교에 도달해야한다. 마지막으로, 많은 궤적통계 학적 기준에 맞게 기록 될 필요와 병용 라벨의 저밀도는 추적을 허용하도록 요구된다.
SPT에 비해 이러한 단점을 많이 극복 형광 상관 분광학 (FCS)는, 분자 역학을 연구하는 매우 유망한 접근 방식을 나타냅니다. 사실, FCS는 일시적으로 형질 세포에 형광 단백질 태그 분자의 역학을 연구 할 수 있도록, 희미하고 조밀 한 레이블 또한 잘 작동합니다. 또한, 제한된 시간에 높은 통계에 도달한다. 마지막으로, 라벨의 "높은"밀도에도 불구하고 FCS는 단일 분자 정보를 제공합니다. 덕분에 모든 속성, FCS는 매우 간단 접근 방식을 나타냅니다 광범위 모델 막에서 라이브 세포 8-10에서 모두 지질과 단백질 역학을 연구하기 위해 적용되었습니다. 많은 다른 접근법은 분자 확산의 상세를 공개하는 FCS의 능력을 증가시키기 위해 제안되었다. 예를 들면, SH이었다자신의 다른 크기의 관찰 영역에 FCS를 수행하여 하나의 분자 운동 (11, 12)의 "FCS 확산 법"계몽 숨겨진 기능을 정의 할 수 있습니다. 크기가 변화되는 외에, 초점 영역도 13 중복 된 고속 카메라 (21, 22)와 선 (14-20) 또는 복합 따라 공간에서 움직였다. 이 '시공간'의 상관 관계에 접근하여, 여러 가지 막 구성 요소의 관련 생물학적 매개 변수를 정량적 모델 막 실제 생물들, 막 공간 조직에 따라서 항복 통찰력을 모두 설명했다.
그러나, 모든 FRAP 및 FCS 애플리케이션에서 설명한 바와 초점 영역의 크기를 극복 할 수없는 공간 해상도의 한계를 나타낸다. 몇몇 초 - 해상도 이미징 방법은 최근이 제한을 우회하기 위해 개발되어왔다. 일부는 같은 확률 광 재건 현미경 (STORM)로, 현지화 정밀도를 기반으로 <SUP> 23,24, photoactivation 현지화 현미경 (PALM) (25), 형광 PALM (FPALM) (26), 및 단일 입자 추적 PALM (sptPALM) 27 : 각 샷에 필요한 광자의 비교적 큰 양 그러나의 시간 해상도를 제한 적어도 몇 밀리 초에 이러한 방법, 따라서 생체 내에서의 적용을 방해.
대조적으로, 슈퍼 해상도 이미징을위한 유망한 대안은 공간적 자극 방출 고갈 방법 (STED 또는 가역 포화 광학 형광 전환 (RESOLFT)) (28, 29)과 형광 방출을 조절함으로써 열렸다. 이러한 접근 방식은 빠른 스캐닝 현미경 및 탐지 시스템을 사용하는 가능성 관찰 볼륨 성형 아니라 아래 회절 한계를 결합한다. 형광 변동 분석과 함께, STED 현미경 직접 지질 및 P의 나노 시공간 역학을 조사 할 수라이브 세포막 (30, 31)에서 roteins.
STED 기반 현미경 같은 물리량이 생균에 형광 표지 된 막 단백질 및 / 또는 지질의 역학 연구에 적합한 개질 시공간적 화상 상관 분광법 (STICS 32,33)의 방법에 의해 얻을 수있다 및 상용 현미경으로. 여기에 제시된 실험 프로토콜은 몇 가지 단계로 구성되어있다. 첫 번째는 막에 관심 영역의 고속 이미징을 필요로한다. 그 후, 화상의 생성 스택 평균 시공간 상관 함수를 계산하는데 사용된다. 상관 함수의 시리즈를 끼워 맞춤으로써, 분자 '확산 법칙은'명백한 확산 계수 (D 앱)의 형태로 영상에서 직접 획득 될 수있다 - - 평균 변위 플롯 대. 이 플롯은 매우 분자에 의해 탐구 환경에 따라 직접 실제 확산 모드를 인식 허용관심 지질 / 단백질.
이전 34와 같이 더 상세하게, 상기 취득 된 이미지 시리즈의 시공간적 자동 상관 함수가 매우 수집 된 이미지 시리즈에서 이동 분자 역학에 의존한다 (같은 추론이 라인 획득에 적용될 수 있음을주의하세요 공간에서 하나의 차원은) 생각입니다. 특히, 상관 함수를로 정의한다 :
 (1)
(1)
여기서  위치 (x)에서 측정 한 형광 강도를 나타내고, Y 및 시간 t에서,
위치 (x)에서 측정 한 형광 강도를 나타내고, Y 및 시간 t에서, ![]() 및
및 ![]() 나타내고 X의 거리각각 y 방향,
나타내고 X의 거리각각 y 방향, ![]() 타임 래그를 나타내고,
타임 래그를 나타내고, ![]() 평균을 나타냅니다. 이 함수는 다음과 같이 표현 될 수있다 :
평균을 나타냅니다. 이 함수는 다음과 같이 표현 될 수있다 :
 (2)
(2)
'N'은 관찰 영역에서 분자의 평균 수를 나타내고, ![]() 공간에서의 컨볼 루션 연산을 나타내고,
공간에서의 컨볼 루션 연산을 나타내고,  쓸모있는 허리의 자기 상관을 나타냅니다. 후자 인해 광 기록 / 셋업 단일 광자 에미 터가 공간 내에 확산되는 방법의 측정으로 해석 될 수있다 (소위 점 분포 함수, PSF, 유전자잘) 가우스 함수에 의해 근사 집회. 마지막으로,
쓸모있는 허리의 자기 상관을 나타냅니다. 후자 인해 광 기록 / 셋업 단일 광자 에미 터가 공간 내에 확산되는 방법의 측정으로 해석 될 수있다 (소위 점 분포 함수, PSF, 유전자잘) 가우스 함수에 의해 근사 집회. 마지막으로,  거리 입자를 찾을 확률을 나타낸다
거리 입자를 찾을 확률을 나타낸다 ![]() 및
및 ![]() 시간 지연 후에
시간 지연 후에 ![]() . 우리는 입자가 모든 방향으로 임의로 이동 순 플럭스가 존재하지되는 확산 역학을 고려하면,이 기능들은 일반적 차이가 움직이는 입자의 평균 제곱 변위 (MSD)로 식별 될 수 가우시안 함수로 근사 . 따라서, 상관 함수의 허리는 (로도 지칭
. 우리는 입자가 모든 방향으로 임의로 이동 순 플럭스가 존재하지되는 확산 역학을 고려하면,이 기능들은 일반적 차이가 움직이는 입자의 평균 제곱 변위 (MSD)로 식별 될 수 가우시안 함수로 근사 . 따라서, 상관 함수의 허리는 (로도 지칭 ![]() ), 입자 근골격계의 합 수단 허리로 정의 될 수 있고, 가우시안 피트에 의해 측정 될 수있다각 시간 지연에 대한 상관 함수의 팅. 측정 전 MSD는 움직이는 분자의 겉보기 확산을 계산하는데 사용될 수있다
), 입자 근골격계의 합 수단 허리로 정의 될 수 있고, 가우시안 피트에 의해 측정 될 수있다각 시간 지연에 대한 상관 함수의 팅. 측정 전 MSD는 움직이는 분자의 겉보기 확산을 계산하는데 사용될 수있다 ![]() 및 평균 변위
및 평균 변위 ![]() 로 :
로 :
 (3)
(3)
 (4)
(4)
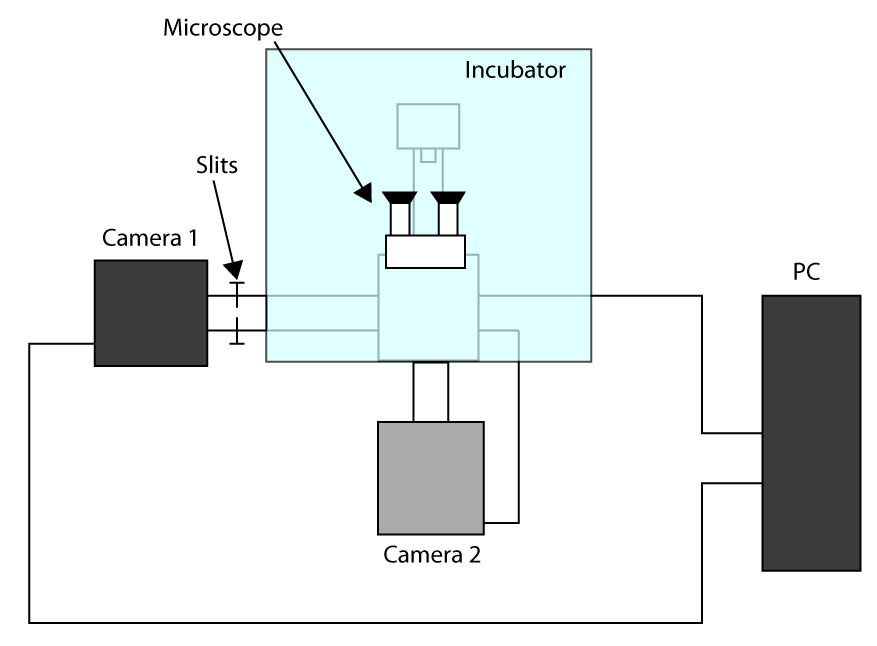
다음 섹션을 통해 독자를 안내 할 수 실험 구성에 대한 몇 가지 고려 사항. 선택적 상업용 TIR 형광을 이용하여, 우리는 전체 내부 반사 (TIR) 조명을 사용하는 살아있는 세포의 기저막에 형광체를 여기하기 위해 (TIRF) 현미경 (상세는 재료 부분에서 찾아 볼 수있다). 또한, 순서 번째 수집전자 형광 우리 고배율 대물 사용하며 (칩 16 μM의 화소의 물리적 크기) EMCCD 카메라 (100X NA 1.47, 높은 개구는 TIRF 조명 필요함). 100nm의 픽셀 크기에 도달하기 위해 우리는 1.6 부가 배율 렌즈를 적용. 후술하는 바와 같이, 1 밀리 초 이하의 시간 해상도는 100 나노 미터 이하로 적절히 빠른 막 지질의 역학을 설명하는 데 필요한 것. 이 시간 해상도에 도달하기 위해 우리는 카메라 (512 X 512)의 온 - 칩보다 작은이자 (ROI)의 영역을 선택해야한다. 이러한 방식으로, 카메라는 시간 해상도를 증가 선의 감소 된 수를 판독 할 것이다. 그러나,이 판독 정권 프레임 시간은 카메라 판독 칩에 노출 전하를 이동하는 데 필요한 512 X 512 픽셀 EMCCD 대 밀리 위해 통상 시간에 의해 제한 될 것이다. 이 한계를 이길하려면 새로운 기술은 와트, ROI 라인 만 대신 전체 프레임을 이동 할 수 있습니다(우리 EMCCD에서 자른 센서 모드라고 함)에 노출 칩 사이즈의 실질적인 감소 효과 제 i. 이 구성에 적용하려면, ROI 외부의 칩은 광로에 장착 슬릿 커플 적용되어야한다. 시간 해상도 다운 10-4 초로 설정이 덕분에 달성 될 수있다. '토론'섹션에 설명 된대로이 방법이 여러 가지 실험 설정과 결합 될 수 있다는 점 유의하시기 바랍니다.
방법은 데모 GFP- ATTO488 표지 모두 1 - 팔미 토일 -2 - 하이드 록시 -3 -glycero SN-phosphoethanolamine (ATTO488-PPE) 및 트랜스페린 수용체 GFP 표지 된 변형을 (사용함으로써, 생균에 제공 될 TFR). 이전에보고 된 바와 같이 30,35 ATTO488-PPE의 경우이 접근법은 성공적 주로 자유 확산을 나타내는 평균 변위의 함수로서 거의 일정한 D 애플리케이션을 복구 할 수있다. 대조적으로, TFR-GFP는 감소 D를 도시
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1 시스템 교정
- 포인트 확산 함수 (PSF) 교정
- 증류수 90 μL에 30 nm의 형광 비드 용액 (약 5 μM) 10 μl를 희석 한 후 20 분 동안 초음파 처리 용액. 사각형 (1cm X 1cm) 아가 로스 겔 (3 %)과 예금 겔의 상단에있는 용액 10 μL의 조각을 잘라. 2cm 페트리 접시의 바닥 유리에 젤의 조각을 뒤집고 유리에 드롭을 짠다.
- 이번 인수 설정을 켭니다 홀더에 샘플을 넣고, 카메라 노출과 EMgain을 설정 (100 밀리 초 및 1000 시스템에 따라 좋은 매개 변수이지만 최적화) 카메라가 식을 때까지 기다립니다.
- 100 밀리 초, 1000에 카메라 EMgain, 획득 모드로 설정 카메라 노출 설정 저장 전송, 100 반복 및 자동 프레임입니다.
- 겔의 테두리에 접안과 투과광 포커스를 사용하고, 겔의 중심부에 대물 이동 및 초점을 조절 STA레이저 정렬 절차 RT (LAS AF에 'TIRF 설정'을 선택하고 자동 정렬 절차를 따르십시오).
- , 참고로 (즉, 일반적으로 구슬 집계를 나타냅니다) 밝은 자리에 초점을 정확하게, 고립 된 하나의 관광 명소와 시야를 찾아 100 프레임을 획득하고 여러 단일 지점을 얻기 위해 단계 5-6 번 반복합니다.
- 데이터 처리 프로그램으로 취득한 일련 오기 및 시간 (도 1a)을 스택 평균 단일 절연 비드를 선택한다. 입자의 응집을 방지하기 위해 최소의 속성을 선택합니다주의하십시오.
- (매트랩 재료의 ICS-matlab에 도구에서) 명령 "gaussfit"을 사용하여 가우시안 함수를 선택한 강도 분포 (단일 구슬 프로파일의 예를 그림 1B에 제시되어있다)를 장착한다. 얻어진 잔류 물을 검사하여 적합도를 확인 (해당 잔차 장착 가우시안 프로파일의 예 난을 제시N 그림 1B).
- 카메라 보정
- 카메라를 켜고 카메라가 식을 때까지 기다리십시오. 카메라 설정 획득 설정 (즉, 사용 된 카메라 우리는 0.5 밀리 초, 1000에 카메라 EMgain, 자른 모드, 32 X 128, 10000 반복에 대한 투자 수익 (ROI) 크기로 획득 모드로 노출을 설정)와 카메라 배경의 획득을 시작 신호.
- 임포트 데이터 처리 프로그램에 프레임 시리즈를 인수. 계산하고 카메라 배경 칩의 선택 지역에서 약 평평한 지 확인하기 위해 각 픽셀의 평균 강도를 검사합니다. 카메라 배경 일반적 경계선 바이어스되기 때문에 자른 모드에서, 각각의 프레임에 대한 제 (ROI의 크기에 따라 3 내지 10) 최후의 수평 라인을 제거한다.
- 값의 히스토그램 만들기 (매트랩 명령 'HIST'를 사용) 스택 획득 된 영상에서 (또한 디지털 레벨, DL 정의) 및 로그를 플롯주파수 결과의 (매트랩 semilogy 명령을 사용하여). 카메라 배경 DL 분포의 예는 그림 2에 제시되어있다.
참고 : 카메라가 잘 작동하면, 플롯은 지수 붕괴 (로그 스케일에서 음의 기울기를 가진 라인으로 다음 제로 광자에 관련된 값의 분포를 나타내는 약 가우스 피크 (로그 스케일에서 포물선 프로파일)을 표시합니다 ) 한 광자 (그림 2)에 관련된 값의 분포를 나타냅니다. 지수 부분의 감쇠 상수가 각각의 단일 광자에 카메라에 의해 할당 된 DL의 추정을 나타내고, 특히, 중앙 및 가우스 함수의 분산은 각각 카메라와 오프셋 에러를 나타낸다. 매트랩 자료를 지원하는 스크립트의 섹션 "CalibrateCamera"를 사용합니다. - 선택된 모든 카메라 EMGain 및 이득에 대한 작업을 반복합니다.
2 레이블셀 준비
- (1,2 - dioleoyl- -glycero SN-3-phosphoethanolamine), 지질 혼입 36 필요한 리포좀을 제조 DOPE 별도로 1 mg의 용해, DOTAP (1,2-dioleoyl-3-트리메틸 프로판) 1 ㎎, 클로로포름 1 ㎖에 ATTO488 PPE-1 밀리그램. 24 시간 동안 진공에서 함께 0.5 도프의 수용액, DOTAP 용액 0.5 ml의 PPE-ATTO488 솔루션의 25 μL 및 건조를 섞는다. 40 ° C에서 15 분 동안 15 분, 초음파 처리에 대한 HEPES 버퍼 20 mm의 0.5 ml의, 소용돌이를 추가합니다.
- 셀을 준비 PBS로 합류 CHO-K1 (중국 햄스터 난소)의 P100 접시를 세 번 씻어 5 분 동안 인큐베이터에 트립신 및 저장 1 ㎖를 추가합니다. 10 % FBS 보충 된 DMEM / F12 배지 9 ml에 첨가 분리 된 세포를 일시 중단하고 동일한 배지 800 μl를 함유하는 페트리 접시에 세포 용액 150 μl를 시드.
- 37 ° C에서 24 시간 동안 인큐베이터에 보관 5 % CO 2. 지질 혼입 들어, 셀 (500)을 가진 매체를 대체무 혈청 배지 ㎕의; 30 분 후, 리포좀 용액 2 μl를 추가; 와 PSB 15 분 세척 한 후 영상에 대한 새로운 DMEM / F12 매체를 추가 할 수 있습니다.
- 형질의 경우, 형질 세포 이미징 전에 인큐베이터에서 TFR-GFP 플라스미드 및 저장 24 시간을 사용하여 리포 펙 타민 프로토콜 (제조사 지침)에 따라.
3 데이터 수집
- 설치 준비
- 현미경을 온도 조절하기 위해, 실험 전 24 시간은 인큐베이터를 켭니다.
- 가장 빠른 달성 획득 시간을 적용하기 위해, 자른 센서 모드에서 작업 (소개 참조) 셀 (카메라 2)를 선택 이미징 (카메라 1)와 두 번째 카메라에 대한 첫 번째 카메라를 사용합니다. 설치 구성의 방식은 보충 그림 S1 표시됩니다. 그런 다음, 현미경에있는 두 개의 카메라 회전을 맞추고 카메라가 식을 때까지 기다려야합니다.
- 두 카메라 투과광 이미징을위한 매개 변수를 설정합니다 (예 :
- 만 (여기에 32 × 32 픽셀의 ROI) 세포 이미징에 사용되는 투자 수익 (ROI)에 빛을 허용하는 슬릿을 밀어 부드럽게 홀더에 샘플을 넣고 접안 렌즈를 사용하여 초점 카메라 하나에 빛을 보내고.
- 선택한 영역에 셀을 이동하고 카메라 2에 빛을 보내, 다음 참조를하기 위해 카메라 2를 제어하는 소프트웨어의 ROI를 그립니다.
- 이미지 (그림 3A)
- 우선, 사용자의 설정의 방법에 따라 TIRF 레이저 정렬. 우리의 설정에서 'TIRF 설정'을 선택하고 자동 정렬 절차를 시작합니다. 레이저가 정렬되면 (약 70 °) 침투 깊이의 70 나노 미터를 설정합니다.
- 모두 카메라 1과 카메라 2에 100에 EMGain 70 밀리 초와 노출 시간을 설정하고, 다음, 카메라 (1)를 사용하여 셀을 선택한 후, 카메라 (2)에 광을 전송하고 정확하게 세포막 초점. 캘리포니아에 최소 노출을 설정메라 2, 1000 EMGain, 자른 센서 모드, 열 다섯 반복 설정하고 자동 저장 파일 (유연한 이미지 전송 시스템, 쉽게 관리 할 수있는 형식)을 맞는있다.
- 이미지 시리즈를 기록하는 인수를 시작합니다. 게인하고, 새로운 세포를 획득하기 전에 온도 안정화를 허용 8-10 세포를 얻기 위해 마지막 두 단계를 반복 자른 모드를 해제합니다.
영상에서 평균 제곱 변위의 4 계산 (I MSD)
NOTE : 다음 프로토콜 직접 미가공 데이터에 적용될 수있다. 동시에, 전체 프로토콜은 매트랩과 SimFCS 모두 시뮬레이션 데이터 수집에 유효하다. 해당 자습서에 대한 링크는 '재료'섹션에서 찾을 수 있습니다.
- matlab에 의해 계산
- ImportImageSeries 스크립트를 사용하여 matlab에 획득 한 시리즈를 가져옵니다. 공동 사용 시간에 각 이미지의 평균 강도를 계산mmand 제 2 차원을 의미하고, 생성 된 벡터를보고 플롯을 사용합니다.
- 광표백의 10 % 이상이 존재하는 경우, 일련의 폐기 또는 이들의 첫 번째 부분을 제거한다. 이 낮 으면 (37) 전에 도시 된 바와 같이, 각각의 이미지를 평균 세기를 감산함으로써 상관 함수에 대한 효과를 해결하기위한 시도.
- 셋째 차원 평균을 이용하여 각 화소의 평균 강도를 계산하고 화상 생성 참조.
참고 : 특별한주의를 인공적인 상관 관계를 방지하기 위해 필요합니다. 사실, 이전에 유사한 기술 (38), 셀 경계뿐만 아니라 포커스 소체에서와 같이 강한 상관을 도입 할 수있다. 평균 이미지의 검사 셀 테두리 또는 초점 소포에서 밝혀, 그렇지 않으면 인수를 폐기 관련된 지역을 제외하려고합니다. 이 부동 구조체의 효과를 해결하기위한 각 화소 (39)로부터 평균 순간 세기를 뺀다. - t를 계산기능 CalculateSTICScorrfunc을 사용하여 그는 시공간의 상관 관계 (G (ξ, χ, τ)). G는 (ξ, χ, 0)으로 인해 낮은 조명 정권의 샷 잡음에 상관 관계가 G를 지배하고 있기 때문에 (0,0,0)를 제거; 인해 검출기에 상관 측정 허리를 증가시킴으로써 τ = 0 G (ξ, χ, τ)을 변형 할 수 노출 시간 동안 G (± 1,0,0), 및 입자 운동을 지배한다 (이 효과는 τ 사라진다 > 0) 34.
- 다음 지지물에 "LogBinStack"기능을 사용하여 소음을 줄이기 위해 대수 시간 빈을 사용하여 평균 G (ξ, χ, τ> 0) (ξ, χ, τ) 기능을 이용하여 "gaussfit"의 결과 G 맞추기 소재의 ICS-매트랩 툴 I MSD (결과 배열의 두 번째 열)를 복구한다.
- 시간의 함수로서 얻어진 허리 σ (τ) 2 (I MSD)를 그린다. 데이터가 너무 시끄러운 경우 acqu의 수를 증가하려고IRED 프레임 함께 레이저 파워, 평균보다 G (ξ, χ, τ)을 증가시킨다.
- SimFCS에 의해 계산
- ImageJ에이 BioFormat 가져 오기 플러그인을 이용하여 획득 된 파일을 열고 티파니 순서로 인수 시리즈를 저장합니다.
- 열기 SimFCS 선택 RICS 도구 및 파일> 가져 오기 여러 개의 이미지 (보충 그림 S2).
- 맞는 창 (보충 그림 S3)을 맞추기를 선택 올바른 인수 매개 변수를 삽입하고 닫습니다.
- 디스플레이> 평균 강도> CH1을 선택하고 photobleaching에의 존재 (보충 그림 S4)를 확인합니다.
- 광표백의 10 % 이상이 본 폐기보기 시리즈 또는 가능하면 다시로드 시리즈의 첫 번째 부분을 제거하는 이미지 시퀀스 인 경우.
- 그것을 표백하는 10 %보다 낮은 도구를 선택하는 경우> 내가 MSD> 매개 변수 설정, 왼쪽에 ROI 패널에 설정 '사용은 이동 평균'확인상대 시간 확산 특성 시간보다 높다고 이동 평균 지불 주목 프레임의 엄버 (1 μM 2 초에서 움직이는 입자 -1 10 초의 시간이 좋은 이동 평균)
- 도구> iMSD> 아이 MSD (보충 그림 S5) 및 착용감을 계산하고 메모 패드에서 내가 MSD를 내보낼 (보충 그림 S6).
내가 MSD의 확산 법칙 (5) 계산
- 절편 (σ 0 2)를 추정하는 최초의 몇 가지 포인트를 장착 (5 점 정도면 충분하지만 더 많은 포인트가 선형 동작을 보여 주면 장착 할 수 있습니다) 및 이전에 측정 된 PSF이와이 값을 비교합니다. 그들은 비교할 경우, 격리 된 형광 물질의 역학을 따라야되고있다. 반면, σ 0 2 >> PSF이 시도는을 보장하기 위해 빠르게 취득하는 경우숨겨진 역학 34 존재한다.
- 겉보기 확산 계수 (D 응용 프로그램)와 식 (3)과 (4) (소개를 참조)를 사용하여 평균 변위 (R)을 계산합니다.
- FCS 12 (그림 3D)를 기반으로 현장의 변화로 측정되는 것과 비슷한 확산 법을 얻기 R의 함수로 플롯 D 응용 프로그램.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
프로토콜 단계 1.1에 기재된 바와 같이 수단 허리를 보정하기 위해, 단일 형광 나노 비드의 이미지를 측정 할 수있다. 이 구슬의 전형적인 형광 이미지를 그림 1에 제시되어있다. 2 차원 가우스 함수에 의해 강도 분포의 피팅 다시 좋은 잔차를 제공하고 270 nm에서 악기의 허리를 측정 할 수 있습니다. 이 값은 레일리 방정식에 의해 추정 된 예상되는 회절 한계와 잘 일치한다. 이 교정은 입자 동역학 측정을 위해 필요한 것은 아니지만 그것 겉보기 입자 크기를 측정하기 위해 필요하다.
카메라의 배경 전형적인 주파수 분포는 약 180 DL에서의 피크는 더 광자 카메라 응답에 기인한다.도 2에 제시하고, 아날로그 디지털 (AD) 변환기의 기여를 나타낸다. 이 기여는 오프셋 추정 가우스 분포 및 분산 같이 근사화 될 수있다신호 기록에 의해 도입. 200 DL 위의 디지털 레벨 분포 지수 (로그 스케일에서 선형)가되어 단일 광자의 평균 카메라의 응답을 나타냅니다. 지수 분포로이 부분을 피팅 각 단일 광자에 할당 된 평균 DL의 측정을 할 수 있습니다. 높은 각 광자 할당 평균 DL 및 AD 변환기 오류 사이의 비율이며, 낮은 계산 된 상관 함수에서 잡음 것이다. 또한, 평균 단일 광자 응답 다이내믹 레인지 카메라의 추정을 허용한다.
전체 실험 과정의 도면도 3에 요약되어 막에 Atto488-PPE의 삽입 그림은도 4a에 나타낸다. Atto488-PPE와 레이블 CHO 세포의 기저막의 대표적인 TIRF 화상은도 4b에 제시된다. 여러 밝은 스폿은 리튬에 의한 세포 밖에서 존재할 수있다posomes 유리에 쌓여있다. 이들은 멤브레인 부에 형광 주로 균일 ROI를 선택하여 폐기 될 수있다 (예., 세포 원형질막). 예상 한 바와 같이이 지질에 대한 측정 된 확산 법칙 (도 4C)은 이전 STED-FCS 측정 30,35 같이 주로 자유 확산을 나타내는 편평하다. 이것은 분명히이 접근의 능력을 나타내는, 모든 도시 된 변위 값이 회절 한계 이하 것을 언급 할 가치가 수십 나노 미터에 잘 회절 한계 이하 및 아래쪽 평균 분자량 변위 슈퍼 - 해결.
막에 TFR-GFP 이량 체 삽입의 도식화는도 5a에 표시됩니다. 많은 연구는이 수용체의 세포질 꼬리 차례로 수용체 이동성 12,40위한 울타리 역할을하는 막 골격과 상호 작용하는 것으로 나타났다. TFR-GFP를 발현하는 CHO 세포의 대표적인 TIRF 이미지이다 prese그림 (b)에 nted. 멤브레인 네이티브 조건 최소화 과발현과 관련된 아티팩트 확률에 가까운 낮은 형광 강도 세포, 바람직 할 것이다. 또한 셀의 중앙 부분이 존재할 수있다 (세포질에서, 예를 들어) 포커스 아웃 형광의 영향과, 피해야한다. 예상대로 측정 확산 법 TFR-GFP에 대한 (그림 5C)는 0.2 μm의 2에 이르기까지 명백한 확산에 필연적 급감 한 후, 약 0.7 μm의 2 초 -1의 평균 D의 응용 프로그램과 함께, 100 나노 미터 이하의 첫번째 플랫 동작을 보여줍니다 초 -1 (값은 일반적으로 측정 회절 한정 FCS 12). 이 결과는 우리의 접근이 용이하게 나노 미터 수십 해상도 GFP 표지 단백질의 평균 변위를 측정 할 수 있음을 보여준다. 또한 D 응용 프로그램이 시작되는 공간 규모 세트하는 특성을 감소이전 추정치 6으로 유지하는 약 120 nm에서 막 골격에 의해 단백질 부분 감금의 공간 규모.

포인트 확산 기능의 그림 1 보정. (A) 절연 비드와 비드 응집체의 의사 색조 이미지. 절연 비드의 강도 프로파일의 (B)의 3D 플롯은 잘 정의 된 가우시안 프로파일을 나타낸다. (C) 가우스 함수에 의한 세기 분포의 맞춤 (상판) 해당 잔차 (하판)와 함께. 장착 분포와 측정 된 강도 프로파일 사이 좋은 계약은 또한 악기 PSF는 가우시안 함수로 근사 할 수있는 증거입니다. vi를하려면 여기를 클릭하십시오 이 그림의 더 큰 버전을 으으.

단일 광자에 카메라 응답의 그림 2 교정. 그림은 자른 센서 모드에서 32 X 128 ROI 카메라 배경, 노출 0.5 밀리 초에 대한 디지털 레벨 (DL) 분포를 보여줍니다. 약 180 DL에서의 피크는 더 광자 카메라의 응답을 나타냅니다. 특히, 아날로그 디지털 (AD) 변환기의 기여를 나타내며, 오프셋 및 신호 기록에 의해 도입 된 분산을 추정하기 위해 가우시안 함수에 의해 근사 될 수있다. DL (200)보다도 디지털 레벨의 분포는 지수되고 단일 광자에 대한 평균 응답 카메라를 나타낸다. 이러한 파라미터의 측정은 수집 중에 기록되는 광자의 밀도를 추정 할 수있다.S / ftp_upload / 51,994 / 51994fig2highres.jpg "대상 ="_ 빈 ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

방법의 그림 3 도식화. (A) EMCCD 카메라에 의해 넓은 시야 이미징 TIRF의 축소 복사가 플라즈마 멤브레인의 정확한 광학 절편을 제공하기 위해 이용되는 동안, 서브 밀리 초 해상도에 도달하기 위해 적용된다. (B) 화상의 생성 스택 평균 공간을 계산하기 위하여 자기 상관된다 -temporal 상관 함수. 이 상관 함수는 잘 가우스 함수에 의해 근사화된다 (개요를 참조) 그리고 입자 변위에 따라 시간에 퍼진다. (C)을 따라서 인해 분자 변위 상관 함수의 확산을 정량화하기 위해, 가우스 피팅 이안의 기능이 수행됩니다. 이것은 평균 변위 플롯 대 겉보기 확산율의 형태로 직접 영상에서 분자 '확산 법'을 측정한다. (D) 덕분에 이러한 플롯에 분자 확산 모드가 직접 대한 해석 모델 또는 가정을 할 필요없이 식별 될 수있다 멤브레인의 공간적 조직. 그들의 이동도 측정의 공간적 규모에 의존하지 않는 점에서, 분자는 자유롭게 확산 상수 명백한 확산 계수를 표시한다. 대조적으로, 부분적으로 제한된 분자 한정 크기보다 작은 변위 아주 명백한 확산 상수를 표시하고 협착 크기보다 큰 공간 규모위한 감소 확산율. 관련된 공간적 스케일이 협착의 공간 확장을 추정하는데 이용 될 수있다 따라서, 명백한 확산의 감소의 외관은, 과도 협착의 지문과 같이 해석 될 수있다. .jove.com / 파일 / ftp_upload / 51,994 / 51994fig3highres.jpg "대상 ="_ 빈 ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

라이브 세포막의 그림 4 ATTO488-PPE 확산 법칙 (A) 세포막에서 ATTO488-PPE 삽입의 도식 표현 (B) ATTO488-PPE으로 표시 CHO 기저막의 TIRF 이미지 :.. ROI (빨간색 상자)를 선택 선택된 ROI 측정 확산 법이 구성 요소에 대한 무료 확산 모델을 확인하는 평면 동작을 보여줍니다 셀 테두리 높은 형광 반점 피 세포의 대부분 균일 한 부분에. (C). 의 더 큰 버전을 보려면 여기를 클릭하십시오 이 그림.

생균 세포막도 5 TFR-GFP 확산 법칙 (A) 세포막에서 TFR-GFP 삽입의 도식 표현 :. 수용체의 세포질 꼬리 수용체 이동성 울타리 역할 막 골격과 상호 작용 (B를. ) CHO 표현 TFR-GFP의 TIRF 이미지 : ROI 인해 과발현에 유물을 피하기 위해 낮은 발현하는 세포를 선호하는 선택 (C)를 TFR (검은 점의 확산 법칙), PPE는 달리 (회색 선) 그림 4에서 촬영. 첫번째 평평한 부분이 D 응용 프로그램의 감소 뒤에 부분적으로 제한 확산의 전형적인 동작은. 보여줍니다 이 파이의 더 큰 버전을 보려면 여기를 클릭하십시오gure.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
단일 입자 추적 (SPT)은 분자 역학을 연구하는 가장 일반적인 전략 중 하나를 의미하며, 입자의 궤적을 측정의 큰 장점을 갖는다. 차례로 이것은 복잡한 시스템에도 몇몇 표지 입자의 거동을 프로빙한다. 그러나, 일반적 SPT 프로브와 매우 밝은 라벨 저밀도 필요 이러한 이점을 달성한다. 생산 시스템으로, 라벨 및 삽입의 복잡한 절차가 필요하다이 경우 : 특히, 높은 시간 해상도 (마이크로 초 범위) 무기 프로브는 일반적으로 요구된다 (예를 들면, 양자 도트 나 금속 나노 입자)를 얻을 수있다. SPT에 비해 본 발명의 방법은 몇 가지 중요한 장점을 보여줍니다. 첫째,이 방법은 형광 단백질과 접합하여 사용할 수있다. 따라서 SPT에 비해,보다 높은 시간 해상도가 광자의 하부 양 덕분 (동일한 라벨) 달성 34 필수. 보다 상세하게는,이 속성은 시간 레졸을 밀어 수 있습니다10 아래의 ution - 3 초는 encodable 형광 단백질을 사용하는 경우,이 척도는 막 성분의 나노 역학에 단독으로 액세스 할 수 있습니다. 마지막으로, 분자 확산 법률이 각 분자를 추적 할 필요없이, 전체 공간 - 시간 상관 함수를 분석하여 설명되어 있다는 점에 주목할 필요가있다.
STED 기반 FCS와 비교도 흥미 롭다. STED-FCS 측정에서 관측 볼륨을 감소시키는 분자의 평균 통과 시간은 형광 신호의 시간 상관에 의해 측정된다. 이것은 또한, 회절 한계 이하의 분자 역학 로컬 측정을 획득한다. 제시된 방법에서는, 확산 법칙은 표준, 회절 제한, 볼륨의 관찰 수단에 의해 관찰 선택된 ROI에서 이동하는 모든 입자의 평균으로서 측정된다. 그러나,이 방법의 결과는 회절에 의해 제한되지 않는다는 것을 입증 있지만 번째 의해보고전자 시간 해상도 사용할 수 있습니다. 회절 제한 취득 잘 회절 한계 이하 (유사 같은 PALM 및 STORM 다른 초 해상 기술에서 수행되는 것과) 변동, 분자 변위를 검출하기 위해 사용되지만 실제로, 이미, (직접) 될 산출 할 분자량 측정하기 STICS을 사용하여 입증 32을 흐른다. 더욱이 STED-FCS 달리이 방법은 쉽게 래스터 주사 현미경 또는 넓은 시야 카메라 기반의 현미경과 같은 상용 기존 현미경 셋업의 넓은 범위에 적용될 수있다. 그것은 분자 확산 법률의 STED-FCS 측정이 엄격하게 쓸모있는 허리의 크기의 형광 의존 교정을 필요로 언급 할 가치가있다. 반대로, 측정 (입자 크기의 추정을 위해 필요한 경우에만) 시스템 교정을 필요로하지 않고 여기에 제시.
제시된 방법 드에 의한 입자의 변위 측정의 실제 해상도우리는 상관 함수를 정확하게 측정 할 수있는 방법에 좌우된다. 결과적으로, 그것은 본질적으로 유사 해상도 입자 "화상"을 정확하게 측정하는 방법에 의존 SPT 경우에, 회절에 의해 제한되지 않는다. 제시된 실험 1 분 미만에서 유의 한 상관 관계를 측정하기 위해, 각 프레임의 각 입자에 대한 몇몇의 광자 (포톤 보통 10 이하)가 충분하다. 상관 함수가 계산 될 때 사실상, 모든 관측 된 입자의 기여는 입자가 절연되어 있지 않은 경우에도, 함께 평균화된다. 이 속성은 변동의 상관 방법의 고유하고 라이브 세포에서 형질 전환 형광 단백질과 같은 희미하고 조밀 한 라벨을 사용하실 수 있습니다.
이를 염두에두고 그 변위 측정 가능한 최소 입자의 확산에 촬상 설정의 시간 해상도에 의존한다는 것을 분명 나타난다. 예로서, 세포막의 분자의 확산을 고려하세요단백질이나 지질의 최대 측정 확산은 약 5 μm의 2 초 -1입니다. 이러한 조건에서, 우리는 50 나노 미터의 평균 변위를 잡으려고 약 10-4 초의 시간 해상도가 필요하다. 여기 켰을 때,이 시간 해상도는, 하나의 라인을 따라 또는 시간 해상도는 노광 시간에 일치하고 빠른 EMCCD 카메라에 의해 빠르게 주사 현미경에 의해 달성 될 수있다.
정확하게 분자 역학을 설명하기 위해이 방법에 대한 추가 필수 요구 사항은 정확한 공간 샘플링이다. 사실, 상관 함수에 맞추기 위해 우리는 쓸모 PSF의 허리보다 낮은 공간적 샘플링 (픽셀 크기)가 필요. 대부분의 상용 현미경 (촛점 또는 와이드 필드)에서는 PSF 허리가 200 nm 내지 500 nm의 스팬 (사용 선택된 대물의 개구 수와 파장에 주로 의존) 쉽게 교정 실험 이용한 나노으로 측정 할 수있다 크기의 형광 구슬. THUS (기악 허리보다 3 배 더 낮은) 70-150 nm의 화소 사이즈가 충분할 수있다. 그러나, 화소의 크기는 고려 간단한 규칙을 가지고 연구중인 시스템에 적용될 수있다 : 상관 함수의 설명에서 픽셀 크기, 더 높은 정확도를 저하. 또한, 취득되는 영상의 최소 크기는 관심의 최대 변위 (플러스 쓸모 허리)보다 적어도 3 배 이상이어야한다. 이것은 피팅 알고리즘의 수렴 좋은 분자 변위의 통계적으로 유의 한 샘플에 도달하기 위해 요구된다. 예를 들어, 수백 나노 미터 (예, 200 nm의) 몇 마이크론의 이미지 크기보다 충분히 작은 평균 분자량 변위 공부. 또한, 상관 함수의 품질에 미치는 영향 (정수 픽셀 크기 복용)을 픽셀들의 전체 숫자. 실제로, 큰 이미지하더라도 시간 resolut의 비용, 상관 함수에 대한 추가 정보를 평균 허용이온. 여기에 사용 된 카메라 기반 시스템에 관한, 칩의 픽셀의 실제 크기가 고정되어 있으므로주의하시기 바랍니다. 따라서, 픽셀 크기를 감소시키는 것은, (픽셀 크기의 제곱에 따라 다름), 각 화소에 신호를 낮춘다 시야를 감소하고, 고배율 전력을 필요로한다. 한편, 관찰 영역은 픽셀 크기는 일반적으로 시간 해상도를 희생 증가 화소 수가 감소하는 결과, 고정 된 스캐닝 시스템에.
사용되는 검출기에 대한 몇 가지 세부 사항이 논의되어야한다. 단일 광자 검출기는 달리 EMCCD 시스템 인해 오프셋의 존재로 수집 된 빛에 직접 비례하지 않는다 평균 강도 (디지털 레벨, DL)을 측정한다. 오프셋이 카메라의 동적 범위에 비해 낮다하더라도 무시할 많은 광자를 수집 실험 (몇 백은이 16 비트 읽기 (16)에 비해), 그것으로 고려되어야상관 함수의 정확한 정규화를 획득. 또한, 수집 된 신호의 양을 식별하기 위해 낮은 - 빛 조건에서 참조로서 사용될 수있다 오프셋. 또한, 취득 중에 수집되는 광자의 평균 양을 추정하기 위해, 평균 레벨 디지털 각 광자 수집에 관련된 측정되어야한다. 이 양이 매우 낮은 빛의 세기에 카메라를 노출하여 검색 할 수 있습니다 (예를 들어, 방에 배경 조명); 실제로,이 경우에, 우리는 합리적으로 측정 된 강도가 제로 또는 하나의 광자 만 관련 될 수있다, 즉 그 단지 단일 광자, 카메라 도달 가정 할 수있다.
마지막으로, 우리는 어떤 대안 수집 시스템 (즉, 다른 현미경 작아도)되게 측정을 수행하는 데 어떻게 사용될 수 있는지에 대해 언급하자. 먼저, 수학 식 2에서 'W'factor (즉 악기 PSF의 자기 상관을 나타낸다) t하도록 구성 될 수있다실험적인 상관 함수를 맞추기 위해 사용되는 그 특정 수집 시스템. 이전 34에 도시 된 바와 같이, 케이스는 쉽게 주사 속도는 입자 역학보다 상당히 높은 경우 취득 레이저 스캐닝 현미경 엉이다. 이러한 경우에, 실제로 획득 시간 동안 입자의 이동 (즉, 라인 시간)은 무시할 수 있고 상관 함수 잘 가우스 함수에 의해 근사된다. 새로운 이미징 기술의 맥락에서 흥미로운 접근 방법은 샘플 (41)을 통해 매우 얇은 광 시트 (1-2 μM)를 생성 할 수있는 가능성에 기초한다. 빛 시트는 3D (42)의 카메라 기반 수집 시스템, 빠른 광 절편과 함께 하나의 샘플에서면 (1면 조명 현미경, SPIM) 및, 선택적으로 조명 할 수 있습니다. 때문에 이러한 특성, SPIM이 성공적으로 FCS (43)가 결합되었으며 vali을 나타낼 수개발 툴은 살아있는 세포의 세포질이나 핵 같은 3D 환경에 제시된 분석을 확장한다.
뷰의 실험 점에서, 요약하면 본 방법은 빠른 획득 모듈이 장착 현미경에만 액세스를 필요로한다. 관심의 단백질은 따라서 여러 가지 빛깔의 이미징 또한 수있는 형광 단백질 또는 유기 형광 태그 할 수 있습니다. 이러한 상황에서, 우리는 분자의 하위 집단을 선택하고 라이브 세포막에 작용 공동 확산을 드러내 I MSD 분석을 사용하여 교차 가능성을 구상. 마지막으로, 우리는이 방법은 세포막에 nanodomains 내에서 동적 분할을 겪고 단백질 및 / 또는 지질을 연구하는 강력한 도구를 나타낼 수 있다고 생각합니다. 이 경우, nanodomains의 고 가변 크기 및 수명이 2 색을 포함하여 상기 촬상 방법 론적 구현을 필요로 실제 데이터의 복잡도의 수준을 추가로 도입 로컬분석 (예를 들어, 2D 쌍 상관) 및 / 또는 형광 이방성.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}